

Бабьева И. П., Чернов И. Ю. Биология дрожжей. М. Товарищество научных изданий КМК, 2004
.pdfторые на срезах представляют собой либо гомогенные структуры, либо комплексы концентрически расположенных мембран. Липиды в дрожжевых клетках хорошо различимы в световом микроскопе и видны, как резко преломляющие свет, блестящие круглые тельца - «жировые капли». Такихкапельвклеткахможетбытьнесколько, илижеонисливаются в одну крупную глобулу, которая у некоторых видов, например
Cryptococcus terricola, Lipomyces starkeyi, можетзаниматьбольшуючасть клетки (рис. 8).
Изменения клеточных структур в онтогенезе
Цитологические особенности дрожжевой клетки могут существенно изменяться в зависимости от ее возраста и стадии онтогенеза. Новая молодая дрожжевая клетка развивается либо из споры при ее прорастании, либо образуется вегетативно при почковании материнской клетки. В споре сохраняются основные клеточные структуры: ядро, митохондрии, цитоплазматическая мембрана и элементы ЭПР. Уже через 1-2 часа после набухания и прорастания споры в клетке видны сильно удлиненные митохондрии. Это начало их подготовки к делению. Через 3-4 часа на вогнутой поверхности ядра происходит активация наружного листка ядерной мембраны ивэтом месте формируются новыемембраны, часть изкоторыхвходитвкомплексГольджи, адругие, возможно, включаются
в систему ЭПР. Затем начинается деление митохондрий и развитие вакуолярного аппарата на основе мембран ЭПР.
При почковании происходит активация цитоплазматической мембраны и транспортных везикул, переход в почку митохондрий и части мембран ЭПР. Эндомитоз ядра начинается в материнской
Рис. 8. Липиды в клетках Lipomyces starkeyi. клетке уже после образо-
21
вания почки. Ядро вытягивается без нарушения целостности ядерной мембраны, передвигаетсякпочкеичерезузкийперешеекперемещается частично в почку. В ядре молодой клетки поры расширены, что свидетельствует о напряженности обменных процессов. Митохондрии в новой клетке образуются путем деления, а другие мембраны развиваются наоснове«затравочных» компонентовпутемудлиненияилиматричного механизма. В более зрелых клетках мембраны эндоплазматической сети образуют отдельные цистерны, которые превращаются в разного рода вакуоли и везикулы.
Клеточнаястенкапомерестаренияклеткистановитсятолще, уаскомицетовых дрожжей в ней насчитывается более 4 слоев. Увеличивается содержание хитина, который локализуется в области шрамов почкования. Многочисленныешрамыпочкованиясильноменяютструктуруклеточной поверхности.
Капсула у молодых клеток иногда сразу хорошо выражена, если формированиепочекпроисходитвсреде, богатойуглеводами. Онапредставляет собой как бы продолжение того слизистого покрова, который окружает материнскую клетку, но обычно на молодой клетке этот слой тоньше. Затем толщина капсулы растет, но часть поверхностного материала постоянно переходит в среду, образуя фракцию так называемых внеклеточных полисахаридов.
Доля запасных веществ - липидов, гликогена - возрастает по мере ростаклетки, апристарениивновьпадаетзасчетихчастичногопотребления клеткой на энергетические нужды.
Цитологические особенности дрожжей в различных условиях роста
Значительные изменения в структурной организации клетки происходят при смене условий роста: при переходе от анаэробного к аэробному культивированию, замене состава среды и концентрации лимитирующих субстратов, при недостатке факторов роста, а также при стрессовыхфизиологическихвоздействиях. Особеннозаметныультраструктурные перестройки мембранного аппарата при аэробно-анаэробных сдвигах. В условиях анаэробного брожения митохондрии неузнаваемо меняют свой облик: они объединяются, кристы распрямляются. И наоборот,
22
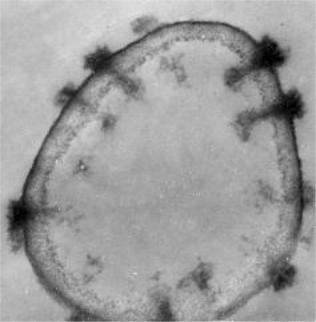
при аэрации митохондрии активно делятся, число их возрастает, в них хорошозаметныкристы. Ванаэробныхусловияхвклеткахнакапливается больше гликогена и он представлен крупными гранулами, а иногда образует конгломераты.
При росте дрожжей на углеводородах в их клеточных стенках появляются особые зоны (каналы), через которые нерастворимые в воде углеводородыпроникаютвклетку (рис. 9). Участкиклеточнойповерхности разрыхляются и приобретают форму купола. Высшие жирные кисло- тыин-алканы, проходячерезклеточнуюстенку, накапливаютсявпериплазматическом пространстве и далее поступают в клетку путем пиноци-
тоза и «просачивания» через цитоплазматическую мемб-
|
рану в каналы ЭПР. |
|
Метилотрофныедрожжи, |
|
растущие на метаноле, обра- |
|
зуюткрупныепероксисомы - |
|
вакуоли, заполненные фер- |
|
ментными белками с упо- |
|
рядоченнойкристаллической |
|
структурой (рис. 10). Они |
|
резко преломляют свет и хо- |
|
рошовидныприфазово-кон- |
|
трастном микроскопирова- |
|
нии в виде блестящих крис- |
Рис. 9. Ультратонкий срез дрожжевой |
таллов внутри клетки. |
клетки, растущей на н-алканах, видны ка- |
У капсулообразующих |
налывклеточнойстенке, черезкоторыене- |
дрожжейприобильномснаб- |
растворимыевводеуглеводородыпроника- |
женииуглеводамиподцитоп- |
ют в клетку. |
|
|
лазматической мембраной |
выявляютсямногослойныемембранныеобразования- плазмолеммосомы, функция которых связана с синтезом капсульного материала.
Пристрессовыхфизическихихимическихвоздействияхнаклеткив них возникают патологические обратимые и необратимые изменения, касающиесяглавнымобразоммембранныхструктур. Повреждениеионизирующей радиацией, например, вызывает задержку лаг-фазы, в тече-
23
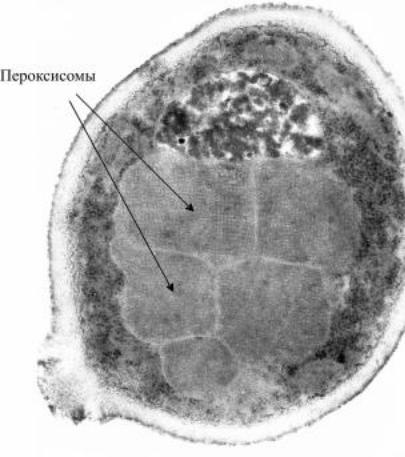
Рис. 10. Пероксисомы в дрожжевой клетке.
ние которой деление митохондрий нарушено. В результате образуются либогигантские, либооченьмелкиемитохондриисповрежденнымикристами. АппаратГольджиприэтомактивизируетсяи, какрезультат, появляютсямногочисленныефагосомы, дискретныевакуолиигранулы. Позженаблюдается появлениегигантскихклеток, вкоторыхгипертрофированы системы мембран Гольджи и ЭПР. Возникают многослойные концентрические зоны, клетка заполняется плотной сетью мембран. Ядро при облучении приобретает неправильную форму, в нем синтезируются внутриядерныемембраныиобразуетсятакназываемое«мозаичное» ядро, неспособное к делению.
При действии на дрожжи некоторых антибиотиков также происходят структурные изменения ядер. Стрептомицин вызывает их множественноеделение, чтоприводиткпоявлениюмногоядерныхклеток. Хлорамфеникол, наоборот, тормозит деление ядер и образуются гигантские
24
клеткисоченькрупнымиядрами. Циклогексимид, вызываядеструкцию нуклеиновыхкислот, снижаетэлектроннуюплотностьнуклеоплазмы. При действии нистатина на Candida albicans в клетках изменяется плазмолемма: на ней образуются глубокие трубковидные инвагинации.
Существенные структурные перестройки происходят в клетке при ее обезвоживании. В ядрах наблюдается конденсация хроматина на одном из полюсов и отчетливо выявляются хромосомы, обычно заметные лишь в профазе-анафазе митоза. Гомогенные участки ядра отделяются путем врастания нуклеолеммы. Поры в этой области ядра расширяются и происходит постепенная ресорбция этого участка. Хромосомыостаютсявсохраняющейсячастиядра. Жизнеспособностьприэтом нетеряетсяивысушенныеклеткилегкореактивируются. Выходизанабиотического состояния начинается с восстановления и реактивации митохондрий.
25
МОРФОЛОГИЯ И БЕСПОЛОЕ РАЗМНОЖЕНИЕ
При характеристике морфологии дрожжевых грибов обычно различаютмакроимикроморфологическиепризнаки, посколькупервыеизучаютвизуально, авторыеспомощьюмикроскопа. Макроморфология включает культуральные признаки, характеризующие рост культуры в жидкихилинаплотныхсредах. Макроморфологическиепризнакиочень изменчивы и сильно зависят от состава среды и условий культивирования, поэтому они имеют весьма ограниченное значение в систематике дрожжей. Темнеменее, многиевидыдрожжейсущественноотличаются по характеру роста на скошенном агаре (рис. 11) или в виде гигантской колонии (рис. 12), что на практике дает возможность их быстрой предварительной идентификации.
Рис. 11. Рост различных видов дрожжей на скошенном агаре.
Дрожжевые культуры, растущие на плотных средах, по консистенции бывают чаще всего пастообразными, а также слизистыми, иногда полностьюстекающиминаднопробирки, вязкими, клейкими, кожистымииликрошащимися. Слизистыйростхарактерендлямногиханаморфных базидиомицетовых дрожжей родов Cryptococcus, Rhodotorula,
26

Рис. 12. Гигантские колонии дрожжей разных видов на сусло-агаре.
Sporobolomyces, образующих большое количество внеклеточных полисахаридов, а также для аскомицетовых почвенных дрожжей рода Lipomyces. У большинства аскомицетовых дрожжей колонии пастообразные, сухие, культура при росте на скошенном агаре не стекает на дно пробирки. Для дрожжеподобных грибов,образующих как одиночные клетки, так и мицелий, характерны колонии с ворсинчатым краем, который хорошо просматривается при просвечивании.
У большинства дрожжей колонии белые, часто приобретающие при старении кремовый или слегка коричневатый оттенок. У некоторых аскоспоровых дрожжей, например из родов Nadsonia или Lipomyces, старые колонии при обильном спорообразовании темнеют и становятся бу-
27
рыми или шоколадными. Многие дрожжи образуют пигменты, окрашивающие их колонии в различные цвета. Наличие каротиноидных пигментов, придающих колониям красную, розовую, оранжевую или желтую окраску, характерно для базидиомицетовых дрожжей родов
Rhodotorula, Sporobolomyces, Cystofilobasidium и др. Аскомицетовые дрожжи Metschnikowia pulcherrima образуют диффундирующий в среду красно-вишневыйпигментпульхерримин. Такназываемые«черныедрожжи», формируют темно бурые или черные колонии за счет накопления меланиноподобных пигментов.
При росте в жидких средах дрожжи вызывают помутнение, образуют осадок, кольцо на стенках пробирки, разного характера пленки. Как правило, рост в виде пленки характеризует способность дрожжей объединяться в мицелиальные структуры или свидетельствует об окислительном типе энергетического метаболизма.
Микроморфология дрожжей Микроморфологиядрожжейвключаетпризнаки, характеризующие
отдельные клетки (форма, размеры), а также способы вегетативного и бесполого размножения и образуемые при этом структуры.
Типы вегетативного размножения. Морфогенез дрожжевой клет-
ки тесно связан со способом вегетативного размножения (рис. 13). Различают два принципиально различных способа образования вегетативныхклетокудрожжей- артрический(талломный) ибластический(зародышевый). При артрическом способе мицелий дрожжеподобных грибов одновременно распадается на отдельные одноклеточные элементы - артроспоры (рис. 14). Они образуются за счет расчленения гифы по поперечным септам после разрушения первичной стенки гифы в местах сочленения. Такой способ вегетативного размножения характерен для дрожжеподобныхгрибовEndomyces, Galactomyces, Arxula, Trichosporon,
причем у двух последних родов образование артроспор сопряжено с их последующим почкованием.
Бластический тип вегетативного размножения - это образование почек, что наиболее характерно для дрожжей. Почка представляет собой вырост на материнской клетке, который по мере увеличения в размерах
28
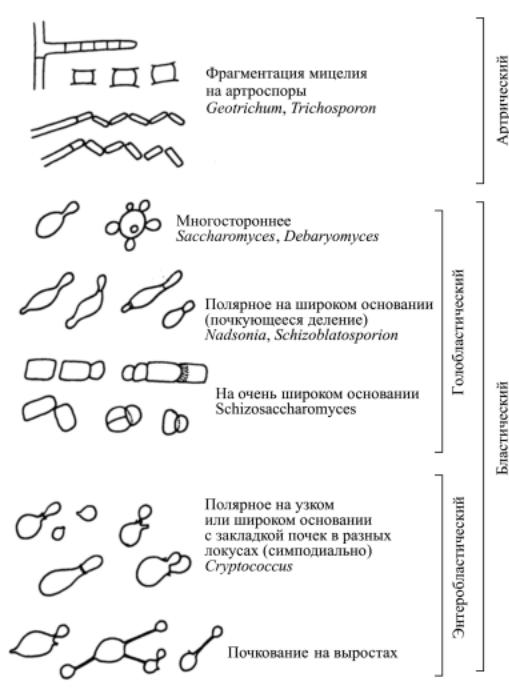
Рис. 13. Типы вегетативного размножения у дрожжей.
отшнуровываетсяотнее. Наматеринскойклеткеприэтомостаетсяшрам почкования, а на отделившейся почке - шрам рождения. Шрамы почкования (рис. 15), илипочечныерубцы, сохраняютсянаматеринскойклетке весь период ее жизни, а шрамы рождения со временем становятся малозаметными.
29
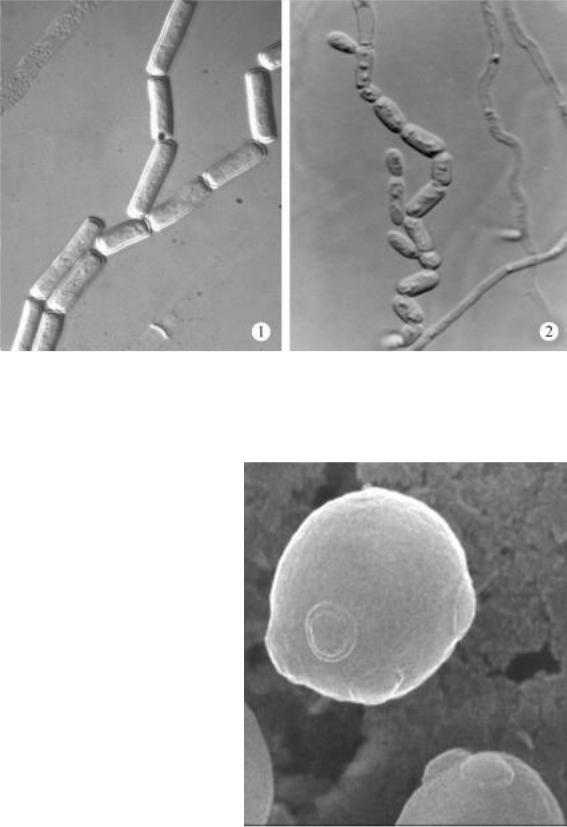
Рис. 14. Мицелий, распадающийся на артроспоры: 1 – Geotrichum candidum, 2 – Trichosporon asahii.
По ультраструктурному механизму образования почек различают голобласти-
ческое и энтеробластичес-
кое почкование (рис. 16). Первый тип характерен для аскомицетовых, второйдля базидиомицетовыхдрожжей, то есть признак этот имеет высокую таксономическую ценность. При голобластическомпочкованиивобразовании почки участвует вся клеточная стенка материнскойклетки, ипочкаприэтом как бы «выдувается» из нее. В какой-то степени такой
Рис. 15. Шрамы почкования на дрожжевой клетке.
30