

1.5. Формообразование и везикулярный транспорт. Роль липидов и белков
До того, как перейти к описанию молекулярных механизмов регуляции везикулярного транспорта, важно, чтобы читатель имел в виду одно существенное обстоятельство, которое уже упоминалось во Введении. Рассматривая клеточные компартменты, можно найти относительно небольшое число форм (рис. 5).
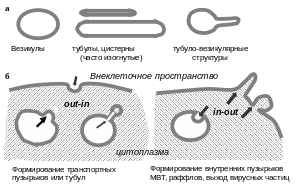
Рис. 5. Формы мембранных структур: (а) – встречающиеся в клетке; (б) – возможные направления инвагинации мембран
Действительно, несколько упрощая, можно сказать, что мембранные органеллы могут иметь форму везикулы, т. е. шара с кривизной поверхности, зависимой от диаметра пузырька (от 50 до 5000 нм), трубочки, или тубулы – цилиндрической замкнутой структуры, диаметр которой, как правило, сравним с диаметром самых маленьких везикул (50–100 нм), а длина существенно больше диаметра; уплощенной везикулы или тубулы (например, цистерны аппарата Гольджи или тубулы ЭПР). Везикулы могут отличаться по форме от идеального шара, часто значительно, а тубулы – различаться по длине и искривленности; кроме того, возможны различные комбинации этих форм, например, тубуло-везикулярные структуры довольно широко распространены в клетке.
Довольно долго считалось, что липидные мембраны приобретают и поддерживают ту или иную форму исключительно под влиянием мембранных белков. Однако эксперименты по формированию искусственных билипидных липосом с единым внутренним пространством, так называемые LUVs (Large Unilamellar Vesicles) показали, что варьируя липидный состав как внутреннего, так и наружного слоя, формируя LUVs в буфере с одним значением рН и перенося их затем в буфер с другой кислотностью, подвергая действию электрического тока или давлению, можно не только получать все наблюдаемые в живой клетке формы структур, но и превращения одних в другие. Более того, в определенных условиях можно было наблюдать на поверхности большой везикулы формирование маленьких везикул и их отпочкование – т. е. процесс, аналогичный формированию транспортных пузырьков. В других же условиях инициируется формирование инвагинаций с последующим отпочкованием маленьких пузырьков внутрь везикулы, т. е. именно то, что происходит при образовании мультивезикулярных тел или эндосом (МВТ или МВЭ) или упаковки секретируемых вирусных частиц на плазматической мембране (рис. 5). «С точки зрения клетки», в первом случае инвагинация мембраны происходит из компартмента в сторону цитоплазмы (out-in), а во втором – наоборот, из цитоплазмы в компартмент (in-out). Слияние LUVs также может происходить с определенной вероятностью, повышающейся с течением времени, без наличия белковой машинерии. Таким образом, теоретически, везикулярный транспорт липидных структур мог бы обойтись без всяких белков. Тем не менее, к настоящему времени уже известно несколько сотен белков, вовлеченных в везикулярный транспорт, и нужны они, в частности, для того, чтобы придать всем процессам специфичность и регулируемость.
Так, белки окаймлений определяют время и место формирования транспортных везикул, и что самое важное, выбор правильного груза; белки слияния определяют, с какой мембраной и когда сливаться транспортным везикулам, а малые ГТФазы Rab-семейства управляют и координируют все эти процессы. Координация везикулярного транспорта с процессами передачи сигнала осуществляют в основном сигнальные белки, но их субстратами могут быть не только белки транспортной машинерии, но и липиды. В результате везикулярный транспорт происходит с очень высокой точностью, и вероятность ошибочных слияний, также как и неверной сортировки, весьма низка, а уровень координации всех протекающих в клетке процессов очень высок.