

«Нервная ткань. Нейроны. Нейроглия»
1. Нервная ткань состоит из:
1) нервные клетки (нейроны, нейроциты),
2) глиальные клетки (нейроглия).
2. Функции нервной ткани:
1) восприятие, проведение и передача сигнала, полученного из внешней или внутренней среды, а затем к клеткам-мишеням.
3. Нервные клетки состоят из:
1) перикарион,
2) цитоплазматические отросткы (аксоны и дендриты).
4. Структурные особенности перикариона нейрона:
1) большой объем цитополазмы,
2) слабобазофильное ядро (содержит много эухроматина),
3) в ядре имеется одно или несколько ядрышек,
4) содержит органеллы общего назначения,
5) содержит специализированные органеллы – хроматофильное вещество, нейрофибриллы,
6) содержит включения (гликоген, липофусцин, пигментные).
5. Хроматофильное вещество цитоплазмы (субстанция Ниссля, тигроид, базофильное вещество):
1) это скопления цистерн грЭПС, где синтезируются нейромедиаторы,
2) проявляется при окрашивании метиленовым синим или толуидиновым синим,
3) имеет вид зернистости,
4) отсутствует в аксонном холмике и аксоне,
5) разрушение или распад глыбок базофильного вещества (тигролиз) наблюдается при реактивных изменениях нейронов, при повреждении и дегенерации.
6. Нейрофибриллы – это элементы цитоскелета (нейрофиламенты и нейротубулы), которые при импрегнации имеют вид нитей:
1) формируют остов нервной клетки,
2) имеют диаметр 8-10 нм, образованы фибриллярными белками,
3) основная функция – обеспечение стабильной формы нейрона и его отростков, транспорт органелл и нейромедиаторов.
7. Нейротубулы:
1) это микротрубочки диаметром около 24 нм, состоящие из молекул глобулярного белка тубулина,
2) выполняют опорную функцию,
3) полярны, отрицательно и положительно заряженные их концы позволяют контролировать диффузионно-транспортные потоки в аксоне (так называемый быстрый и медленный аксоток).
8. Дендриты – короткие отростки, ветвятся, в начальных сегментах содержат органеллы подобно телу нейрона, хорошо развит цитоскелет, несут потенциал действия от нервного окончания к телу нейрона.
9. Аксон (нейрит) – один, длинный, слабо ветвится или не ветвится, нейротубулы и нейрофиламенты располагаются упорядочено, в основном миелинизированы и окружены отростками олигодендроцитов в ЦНС, или леммоцитами в периферической нервной системе. Начальный сегмент аксона нередко расширен и имеет название аксонного холмика, где происходит суммация поступающих в нервную клетку сигналов, и формируется ответный потенциал действия, направляющийся вдоль аксона в синапс.
10. Аксоток (аксоплазматический транспорт веществ) - транспорт веществ в перикарионе и отростках нейрона, бывает:
1) от тела клетки на периферию (антероградный аксоток)
2) с периферии к центру (ретроградный аксоток).
11. По скорости аксоток бывает:
1) быстрый (со скоростью 100-1000 мм/сут), одинаков для всех нервных волокон; требует энергозатрат АТФ; происходит с участием транспортных пузырьков, содержащих медиаторы и модуляторы;
2) медленный (со скоростью 1-10 мм/сут), от центра к периферии клетки распространяются биологически активные вещества, а также компоненты биомембран и белков.
12. Классификация нейронов (морфологическая):
1) униполярные нейроны (с одним отростком),
2) псевдоуниполярные (спинномозговые ганглии),
3) биполярные нейроны — с одним аксоном и одним дендритом (сетчатка глаза, спиральный ганглий),
4) мульт иполярные нейроны — с множеством дендритов.
13. По функции (расположению в рефлекторной дуге) различают:
1) афферентные (чувствительные) нейроны – воспринимают информацию и передают ее в нервные центры (нейроны спинномозговых и черепно-мозговых узлов);
2) ассоциативные (вставочные) нейроны - осуществляют взаимодействие между нейронами, их большинство в ЦНС;
3) эфферентные (двигательные) нейроны - генерируют нервный импульс и передают сигнал другим нейронам или другим клеткам-мишеням (мышечным, секреторным).
14. Регенерация нейронов - внутриклеточная регенерация, потому что нейроны являются стабильной популяцией высокодифференцированных клеток, которые не делятся.
Нейроглия
15. Глиальные клетки (нейроглия) — клетки, окружающие нейроны; различают гистогенетических типа глиальных клеток:
1) макроглия – источник их развития нервная трубка,
2) микроглия – источник их развития гемопоэтическая стволовая клетка.
16. Клетки макроглии имеют разное строение в ЦНС:
1) астроциты (волокнистые и протоплазматические),
2) олигодендроциты,
3) эпендимоциты (в том числе и танициты).
17. Клетки макроглии периферической нервной системы:
1) сателлитоциты,
2) нейролеммоциты (шванновские клетки).
18. Функции макроглии:
1) изолирующая;
2) защитная (фагоцитоз микроглией),
3) трофическая,
4) секреторная,
5) метаболическая (метаболизм избытка нейромедиаторов в синапсе).
19. Астроциты – отростчатые клетки, многочисленные отростки которых ветвятся и окружают перикарон и дендриты нейронов, находятся только в органах ЦНС и анализаторах – производных нервной трубки. Разновидности астроцитов:
1) волокнистые – в белом веществе органов ЦНС,
2) протоплазматические – в сером веществе органов ЦНС.
20. Терминали отростков астроцитов - имеют вид расширений (ножки астроцитов), ножки соседних астроцитов плотно связаны между собой, формируя пластинки (глиальные мембраны):
1) вокруг гемокапилляров – периваскулярные,
2) вокруг тел нейронов – перинейрональные,
3) на поверхностях мозга – наружные и внутренние пограничные.
21. Специфический маркер астроцитов - глиальный фибриллярный кислый белок, из которого в них образуются промежуточные филаменты.
22. В постнатальном периоде жизни человека астроциты способны к миграции, особенно в зоны повреждения и способны к пролиферации (из них образуются доброкачественные опухоли астроцитомы).
23. Основные функции астроцитов:
1) формируют гистогематические барьеры в нервной ткани – гематоэнцефалический,
2) защитная, трофическая, регуляторная функции;
3) фагоцитоз погибших нейронов,
4) секреция биологически активных веществ: ФРФ, ангиогенные факторы, ЭФР, интерлейкин-1, простагландины.
24. Олигодендроциты – клетки с небольшим числом отростков, образуют миелиновую оболочку вокруг аксона нейронов. Олигодендроциты находятся в сером и белом веществе ЦНС, в периферической нервной системе.
25. Основные функции олигодендроцитов:
1) образуют миелиновую или безмиелиновую (нейролемму) оболочки вокруг нейронов, 2) изолирующая, трофическая, опорная, защитная функции;
3) участвуют в проведении нервного импульса, в регенерации поврежденных нервных клеток, фагоцитозе остатков осевых цилиндров и миелина при нарушении структуры аксона дистальнее места повреждения.
26. Эпендимоциты – клетки, по строению и архитектонике сходны с эпителем:
1) формируют непрерывный пласт, выстилающий полости мозга;
2) связаны друг с другом и базальной мембраной с помощью плотных, щелевидных и десмосомальных контактов;
3) апикальная поверхность содержит реснички и микроворсинки, участвующие в циркуляции ликвора;
4) базальная плазмолемма образует складки (инвагинации) или длинные тонкие отростки (таннициты), которые проникают до периваскулярных пространств микрососудов мозга.
27. Основные функции эпендимоцитов:
1) секреторная (образование ликвора),
2) защитная (селективная проницаемость гемато-ликворного барьера),
3) опорная,
4) регуляторная (в эмбриональном периоде развития предшественники таницитов – радиальные глиоциты, направляют миграцию нейробластов в нервной трубке).
28. Клетки микроглии (нейральные макрофаги) – клетки мезенхимного происхождения (производные моноцитов), диффузно распределенные в ЦНС в периваскулярном пространстве, имеют многочисленные сильно ветвящиеся отростки, способны к миграции, в цитоплазме много лизосом, гранул липофусцина.
29. Функция микроглии - защитная (фагоцитоз, участие в иммунных реакциях).
НЕРВНЫЕ ВОЛОКНА
1. Нервные волокна представляют собой отростки нейронов, покрытые глиальными оболочками. Различают два вида нервных волокон - безмиелиновые и миелиновые. Оба вида состоят из центрально лежащего отростка нейрона (осевого цилиндра), окруженного оболочкой из клеток олигодендроглии (в ПНС они называются леммоцитами или шванновскими клетками).
2. Безмиелиновые нервные волокна у взрослого располагаются преимущественно в составе вегетативной нервной системы и характеризуются сравнительно низкой скоростью проведения нервных импульсов (0.5-2 м/с). Они образуются путем погружения осевого цилиндра (аксона) в цитоплазму леммоцитов, располагающихся в виде тяжей. При этом плазмолемма леммоцита прогибается, окружая аксон, и образует дупликатуру - мезаксон. Нередко в цитоплазме одного леммоцита могут находиться до 10-20 осевых цилиндров. Такое волокно напоминает электрический кабель и поэтому называется волокном кабельного типа. Поверхность волокна покрыта базальной мембраной. В ЦНС, в особенности, в ходе ее развития, описаны безмиелиновые волокна, состоящие из "голого" аксона, лишенного оболочки из леммоцитов.
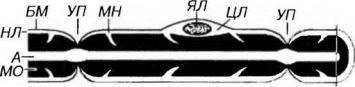
Рис. 14-8. Строение миелинового нервного волокна. Миелиновое волокно состоит из осевого цилиндра, или аксона (А), непосредственно окруженного миелиновой оболочкой (МО) и нейролеммой (НЛ), включающей цитоплазму (ЦЛ) и ядро леммоцита (ЯЛ). Снаружи волокно покрыто базальной мембраной (БМ). Участки МО, в которых сохраняются промежутки между витками миелина, заполненные ЦЛ и поэтому не окрашиваемые осмием, имеют вид миелиновых насечек (МН) МО отсутствует в участках, соответствующих границе соседних леммоцитов - узловых перехватах (УП).
3. Миелиновые нервные волокна встречаются в ЦНС и ПНС и характеризуются высокой скоростью проведения нервных импульсов (5-120 м/с). Миелиновые волокна обычно толще безмиелиновых и содержат осевые цилиндры большего диаметра. В миелиновом волокне осевой цилиндр непосредственно окружен особой миелиновой оболочкой, вокруг которой располагается тонкий слой, включающий цитоплазму и ядро леммоцита - нейролемма (рис. 14-8 и 14-9). Снаружи волокно также покрыто базальной мембраной. Миелиновая оболочка содержит высокие концентрации липидов и интенсивно окрашивается осмиевой кислотой, имея под световым микроскопом вид однородного слоя, однако под электронным микроскопом обнаруживается, что она возникает в результате слияния многочисленных (до 300) мембранных витков (пластин).
Образование миелиновой оболочки происходит при взаимодействии осевого цилиндра и клеток олигодендроглии с некоторыми различиями в ПНС и ЦНС.
Образование миелиновой оболочки в ПНС: погружение осевого цилиндра в леммоцит сопровождается формированием длинного мезаксона, который начинает вращаться вокруг аксона, образуя первые рыхло расположенные витки миелиновой оболочки (см. рис. 14-7). По мере увеличения числа витков (пластин) в процессе созревания миелина они располагаются все более плотно и частично сливаются; промежутки между ними, заполненные цитоплазмой леммоцита, сохраняются лишь в отдельных участках, не окрашиваемых осмием - миелиновых насечках (Шмидта-Лантермана). При формировании миелиновой оболочки цитоплазма и ядро леммоцита оттесняются к периферии волокна, образуя нейролемму. По длине волокна миелиновая оболочка имеет прерывистый ход.
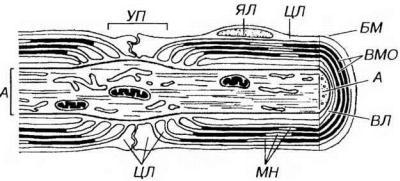
Рис. 14-9. Ультраструктурная организация миелинового нервного волокна. Вокруг аксона (А) располагаются витки миелиновой оболочки (ВМО), снаружи покрытые нейролеммой, а которую входят цитоплазма (ЦЛ) и ядро леммоцита (ЯЛ). Волокно окружено снаружи базальной мембраной (БМ). ЦЛ, помимо нейролеммы, образует внутренний листок (ВЛ), непосредственно прилежащий к А (расположенный между ним и ВМО), она содержится также в зоне, соответствующей границе соседних леммоцитов - узловом перехвате (УП), где миелиновая оболочка отсутствует, и в участках неплотной укладки ВМО - миелиновых насечках (МН).
4. Узловые перехваты (Ранвье) - участки в области границы соседних леммоцитов, в которых миелиновая оболочка отсутствует, а аксон прикрыт лишь интердигитирующими отростками соседних леммоцитов (см. рис. 14-9). Узловые перехваты повторяются по ходу миелинового волокна с интервалом, равным, в среднем, 1-2 мм. В области узлового перехвата аксон часто расширяется, а в его плазмолемме присутствуют многочисленные натриевые каналы (которые отсутствуют вне перехватов под миелиновой оболочкой).
5. Распространение деполяризации в миелиновом волокне осуществляется скачками от перехвата к перехвату (сальтаторно). Деполяризация в области одного узлового перехвата сопровождается ее быстрым пассивным распространением по аксону к следующему перехвату, (так как утечка тока в межузловом участке минимальна благодаря высоким изолирующим свойствам миелина). В области следующего перехвата импульс вызывает включение имеющихся ионных каналов и возникает новый участок локальной деполяризации и т.д.
Образование миелиновой оболочки в ЦНС: осевой цилиндр не погружается в цитоплазму олигодендроцита, а охватывается его плоским отростком, который в дальнейшем вращается вокруг него, теряя цитоплазму, причем его витки превращаются в пластинки миелиновой обо-
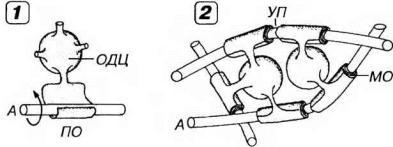
Рис. 14-10. Образование олигодендроцитами миелиновых волокон в ЦНС. 1 - аксон (А) нейрона охватывается плоским отростком (ПО) олигодендроцита (ОДЦ), витки которого превращаются в пластинки миелиновой оболочки (МО). 2 - один ОДЦ своими отростками может участвовать в миелинизации многих А. Участки А в области узловых перехватов (УП) не прикрыты цитоплазмой ОДЦ.
лочки (рис. 14-10). В отличие от шванновских клеток, один олигодендроцит ЦНС своими отростками может участвовать в миелинизации многих (до 40-50) нервных волокон. Участки аксона в области перехватов Ранвье в ЦНС не прикрыты цитоплазмой олигодендроцитов.
6. Нарушение образования и повреждение образованного миелина лежат в основе ряда тяжелых заболеваний нервной системы. Миелин в ЦНС может явиться мишенью для аутоиммунного поражения Т-лимфоцитами и макрофагами с его разрушением (демиелинизацией). Этот процесс активно протекает при рассеянном склерозе - тяжелом заболевании неясной (вероятно, вирусной) природы, связанном с расстройством различных функций, развитием параличей, потерей чувствительности. Характер неврологических нарушений определяется топографией и размерами поврежденных участков. При некоторых метаболических расстройствах возникают нарушения образования миелина - лейкодистрофии, проявляющиеся в детстве тяжелыми поражениями нервной системы.
7. Тип А имеет подгруппы:
- Аa — обладают наибольшей скоростью проведения возбуждения — 70-120 м/с (соматические двигательные нервные волокна);
- Аb — скорость проведения составляет 40-70 м/с. Это соматические афферентные нервы и некоторые эфферентные соматические нервы;
- Аg — скорость проведения составляет 15-40 м/с — афферентные и эфферентные симпатические и парасимпатические нервы;
- Аd (дельта) — скорость проведения 5-18 м/с. По этой группе афферентных соматических нервов проводятся первичная (быстрая) боль.
Тип В – скорость проведения от 3 до 14 м/с – преганглионарные симпатические волокна, некоторые парасимпатические волокна, то есть это вегетативные нервы.
Тип С – скорость проведения 0,5-3 м/с: постганглионарные вегетативные волокна (безмиелиновые). Проводят болевые импульсы медленной вторичной боли (от рецепторов пульпы зуба).
8. Нейрогенез - На 15-17 сутки внутриутробного развития человека под индуцирующим влиянием хорды из первичной эктодермы формируется нервная пластинка (скопление продольно лежащего клеточного материала). С 17 по 21 сутки пластинка инвагинирует и превращается сначала в нервный желобок, а затем в трубку. К 25 суткам эмбриогенеза происходит отщепление нервной трубки от эктодермы и замыкание переднего и заднего отверстий (нейропоров). По бокам от нервного желобка располагаются структуры нервного гребня.
9. На ранних сроках развития нервная трубка сформирована медулобластами – стволовыми клетками нервной ткани ЦНС. Из нервного гребня образуется ганглиозная пластинка состоящая из ганглиобластов – стволовых клеток нейронов и нейроглии периферической нервной системы. Медулобласты и ганглиобласты интенсивно иммигрируют, делятся и затем дифференцируются.
10. В ранние сроки внутриутробного развития нервная трубка представляет собой пласт отростчатых клеток, лежащих в виде одного слоя, но в несколько рядов. Изнутри и снаружи они ограничены пограничными мембранами. На внутренней поверхности (прилежащей к полости нервной трубки) медулобласты делятся.
11. В последующем нервная трубка формирует несколько слоев. Среди них можно выделить:
1) внутренняя пограничная мембрана: отделяет полость нервной трубки от клеток;
2) эпендимный слой (вентрикулярный в области мозговых пузырей) представлен бластными клетками-предшественниками макроглии;
3) субвентрикулярная зона (только в передних мозговых пузырях), где происходит пролиферация нейробластов;
4) мантийный (плащевой) слой, содержащий мигрирующие и дифференцирующиеся нейробласты и глиобласты;
5) маргинальный слой (краевая вуаль) сформирован отростками глиобластов и нейробластов. В ней можно видеть тела отдельных клеток.
6) наружная пограничная мембрана.
12. Диффероны нервной ткани центральной нервной системы:
- Дифферон нейрона: медулобласт – нейробласт – молодой нейрон – зрелый нейрон.
- Дифферон астроцита: медулобласт – спонгиобласт – астробласт – протоплазматический или волокнистый астроцит.
- Диферрон олигодендроцита: медулобласт — спонгиобласт – олигодендробласт – олигодендроцит.
- Дифферон эпендимной глии: медулобаст – эпендимобласт – эпендимоцит или таницит.
- Дифферон микроглии: стволовая клетка крови – полустволовая клетка крови (КОЕ ГЭММ) – КОЕ ГМ – КОЕ М – монобласт – промоноцит – моноцит – микроглиоцит покоя – активированный микроглиоцит.
13. Диффероны нервной ткани в периферической нервной системе:
- Дифферон нейрона: ганглиобласт – нейробласт – молодой нейрон – зрелый нейрон.
- Дифферон нейролеммоцита: ганглиобласт – глиобласт – нейролеммоцит (шванновская клетка).
14. Механизмы нейрогенеза -в процессе внутриутробного развития нейробласты мигрируют в области анатомических закладок нервных центров. При этом они прекращают делиться. В ЦНС миграция нейробластов контролируется адгезивными межклеточными взаимодействиями (с помощью кадгеринов и интегринов радиальной глии), сигнальными молекулами межклеточного вещества (в том числе фибронектинами и ламининами). После того как нейробласты достигают области своей постоянной локализации, они начинают дифференцироваться и формировать отростки. Направление роста отростков также контролируется упомянутыми адгезивными молекулами (кадгерины, интегрины, сигнальные молекулы межклеточного вещества).
15. Во внутриутробном развитии и после рождения происходит конкурентное взаимодействие между аналогичными нейронами нервных центров. При этом нервные клетки, не успевшие занять соответствующую зону, либо сформировать контакты, подвергаются апоптозу. В раннем развитии погибает от трети до половины нервных клеток.
16. В последующем развитии вокруг нервных клеток формируется глиальное окружение и происходит миелинизация нервных волокон. Нервные клетки до полового созревания продолжают формировать отростки и синаптические контакты. Максимального развития нервная ткань достигает к 25-30 годам.
17. С возрастом наблюдается гибель части нервных клеток и компенсаторная гипертрофия других. В нейронах может накапливаться липофусцин. Области с погибшими телами нервных клеток замещаются глиальными рубцам, образованными скоплением гипертрофированных астроцитов.
18. Дендриты сильно ветвятся, образуя дендритное дерево, и обычно короче аксона. От дендритов возбуждение направляется к телу нервной клетки. Они формируют постсинаптические структуры, воспринимающие возбуждение. Дендритов много, но может быть один.
19. Аксон присутствует всегда, по одному на каждую нервную клетку. Он не ветвится или слабо ветвится в терминальных областях и заканчивается синаптическим бутоном, передающим возбуждение на другие клетки (пресинаптическая зона). Нейроны передают возбуждение с помощью специализированных контактов (синапсов). Вещество, обеспечивающее передачу возбуждения, называется медиатором. В каждом нейроне обычно обнаруживается один основной медиатор.
20. Регенерация нервных волокон в периферической нервной системе
1) после перерезки нервного волокна проксимальная часть аксона подвергается восходящей дегенерации,
2) миелиновая оболочка в области повреждения распадается,
3) перикарион нейрона набухает,
4) ядро смещается к периферии,
5) хроматофильная субстанция распадается,
6) дистальная часть, связанная с иннервируемым органом, претерпевает нисходящую дегенерацию с полным разрушением аксона, распадом миелиновой оболочки и фагоцитозом детрита макрофагами и глией,
7) нейролеммоциты сохраняются и митотически делятся, формируя тяжи – ленты Бюнгнера,
8) через 4-6 недель структура и функция нейрона восстанавливается, от проксимальной части аксона дистально отрастают тонкие веточки, растущие вдоль лент Бюнгнера,
9) в результате регенерации нервного волокна восстанавливается связь с органом-мишенью.
21. При возникновении преграды на пути регенерирующего аксона (например, соединительнотканного рубца), восстановления иннервации не происходит.
22. МЕЖНЕЙРОННЫЕ КОНТАКТЫ (СИНАПСЫ)
Межнейронные контакты (синапсы) подразделяются на электрические и химические.
Электрические синапсы в ЦНС млекопитающих редки; они имеют строение щелевых соединений, в которых мембраны синаптически связанных клеток (пре- и постсинаптическая) разделены промежутком шириной 2 нм, пронизанным коннексонами. Последние представляют собой трубочки, образованные белковыми молекулами и служащие водными каналами, через которые мелкие молекулы и ионы могут транспортироваться из одной клетки в

Рис. 14-12. Строение химического синапса. Пресинаптическая часть (ПРСЧ) имеет вид концевого бутона (КБ) и включает: синаптические пузырьки (СП), митохондрии (МТХ), нейротрубочки (НТ), нейрофиламенты (НФ), пресинаптическую мембрану (ПРСМ) с пресинаптическим уплотнением (ПРСУ). В постсинаптическую часть (ПОСЧ) входит постсинаптическая мембрана (ПОСМ) с постсинаптическим уплотнением (ПОСУ). В синаптической щели (СЩ) находятся интрасинаптические филаменты (ИСФ).
другую. Когда потенциал действия, распространяющийся по мембране одной клетки, достигает области щелевого соединения, электрический ток пассивно протекает через щель от одной клетки к другой. Импульс способен передаваться в обоих направлениях и практически без задержки.
23. Химические синапсы - наиболее распространенный тип у млекопитающих. Их действие основано на преобразовании электрического сигнала в химический, который затем вновь преобразуется в электрический. Химический синапс состоит из трех компонентов: пресинаптической части, постсинаптической части и синаптической щели (рис. 14-12). В пресинаптической части содержится (нейро)медиатор, который под влиянием нервного импульса выделяется в синаптическую щель и, связываясь с рецепторами в постсинаптической части, вызывает изменения ионной проницаемости ее мембраны, что приводит к ее деполяризации (в возбуждающих синапсах) или гиперполяризации (в тормозных синапсах). Химические синапсы отличаются от электрических односторонним проведением импульсов, задержкой их передачи (синаптической задержкой длительностью 0.2-0.5 мс), обеспечением как возбуждения, так и торможения постсинаптического нейрона.
1.Пресинаптическая часть образуется аксоном по его ходу (проходящий синапс) или представляет собой расширенную конечную часть аксона (концевой бутон). В ней содержатся митохондрии, аЭПС, нейрофиламенты, нейротрубочки и синаптические пузырьки диаметром 20-65 нм, в которых находится нейромедиатор. Форма и характер содержимого пузырьков зависят от находящихся в них нейромедиаторов. Круглые светлые пузырьки обычно содержат ацетилхолин, пузырьки с компактным плотным центром - норадреналин, крупные плотные пузырьки со светлым подмембранным ободком - пептиды. Нейромедиаторы вырабатываются в теле нейрона и механизмом быстрого транспорта переносятся в окончания аксона, где происходит их депонирование. Частично синаптические пузырьки образуются в самом синапсе путем отщепления от цистерн аЭПС. На внутренней стороне плазмолеммы, обращенной к синаптической щели (пресинаптической мембраны) имеется пресинаптическое уплотнение, образованное фибриллярной гексагональной белковой сетью, ячейки которой способствуют равномерному распределению синаптических пузырьков по поверхности мембраны.
2.Постсинаптическая часть представлена постсинаптической мембраной, содержащей особые комплексы интегральных белков - синаптические рецепторы, связывающиеся с нейромедиатором. Мембрана утолщена за счет скопления под ней плотного филаментозного белкового материала (постсинаптическое уплотнение). В зависимости от того, является ли постсинаптической частью межнейронного синапса дендрит, тело нейрона или (реже) его аксон, синапсы подразделяют на аксо-дендритические, аксосоматические и аксо-аксональные, соответственно.
3.Синаптическая щель шириной 20-30 нм иногда содержит поперечно расположенные гликопротеиновые интрасинаптические филаменты толщиной 5 нм, которые являются элементами специализированного гликокаликса, обеспечивающими адгезивные связи пре- и пост-синатической частей, а также направленную диффузию медиатора.
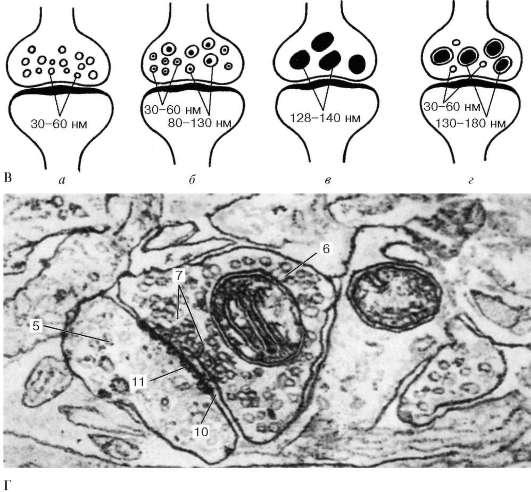
схема строения синаптических пузырьков: а - холинергических (светлых); б - адренергических; в - пуринергических; г - пептидергических (по Л. Д. Маркиной); Г - электронная микрофотография аксодендритного синапса (препарат И. Г. Павловой). 1 - аксосоматический синапс; 2 - аксодендритные синапсы; 3 - аксоаксональный синапс; 4 - дендриты; 5 - дендритный шипик; 6 - аксон; 7 - синаптические пузырьки; 8 - пресинаптическая мембрана; 9 - постсинаптическая мембрана; 10 - синаптическая щель; 11 - постсинаптические уплотнения
24. Механизм передачи нервного импульса в химическом синапсе. Под действием нервного импульса происходит активация потенциалзависимых кальциевых каналов пресинаптической мембраны; Са2+ устремляется в аксон, мембраны синаптических пузырьков в присутствии Са2+ сливаются с пресинаптической мембраной, а их содержимое (медиатор) выделяется в синаптическую щель механизмом экзоцитоза. Воздействуя на рецепторы постсинаптической мембраны, медиатор вызывает либо ее деполяризацию, возникновение постсинаптического потенциала действия и образование нервного импульса, либо ее гиперполяризацию, обусловливая реакцию торможения. Медиаторами, опосредующими возбуждение, например, служат ацетилхолин и глутамат, а торможение опосредуется ГАМК и глицином.
25.После прекращения взаимодействия медиатора с рецепторами постсинаптической мембраны большая часть его эндоцитозом захватывается пресинаптической частью, меньшая рассеивается в пространстве и захватывается окружающими глиальными клетками. Некоторые медиаторы (например, ацетилхолин) расщепляются ферментами на компоненты, которые далее захватываются пресинаптической частью. Мембраны синаптических пузырьков, встроенные в пресинаптическую мембрану, в дальнейшем включаются в эндоцитозные окаймленные пузырьки и повторно используются для образования новых синаптических пузырьков.
26. В отсутствие нервного импульса пресинаптическая часть выделяет отдельные небольшие порции медиатора, вызывая в постсинаптической мембране спонтанные миниатюрные потенциалы.