

3. Генетический аппарат эукариотических микроорганизмов (грибов).
Ядерный аппарат эукариотических клеток имеет ряд отличий от прокариотических: ДНК-содержащий компонент отделен от цитоплазмы ядерной оболочкой; количество ДНК в ядрах эукариот в тысячи раз больше, чем в составе нуклеоидов бактерий; ДНК эукариот представляет собой сложный нуклеопротеидный комплекс, образующий специальную структуру - хроматин, из которого и состоят эукариотические хромосомы. Далее, в состав ядер эукариот входят несколько физически не связанных хромосом, каждая из которых содержит одну линейную гигантскую молекулу ДНК.
Каждая хромосомная ДНК представляет собой полирепликонную структуру, т.е. содержит множеств автономно реплицирующихся участков. Синтез и образование транскриптов эукариотических клеток сопровождаются процессами вторичной их перестройки, «созревания», включающей в себя как фрагментацию (процессинг), так и сращивание отдельных фрагментов ДНК (сплайсинг). В ядрах не происходит синтеза белков, т.е. в эукариотических клетках процессы синтеза ДНК и РНК разобщены от процесса синтеза белков.
Клеточное ядро состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка и других продуктов синтетической активности, ядерного белкового остова (матрикса) и кариоплазмы (или ядерного сока). Эти основные компоненты встречаются практически во всех неделящихся клетках эукариотических одно- или многоклеточных организмов.
Главный компонент ядер — хроматин, является структурой, выполняющей генетическую функцию клетки, в хроматиновой ДНК заложена практически вся генетическая информация. Ядерная оболочка выполняет сложную барьерно-рецепторную, а также транспортную и каркасную функции. Нехроматиновый ядерный белковый остов (матрикс) обеспечивает не только пространственное расположение хромосом в ядре, но и участвует в их функциональной активности. Одним из хромосомных участков, определяющих синтез рРНК и образование клеточных рибосом, является ядрышко. Между всеми этими компонентами заключена жидкая фаза клеточного ядра - кариоплазма, в которой протекают многие процессы, связанные как с ядерным метаболизмом, так и с внутриядерным транспортом белков и РНК.
Для грибов характерна большая вариабельность в строении ядерного аппарата, его гетерогенность. У многоклеточных грибов может быть дикариотический и даже гетерокариотический ядерный аппарат. В последнем случае ядра одной клетки отличаются хромосомным составом, набор хромосом у грибов может быть как диплоидным, так и гаплоидным.
4. Лизогения у бактерий. Специфическая трансдукция: особенности и механизмы.
Умеренные бактериофаги вызывают лизогенизацию бактериальной клетки.
1. Нуклеиновая кислота умеренного фага интегрируется в геном бактериальной клетки, превращаясь в профаг. 2. Транскрипция профага репрессируется фаговым репрессором, вследствие чего информация с него не снимается и он реплицируется в составе генома лизогенной бактериальной клетки. Все клетки бактериальной культуры, ведущей начало от такой клетки содержат профаг. Поэтому и культура в целом называется лизогенной. 3. В дальнейшем с профага какой-либо клетки или совокупности клеток лизогенной культуры может начаться процесс снятия информации (такое явление называется индукция профага) и тогда разовьется продуктивная инфекция с лизисом клетки и выходом из нее зрелых корпускул умеренного бактериофага.
Лизогенное состояние устойчиво воспроизводится. Профаг при этом теряется с частотой около 1 на 10 —10 клеточных делений. В лизогенных культурах может происходить индукция бактериофага, в результате чего наблюдается массовый лизис бактерий. Такое явление происходит спонтанно и стимулируется целым рядом агентов, повреждающих ДНК ультрафиолетовыми и рентгеновскими лучами, алкилирующими соединениями, азотистым ипритом, органическими перекисями и т. д. Следует подчеркнуть, что, заражая бактериальную клетку, умеренный фаг может вызвать как литическую, так и лизогенную реакцию. Вероятность того и другого варианта зависит от физиологического состояния культуры.
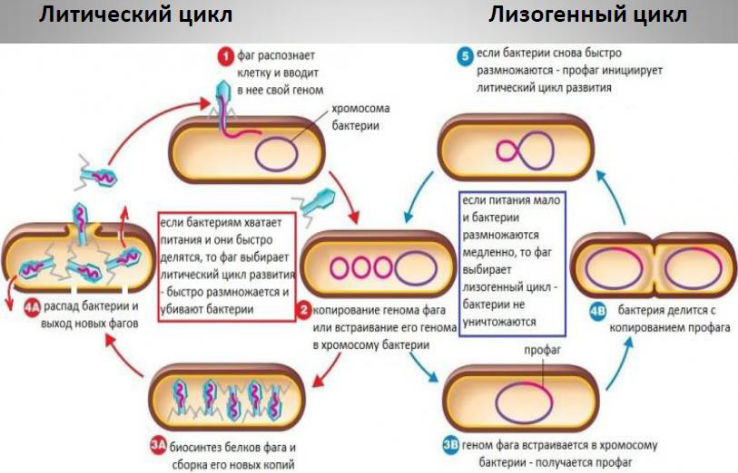
Трансдукция – передача бактериальной ДНК посредством бактериофага:
Специфическая трансдукция – перенос умеренным фагом определенного фрагмента ДНК (прилегающий к месту включения фаговой ДНК).
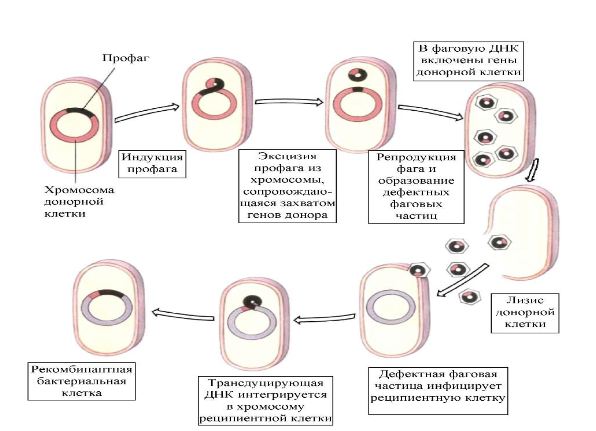
Общая (неспецифическая) трансдукция – перенос вирулентным бактериофагом фрагмента любой части бактериальной хромосомы.
Абортивная трансдукция – внесенный фрагмент ДНК донора не встраивается в хромосому реципиента, а остается в цитоплазме и там самостоятельно функционирует.