

2.2.2. Основные компартменты эндоцитозного пути. Ранние, рециклирующие и поздние эндосомы. Мультивезикулярные тела (мвт). Лизосомы
Процесс формирования и отпочковывания от плазматической мембраны пузырька с грузом, в результате чего этот пузырек начинает свою самостоятельную жизнь в цитоплазме клетки, называется интернализацией. Судьба такого пузырька может быть различной и определяется рядом факторов, однако ключевыми являются природа груза и портал, использованный для входа. Основными компартментами эндоцитозного пути являются эндосомы, которые делятся на так называемые ранние и поздние (по времени появления в них интернализованных грузов), а также рециклирующие.
Как описывалось выше, несмотря на легкость визуальной идентификации основных компартментов биосинтетического пути, в вопросах его функционирования еще остается много неясного. В еще большей степени это относится к противоположному, эндоцитозному пути. Уже само название его главных компартментов ― «ранних» и «поздних» эндосом, основано не на каких-то характерных морфологических особенностях или локализации, а на времени попадания в них интернализованных белков-грузов, хотя и этот показатель весьма относителен. Так, в ранних эндосомах грузы выявляются уже через 5–20 мин после начала эндоцитоза, а в поздних могут обнаруживаться через 15–60 мин. Уже из приводимых цифр видно, что даже временные границы существования ранних и поздних эндосом неопределенны. Строго выполняется лишь одно условие – в поздние эндосомы интернализованный груз попадает из ранних.
Первоначально ранние эндосомы описывали как гетерогенную популяцию пузырьков, представленную отдельными везикулами диаметром до 200 нм, тубуло-везикулярными структурами (т. е. структурами, в которых от основной везикулярной части вытягиваются недлинные «рукоподобные» (arm-like) тубулярные выросты), а иногда – как довольно протяженную тубуло-везикулярную сеть. Кроме того, эндоцитированные грузы часто выявляли в так называемых мультивезикулярных телах, или эндосомах (МВТ, МВЭ) – крупных, диаметром до нескольких микрон, образованиях, содержащих разное количество внутренних пузырьков диаметром 50–100 нм. Их место и роль в эндоцитозном пути оставалась долго неясной, хотя эти структуры встречаются во всех клетках от дрожжей до млекопитающих. Даже конечный пункт деградационного пути – лизосомы, выглядящие как обычные везикулы, не вносили дополнительной ясности в определение границ компартментов и в механизмы переноса грузов между ними.
2.2.3. Общие принципы организации эндоцитозного пути
Несмотря на существование проблем идентификации компартментов эндоцитозного пути, многочисленные исследования позволили выявить некоторые закономерности (рис. 8).
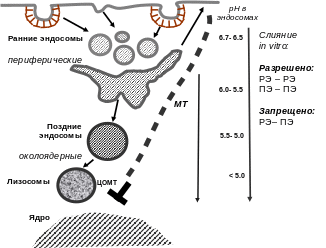
Рис. 8. Общая характеристика эндоцитозного пути. РЭ – ранние эндосомы; ПЭ – поздние эндосомы
Во-первых, эндоцитозный путь имеет векторную организацию – от мембраны к околоядерной области, – за счет того, что транспорт эндосом происходит по микротрубочкам, наиболее часто организованным в радиальную сеть с центром схождения в околоядерной области (им часто является ЦОМТ, центр организации микротрубочек). Ранние эндосомы располагаются преимущественно на периферии клетки. С течением времени грузы обнаруживаются в более крупных эндосомах, которые находятся в околоядерной области, расположенной в непосредственной близости от центра организации микротрубочек. Таким образом, для появления груза в поздних околоядерных эндосомах необходимы микротрубочки. Предполагалось, что околоядерные эндосомы ― это уже поздние эндосомы и их локализация облегчает взаимодействие с лизосомами, также расположенными преимущественно около ядра, рядом с ЦОМТ.
Во-вторых, было доказано, что по ходу эндоцитозного пути происходит закисление внутреннего пространства эндосом за счет работы протонной вакуолярной помпы, встроенной в мембрану эндосом.
В третьих, исследования слияния эндосом в системах in vitro показало, что существуют ограничения в слиянии эндосом друг с другом. Так, ранние эндосомы способны сливаться с ранними, а поздние ― с поздними, что говорит об общих свойствах их мембран (в этом случае мембраны называется гомотипичными), тогда как гетеротипические слияния (ранних эндосом с поздними) практически не происходят. Невозможность гетеротипических слияний отражает существенные различия в свойствах мембран ранних и поздних эндосом.