

Протисты / Карпов С.А. Строение клеток протистов
.pdfГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
гих водорослей насчитывается более 2 мембран. Такие пластиды называются сложными и они, как было показано в предыдущей главе, характерны для динофитовых и эвгленовых, имеющих 3 мембраны в оболочке пластиды, а также для всех охрофитов, хлорарахниофитов и криптофитовых, имеющих 4 мембраны. Наружная мембрана таких пластид обычно считается принадлежащей самой клетке, и интерпретируется как мембрана бывшей симбионтофорной вакуоли.
Редкие авторы относят к сложным пластидам также хлоропласты красных и зеленых водорослей, считая их результатом вторичного эндосимбиоза, в процессе эволюции утративших одну или 2 мембраны.
Наиболее веским доказательством приобретения сложных пластид путем эндосимбиоза с эукариотными водорослями считается наличие нуклеоморфа в перипластидном пространстве криптофитовых, хлорарахниофитовых и некоторых динофитовых.
Ультраструктурные исследования нуклеоморфа стимулировали его углубленное изучение молекулярными методами. Было показано, что нуклеоморф криптомонад и Chlorarachnion содержит ДНК, организованную в 3 хромосомы. Хромосомы очень короткие (всего 380 000 оснований) и содержат несколько генов, кодирующих рРНК малой субъединицы, сплайсинговые белки, которые обеспечивают процесс считывания ДНК, некоторые рибосомальные белки и протеазы. Наличие этих генов и процесса считывания информации позволило предположить, что нуклеоморф является сильно редуцированным ядром эукариотной клетки. Другие гены, необходимые для функционирования пластид, были перенесены в ядро клетки хозяина. Таким образом, синтезируемые в цитоплазме клетки белки пластид должны преодолеть 4 мембраны, чтобы попасть в хлоропласт на место своей окончательной локализации.
Сравнение сиквенсов генов нуклеоморфа, ядра клетки и пластид, а также состав пигментов и морфологические данные показывают, что сложные пластиды хлорарахниофитов и криптомонад приобретены независимо. Пластиды хлорарахниофитов получены в результате симбиоза амебы с зелеными во-
250

4.5. ЯДРО И МИТОТИЧЕСКИЙ АППАРАТ
дорослями (рис. 4.75), а хлоропласты криптофитов – в результате симбиоза похожего на Goniomonas криптофита и красной водоросли.
Эти примеры показывают, что и другие пластиды с 3 и 4 мембранами могли появиться в результате симбиоза гетеротрофных протистов и фотосинтезирующих водорослей (рис. 4.75). При этом предполагается, что у водорослей без нуклеоморфа в хлоропласте редукционные процессы зашли дальше, чем у криптомонад и хлорарахниофитов и привели к полной утрате ядра и редукции перипластидного пространства.
Полный цикл приобретения и дальнейшей утраты пластид, по-видимому, имел место у криптофитовых. Среди них есть первично бесцветные формы (Goniomonas), фототрофные организмы с хлоропластами (Cryptomonas) и вторично бесцветные формы (Chilomonas), которые содержат в перинуклеарном пространстве лейкопласт, не способный к фотосинтезу.
4.5. Ядро и митотический аппарат
Клеточные ядра протистов имеют типичную для ядер эукариот структуру. Вместе с тем, они значительно различаются по форме и количеству в клетке. Способы деления ядер протистов гораздо многообразнее, чем у многоклеточных животных и растений. В пределах протистов происходило, вероятно, становление мейоза и полового процесса, что нашло отражение в существенных различиях этих процессов у разных групп.
4.5.1. Число и размеры ядер
Большинство протистов обладают одним клеточным ядром. У большинства микроспоридий, а также у многих грибов и некоторых мастигамеб клетка содержит два ядра (диплокарион, или дикарион). Некоторые солнечники, крупные свободноживущие амебы, миксомицеты – многоядерны. Размеры и форма ядер варьируют в самых широких пределах, так же, как их морфология и соотношение структурных компонентов ядра. Наиболее мелкие ядра (у пироплазмид, лейшманий, микронукле-
251

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
усы некоторых инфузорий) имеют диаметр 1–3 мкм. Крупные сферические ядра некоторых полицистин достигают в диаметре 400 мкм и даже 1 мм. Форма ядер большинства простейших приближается к сферической, но иногда может принимать самые причудливые формы. Это относится преимущественно к богатым хроматином вегетативным ядрам инфузорий.
4.5.2.Структурные компоненты ядер
Óпротистов они те же, что и в других эукариотических клетках (рис. 4.76). Ядерная оболочка состоит из двух мембран и имеет поры, число которых у разных видов варьирует в широких пределах, но особенно велико у тех ядер, которые обладают высокой функциональной активностью. Снаружи ядерная оболочка обыч- но покрыта рибосомами, но к ней могут примыкать дополнительные слои, располагающиеся как снаружи, так и под оболоч- кой со стороны кариолимфы. В частности, у многих свободноживущих амеб к внутренней стороне ядерной оболоч- ки прилегает так называемый фиброзный, или сотовый, слой, имеющий ячеистое строение и значительно превосходящий ядерную оболочку по толщине. У Sticholonche этот слой достигает зна- чительной толщины, образуя так называемую нуклеотеку. Фибриллярный материал нуклеотеки откладывается, по-видимому,
ñвнутренней стороны ядра. Определенные сайты на наружной мембране оболочки могут служить ЦОМТами аксонем у актинофриидных солнечников, пединеллид и некоторых полицистин. Наружная мембрана может усиливаться фибриллярным материалом у некоторых рафидофитовых, гипермастигин, фораминифер и голых амеб. Помимо фибриллярного материала к поверхности ядра может прилегать гранулярный слой, который дополняется еще слоем мелких пузырьков, как у раковинной амебы Hyalosphenia. Многие фораминиферы имеют ядра с прилегающим снаружи слоем мелко вакуолизированной цитоплазмы, образующей так называемый экзонуклеарный вакуом.
Некоторое представление о многообразии ядер дает рисунок 4.77, на котором изображены только неполиплоидные интерфазные ядра различных протистов, как они видны в световой
252
4.5. ЯДРО И МИТОТИЧЕСКИЙ АППАРАТ
микроскоп. В перинуклеарном пространстве у многих водорослей располагаются хлоропласты или лейкопласты, а у некоторых видов протистов обнаружены также симбиотические бактерии.
Обязательным компонентом всех ядер протистов является хроматин, представляющий ДНК в комплексе с основными белками. Ультраструктурный анализ показывает, что он состоит из линейно расположенных глобулярных телец - нуклеосом (включающих как ДНК, так и гистоны). Упаковка нити ДНК в ядре показана на рисунке 4.78. Кроме хроматина хромосомной природы, в ядре может находиться внехромосомный хроматин, возникающий в результате повторного удвоения (амплификации) некоторых генов. Такой внехромосомный хроматин может входить в состав ядрышек вместе с РНК.
Распределение хроматина в интерфазных ядрах у простейших носит различный характер. Подчас происходит сильная деспирализация и деконденсация хроматина. В других случа- ях деконденсация охватывает часть хроматина. В некоторых интерфазных ядрах хроматин, наоборот, очень сильно конденсирован и представляет собой сплошную массу, как в головках спермиев многоклеточных животных.
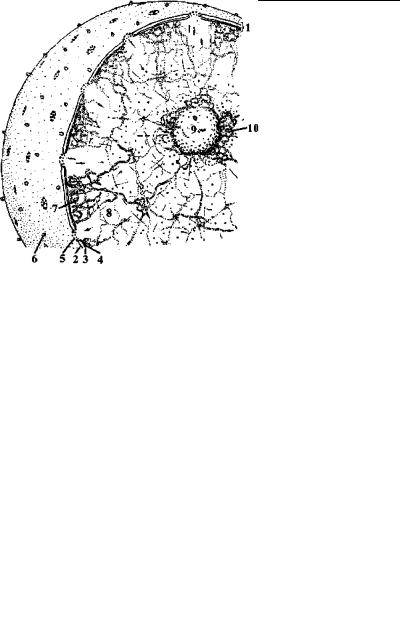
Рис. 4.76. Схема строения интерфазного ядра эукариотной клетки. (По: Заварзин и др., 1992.)
1 – поверхностный аппарат ядра, 2 – наружная и 3 – внутренняя мембраны, разделенные перинуклеарным пространством, 4 – плотная пластинка, 5 – поровый комплекс, 6 – рибосомы, 7 – гетерохроматин, 8 – эухроматин, 9 – ядрышко, 10 – околоядрышковый гетерохроматин.
253
ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
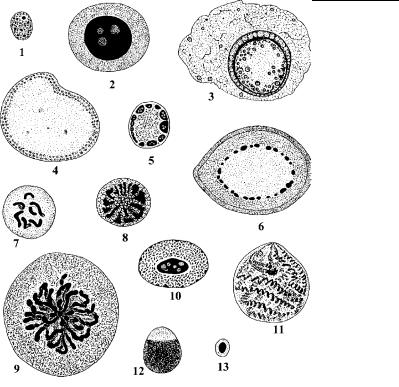
Рис. 4.77. Многообразие интерфазных неполиплоидных ядер протистов. (По: Raikov, 1982.)
1 – Trichomonas muris, 2500x; 2 – Amoeba sphaeronucleolus, 1500x; 3 – макрогамета Aggregata eberthi, 1000x; 4 – Amoeba crystalligera, 1500x; 5 – Amoeba terricola, 1500x; 6 – Endamoeba blattae, 1500õ; 7 – Euglypha sp., 1000x; 8 – Actinosphaerium eichorni, 1450x; 9 – Duboscquella aspida, 1200x; 10 – Euglena sp., 1200x; 11 – Holomastigotoides psammotermitidis, 1500x; 12 – микронуклеус Paramecium caudatum, 1500x; 13 – микронуклеус
P. aurelia, 1500x.
Ядрышки, которых может быть в ядре одно или несколько, а иногда и очень много, в основном состоят из рибонуклеопротеинов – предшественников рибосом. Электронная микроскопия показывает, что ядрышки образованы из двух структурных компонентов – гранулярного и фибриллярного. Фибриллы диаметром 5–10 нм обычно локализованы в центральных частях
254
4.5. ЯДРО И МИТОТИЧЕСКИЙ АППАРАТ
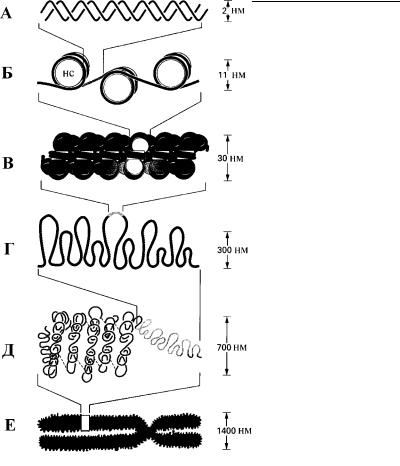
Рис. 4.78. Модель упаковки хроматина в ядре эукариотной клетки. (По разным авторам.)
А – двойная спираль ДНК, Б – нуклеосомная нить (ДНК «намотана» на нуклеосомы (нс), состоящие из гистонов), В – хроматиновая фибрилла – следующий уровень упаковки нуклеосомной нити, Г – участок деспирализованного хроматина (эухроматин), Д – участок спирализованного хроматина (гетерохроматин), Е – метафазная хромосома.
ядрышка. Во многих ядрышках у протистов описаны включе- ния хроматина, которые иногда, но не всегда, морфологически
255

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
связаны с определенными хромосомами. Это – ядрышковые организаторы, образующиеся в результате амплификации рибосомальных генов.
В ядрах простейших в большем или меньшем количестве присутствует ядерный сок (кариолимфа). В нем могут быть также мельчайшие гранулы разной природы, часть которых является, вероятно, субъединицами рибосом. Встречаются также паракристаллические или фибриллярные белковые образования, а также особые спиральные структуры (у крупных амеб), содержащие, по-видимому, РНК.
Наиболее существенные отклонения от описанной структуры ядра отмечены только для динофитовых. Хроматин ядра динофитовых (динокариона) лишен гистонов или сильно ими обеднен, поэтому у них отсутствует нуклеосомная организация хроматина. ДНК динофитовых укладывается особым образом. Кольцевые хроматиновые нити, из которых состоят хромосомы, уложены в виде восьмерок (рис. 4.79), и на ультратонких срезах выглядят совершенно иначе, чем хроматин остальных эукариот. Сама хроматиновая нить состоит из спирально упакованной молекулы ДНК, не связанной с белками.
Этот тип организации интерфазного ядра долгое время счи- тали мезокарионом – промежуточным между прокариотическим и собственно эукариотическим, а отсутствие гистонов счи- талось первичным признаком. В настоящее время общепринята точка зрения, что у динофитовых произошла утрата гистонов в процессе эволюции.
Рис. 4.79. Схема строения хромосомы динокариона. (По: Райков, 1978.)
хн – хроматиновая нить, хс – контуры хромосомы.
256

4.5. ЯДРО И МИТОТИЧЕСКИЙ АППАРАТ
4.5.3. Гетероморфизм ядер
Подавляющее большинство многоядерных протистов содержит в клетке одинаковые (гомоморфные) ядра. Лишь инфузории, миксоспоридии, некоторые фораминиферы, а также акантарии обладают двумя сортами ядер к клетке и обычно называются гетероморфными. Ядра различаются морфологически и функционально. У инфузорий эта их особенность называется ядерным дуализмом. Обычно клетка содержит макронуклеус (Ма) и микронуклеус (Ми). Макронуклеус – крупное ядро нередко весьма причудливой формы – является соматическим ядром и определяет фенотип особи. В нем часто наблюдается амплификация генов и ярко выражена экспрессия генома. Микронуклеус – небольшое ядро с компактным хроматином – является генеративным ядром, хранящим наследственную информацию в диплоидном состоянии и почти лишенным экспрессии генов. Ма некоторых высокоорганизованных инфузорий (гипотрихи) может содержать всего 5% генома Ми. Он состоит только из функционирующих генов, но тиражированных миллионами копий. Это связано с большими размерами клетки инфузорий, т.к. количество копий в Ма, как и его размеры, обычно пропорционально размерам особи.
Дифференцировка ядер на генеративные и соматические характерна для всех изученных миксоспоридий. Часто она происходит после второго или третьего деления ядра амебоидного зародыша и приводит в конечном итоге к образованию дифференцированных клеток многоклеточного организма.
Гетерокариоз фораминифер отмечен только на некоторых стадиях их жизненного цикла (агамонты). Он менее изучен, чем у инфузорий, и характерен только для эволюционно продвинутых видов.
У акантарий дифференцировка ядер на соматические и генеративные происходит перед инцистированием. И те и другие формируются из одного полиплоидного ядра, от которого сна- чала отпочковываются соматические ядра, а оставшееся вторич- ное ядро дает начало генеративным ядрам, количество которых в результате многократных делений достигает сотен на клетку.
257

ГЛАВА 4. СТРОЕНИЕ КЛЕТКИ ПРОТИСТОВ
Гетерокариотическое состояние следует отличать от наличия в клетках некоторых протистов разнокачественных геномов, принадлежащих разным видам. Так, некоторые динофитовые содержат в цитоплазме ядра симбиотических эукариот, которые отличаются по составу генома и организации хроматина. Проблема взаимодействия различных геномов в клетке протистов очень сложна и необычайно интересна. В некоторых группах протистов, например у криптофитовых, насчитывается до 4 различных геномов в одной клетке (ядерный, пластидный, митохондриальный и геном нуклеоморфа). Их даже образно называют клетками-химерами.
4.5.4. Митоз
Основной формой деления ядра простейших, как и всех эукариотических организмов, является митоз. Главная черта митоза – закономерное расхождение в дочерние ядра двух копий каждой реплицированной хромосомы (хроматиды) – хорошо известна. Этот важнейший биологический процесс обеспечивает преемственность и непрерывность наследственной информации. Хотя общая схема и сущность митоза едины для всех эукариот, в деталях этот процесс может протекать по-разному. Поведение и судьба участвующих в митозе компонентов – ядерной оболочки, центриолей, ахроматинового аппарата клетки – варьируют у протистов в широких пределах. Здесь как бы осуществлялись разные попытки реализации митоза, из которых лишь немногие были заимствованы наземными растениями, настоящими грибами и животными.
Классификация типов митоза у протистов предпринималась неоднократно. По мере накопления знаний она становилась все более дифференцированной. Особенное значение для углубления представлений о многообразии форм митоза имела электронная микроскопия, которая позволила раскрыть ряд ранее неизвестных фактов.
В настоящей книге мы принимаем классификацию И.Б.Райкова (Raikov, 1982, 1994), которая, в свою очередь, представляет собою дальнейшее усовершенствование классификации,
258

4.5. ЯДРО И МИТОТИЧЕСКИЙ АППАРАТ
предложенной ранее А.Холландом (Hollande, 1972). И.Б.Райков выделяет следующие 6 форм митоза, хорошо отличающихся друг от друга на стадии метафазы (рис. 4.80):
Открытый митоз
1.Открытый ортомитоз (эумитоз)
Полузакрытый митоз
2.Полузакрытый ортомитоз
3.Полузакрытый плевромитоз (парамитоз)
Закрытый митоз с внутренним веретеном
4.Внутриядерный плевромитоз
5.Внутриядерный ортомитоз
Закрытый митоз с внеядерным веретеном
6. Внеядерный плевромитоз При открытом митозе, в отличие от закрытого, разрушается
ядерная оболочка, а хромосомы оказываются свободно лежащими в цитоплазме. Закрытым митозом именуют такие его формы, при которых ядерная оболочка сохраняется в течение всего процесса. При ортомитозе в метафазе хромосомы становятся по экватору веретена, образуя метафазную экваториальную пластинку. При плевромитозе экваториальная пластинка, как правило, не образуется и ахроматиновое веретено представлено двумя полуверетенами, расположенными под углом друг к другу вне или внутри ядра, в результате чего вся фигура деления приобретает асимметричный (радиально несимметричный) характер. Различные сочетания указанных черт и образуют 6 категорий митоза (рис. 4.80), которые определяются по митотическим фигурам.
Строение центров организации микротрубочек (ЦОМТ) митотического веретена также весьма разнообразно. При открытых и полузакрытых митозах ими чаще всего являются центриоли, каждая из которых представлена парой кинетосом. Иногда кинетосомы присутствуют в клетке наряду с центриолями (у некоторых зеленых водорослей). ЦОМТы могут быть представлены фибриллярными корешками (ризопласты у хлорофитовых и хризофитовых, или атрактофоры у гипермастигин) или образованным тубулином аморфным материалом, из которого формируются микротрубочки веретена деления ядра.
259