

8.2.1. Кратковременная и долговременная память
Образование памяти представляет собой ступенчатый процесс, начинающийся с поступления информации, закодированной и переработанной сенсорными системами (Справка 8.2). Новая информация в течение небольшого отрезка времени, около 20 – 30 секунд, сохраняется в кратковременной памяти, ёмкость которой незначительна: в среднем 72 элементов информации, т.е. семь слов, букв, цифр или символов. Приблизительно 20-30 секунд мозг обрабатывает эту информацию, решая, насколько она важна и стоит ли сохранять её в дальнейшем. Повторное поступление той же самой информации повышает вероятность её преобразования в долговременную память, ёмкость которой практически не ограничена, а длительность хранения варьирует от нескольких минут до десятилетий.
Представьте, что по справочному телефону вы узнали номер интересующего вас абонента: некоторое время вы можете удерживать в своей кратковременной памяти названную последовательность цифр, набирая её на своём телефонном аппарате. Но, если в это время что-нибудь отвлечёт на себя ваше внимание, вы можете позабыть ещё не записанный номер, не успевший поступить в долговременную память. Образованию следов долговременной памяти могут помешать травмы мозга или электрический шок, которые нередко сопровождаются потерей в памяти событий, непосредственно предшествовавших травме, поскольку поступившая информация не успевает в таких случаях преобразоваться из кратковременной формы в долговременную. Ступенчатый процесс образования памяти схематично можно представить как:
Поступление Воспроизведение
информации информации
Кратковременная память память
Долговременная
память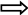

Регистрация
и извлечение
8.3. Нейронные механизмы памяти
Поступившее в центральную нервную систему возбуждение способно к непродолжительной циркуляции в замкнутых цепях нейронов, что называется реверберацией (Рис. 8.2). Потенциалы действия возвращаются к одним и тем же нейронам, сохраняя их возбуждённое состояние в течение некоторого времени: этим, как принято считать, обеспечивается кратковременная память. В возбуждённых нейронах фосфорилируются ферменты, способствующие кратковременному сохранению активности клеток. Электрошок, наркоз, контузия мозга, гипоксия (дефицит кислорода) и гипотермия (понижение температуры) препятствуют этому процессу, не позволяя кратковременной памяти преобразоваться в долговременную. Многократное повторяющееся действие раздражителей активирует одни и те же синапсы, что повышает эффективность их последующей деятельности. Стойкое изменение эффективности синаптической передачи означает консолидацию (от лат. consolidatio – укрепление) следов памяти и лежит в основе долговременной памяти.
8.3.1. Габитуация и сенситизация
Нейронные механизмы габитуации и сенситизации были установлены в лаборатории Эрика Кэндела (Kandel E.R.) при экспериментальных исследованиях защитного рефлекса у морского брюхоногого моллюска аплизии. Выбор именно этого объекта обусловлен наличием у него крупных нейронов, сгруппированных в нескольких ганглиях, что позволяет с уверенностью находить у разных особей однотипные сенсорные, моторные и интернейроны, а затем производить с ними все необходимые для исследования манипуляции, которые по многим причинам нельзя выполнить на нейронах человеческого мозга. Механизмы, осуществляющие синаптический перенос информации, однотипны у разных видов животных, поэтому обнаруженные у аплизии изменения эффективности работы синапсов, приводящие в итоге к возникновению новых форм поведения, считаются универсальными.
Аплизия рефлекторно прячет свой орган дыхания - жабру в специальную мантийную полость в ответ на раздражение самой мантии или кожи сифона, представляющего собой окружающее жабру возвышение, в толще которого находится мышца, втягивающая жабру (Рис. 8.3). Этот защитный рефлекс легко возникает у аплизии при стимуляции имеющихся в коже механорецепторов прикосновением или водяной струёй, выпущенной из резинового баллончика. При повторении раздражения через небольшие промежутки времени защитный рефлекс проявляется с каждым разом всё слабее и может исчезнуть совсем, что является одним из примеров габитуации.
Реакция привыкания у аплизии обусловлена инактивацией тока ионов кальция в пресинаптические окончания сенсорных нейронов. При повторяющихся раздражениях механорецепторов кожи сифона струёй воды или прикосновением всё меньше и меньше кальция входит в пресинаптические окончания сенсорных нейронов при их возбуждении. Следствием пониженной концентрации ионов кальция в пресинаптическом окончании сенсорного нейрона является уменьшение выделения медиатора из этого окончания и уменьшение постсинаптического потенциала, который становится ниже порогового. Открытие механизма габитуации продемонстрировало чрезвычайно простой способ образования долговременной памяти, связанный всего лишь с одним синапсом (гомосинаптическая депрессия).
Если на фоне габитуации будет использован какой-либо сильный раздражитель, действующий на другой участок тела (например, булавочный укол кожи хвоста или действие на него электрического тока), то реакция привыкания сразу же исчезнет (дегабитуация). После получения болевого стимула у аплизии развивается реакция сенситизации, выражающаяся в повышенном реагировании даже на безобидные тактильные стимулы, что наблюдается на протяжении достаточно долгого промежутка времени.
В реакции сенситизации кроме сенсорных и моторных нейронов участвуют интернейроны (Рис. 8.4), поэтому сенситизацию можно определить как гетеросинаптическое усиление. Реагирующий на булавочный укол сенсорный нейрон возбуждает интернейрон, который, в свою очередь, действует с помощью медиатора серотонина на пресинаптическое окончание другого сенсорного нейрона, передающего возбуждение от механорецепторов кожи сифона. Серотонин связывается с метаботропными рецепторами сенсорного нейрона, и вследствие этого в нём активируются вторичные посредники: цАМФ и цАМФ-зависимая протеинкиназа, которая фосфорилирует белки кальциевых мембранных каналов в пресинаптическом окончании. В результате при возбуждении в это окончание входит больше ионов кальция, соответственно больше выделяется глутамата, действующего на мотонейрон. Это приводит к увеличению амплитуды и длительности постсинаптических потенциалов, хотя количество и чувствительность постсинаптических рецепторов не изменяется. В экспериментах на изолированных нейронах для получения сенситизации не обязательно использовать болевой стимул: достаточно подействовать серотонином на пресинаптическое окончание сенсорного нейрона.