

5. Особенности протекания и варианты кроссинговера
Перекрест между гомологичными хромосомами может происходить не в одной, а одновременно в нескольких точках. Соответствующие названия кроссинговера: одинарный, двойной, тройной, …, множественный. Морган доказал это на дрозофиле, а затем эта закономерность подтвердилась на других организмах.
Чем больше расстояние между локусами, тем больше вероятность множественных перекрестов между ними и тем труднее их учитывать; они могут остаться незамеченными; нужен маркер в промежутке. И, наоборот, чем меньше расстояние между генами, тем точнее оно может быть определено.
В 1916 г. Г. Меллер открыл явление интерференции. Суть его в том, что кроссинговер, происшедший в одном участке, препятствует одновременному кроссинговеру в близлежащих участках хромосомы. Бывает положительная интерференция (когда препятствует) и, реже, - отрицательная (когда кроссинговер в одном локусе усиливает его в соседнем участке). Вблизи центромеры интерференция сильная.
Иногда при кроссинговере происходит обмен неравными участками – неравный кроссинговер. Впервые это установил в 1925 г. Альфред Хенри Стёртервант, изучая мутацию Bar – полосковидные глаза у самки дрозофилы. Соответствующий доминантный ген (В) находится в половой (Х) хромосоме.У самки дикого типа в глазу 800 фасеток, у гетерозиготы по гену В – 350, у гомозиготы – 70. Изредка встречается мутация «двойной» Bar (BB). В гетерозиготном состоянии он уменьшает число фасеток до 50, а в гомозиготном до 25. Стертевант предположил, что указанные различия связаны не с изменением самого гена, а с увеличением числа его копий за счет обмена гомологичных хромосом не совсем одинаковыми участками при кроссинговере.
Стертевант пометил Х – хромосому слева и справа от В двумя маркерными генами, провел скрещивания и доказал свое предположение, изучив фенотипы потомков.



 В В
В В

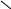
В


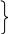

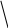 ВВ Кроссоверные
ВВ Кроссоверные
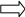
 В
В+
гаметы
В
В+
гаметы
В ВВ
Когда появилась возможность наблюдать строение гигантских хромосом у мух, то одновременно 3 группы ученых (Г. Меллер, А. А. Прокофьева – Бельговская и К. Бриджес) подтвердили цитогенетически, что у мух:
- дикого типа (В+) → один набор дисков на хромосоме;
- у мутантов В → два набора дисков на хромосоме;
- у «двойных» мутантов (ВВ) → три набора дисков на хромосоме.
Неравный кроссинговер наблюдали также на кукурузе.
Известен также митотический (соматический) кроссинговер. Он происходит с частотой ~10-4 в соматических клетках. Конъюгация происходит в интерфазе на стадии 4х хроматид. В митоз гомологичные хромосомы входят спаренными. Рентгеновское облучение может вызвать обмен участками несестринских хроматид.
Пример: у самки дрозофилы – гетерозиготы по двум аллелям окраски глаз w – белые глаза и wco – white corall (темно – красные глаза) – глаза бледно – розовые. После рентгеновского облучения таких самок (личинок) появляются на розовом фоне глаз белые и темно – красные пятна – результат соматического кроссинговера.