
Окисление жирных кислот
.pdfОкисление жирных кислот
Триглицериды являютсяоченьважным источником питательныхвеществиэнергиив животныхорганизмах. Онинакапливаютсяи сохраняютсявбольшихколичествахв жировой ткани.Впечени,сердце,ивскелетных мышцах свыше половиныэнергиипоставляют триглицериды.
Около95%всейдоступнойэнергиивмолекуле триглицеридовзаключаютвсебе остаткитрех жирныхкислотс длиннойцепью,а 5% приходятсянаглицерол. Клеткавкачестве источникаэнергииможетиспользоватьтолько свободные жирные кислоты,поэтому
триглицериды должны сперва расщепиться.Расщепление липидовназывается
.Гидролизтриглицеридовосуществляетфермент .Под действиемлипол зомтриглицеридлипазы происходит гидролизмолекулыЛипазатриглицерида,и образуютсяжирныекислоты,которыезатем окисляются.
Жирные кислоты в энергетическийобменмогутпопастьиз пищи,атакже из запасов жирав клетках. Впроцессе перевариванияпищевыелипидырасщепляются панкреатическойлипазой,ферментомподжелудочной железы,в тонком кишечнике. Продуктыгидролизалипидов —жирные кислоты с длинным углеводороднымрадикалом,2-моноацилглицеролы,холестерол,атакже соли жёлчныхкислотобразуютв просвете кишечникаструктуры,называемые смешаннымимицеллами.Мицеллы сближаютсясо щёточнойкаймойклеток слизистойоболочкитонкогокишечника,и липидные компонентымицелл диффундируют через мембраны внутрьклеток.Вместес продуктамигидролиза липидов всасываютсяжирорастворимые витамины A,D,Е,К исолижёлчныхкислот. После всасыванияпродуктовгидролизажиров жирные кислоты и2моноацилглицеролы в клеткахслизистойоболочкитонкого кишечникавключаются в процесс ресинтеза с образованиемтриацилглицеролов.
Липиды в воднойсреде(азначит,ив крови)нерастворимы,поэтомудлятранспорта липидов кровьюв организме образуютсякомплексы липидов сбелками— липопротеины. Жиры,образовавшиесяврезультате ресинтезав клетках слизистойоболочкикишечника,упаковываютсяв Хиломикроны,которые транспортируютсявкровьлимфатическойсистемой.Ониобеспечивают транспорт пищевыхлипидов откишечникадо печени ив другие ткани. В мышцахи жировойтканиониразрушаются липопротеин-липазой. ЛП-липазасвязанас гепарансульфатом(гетерополисахаридом), находящимсянаповерхности эндотелиальныхклеток, выстилающихстенкикапилляров кровеносныхсосудов. ЛП-липазатакже как ипанкреатическаялипаза, гидролизуетмолекулы жиров до глицеролаи 3молекул жирныхкислот. Поддействиемэтого ферментахиломикроны быстро теряютбóльшуючастьсвоихтриацилглицеринов.Остаткихиломикронов утилизируютсяпеченью. Основнаямассажирныхкислотпроникаетв ткани.В
ЛП-липазасвязанас гепарансульфатом(гетерополисахаридом), находящимсянаповерхности эндотелиальныхклеток, выстилающихстенкикапилляров кровеносныхсосудов. ЛП-липазатакже как ипанкреатическаялипаза, гидролизуетмолекулы жиров до глицеролаи 3молекул жирныхкислот. Поддействиемэтого ферментахиломикроны быстро теряютбóльшуючастьсвоихтриацилглицеринов.Остаткихиломикронов утилизируютсяпеченью. Основнаямассажирныхкислотпроникаетв ткани.В

жировой тканив абсорбтивныйпериоджирные кислоты депонируются в виде триацилглицеролов,в сердечноймышце иработающихскелетныхмышцах используются как источник энергии.Другойпродуктгидролизажиров,глицерол, растворимв крови,транспортируетсяв печень,где в абсорбтивныйпериодможет бытьиспользовандля синтезажиров.
Высвобождение жирныхкислотиз адипоцитов –гормонзависимыйпроцесс. Под действиемгормоновадреналинаиглюкагона активируютсяпротеинкиназы.Они активируют триглицерид липазу, котораяосуществляет гидролиз эндогенных триглицеридов. Из адипоцитов жирные кислоты попадаютв кровоток,где связываютсясальбумином кровиипереносятсяв другие ткани.Альбуминкак и липопротеины служитдлятранспортировкигидрофобныхлипидов по кровяному руслу.
Липопротеиновые комплексы представляютсобойшаровидные агрегаты, состоящие изядра,образованного неполярнымилипидами(триацилглицеринами и ацилхолестеринами), иоболочкитолщиной примерно 2нм,построеннойиз апопротеинов иамфифильныхлипидов (фосфолипидов ихолестерина).Наружная сторонаоболочкиполярна,вследствие этого липиды растворимы в плазме.Чем больше липидное ядро,т.е.чембольшуючастьсоставляютнеполярные липиды,тем
меньше плотностьлипопротеиновогокомплекса.
Поддействиемлипазобразуютсясвободные жирныекислоты,которыезатем
окисляются. |
|
|
матриксе митохондрий |
|
|
|||||
Окисление жирныхкислотпроисходитв |
|
β (бета)-Окисление,а продуктыих |
||||||||
специфическийпуть катаболизмажирныхкислот, прикотором от |
||||||||||
распадавключаютсяв цикллимоннойкислоты. |
|
|
|
|
— |
|||||
карбоксильногоконцажирнойкислотыпоследовательно отделяется по 2 |
||||||||||
атома углеродав виде ацетил-КоА. |
|
|
|
|
β |
β |
|
|||
|
|
|
|
Метаболическийпуть — -окисление —назван |
||||||
такпотому,что реакцииокисленияжирной кислоты происходят у |
|
-углеродного |
||||||||
атома (C-β3). |
|
|
|
|
|
|
|
|
||
Реакции |
-окисления ипоследующего |
|
|
|
|
|
|
|||
окисленияацетил-КоА вцикле Кребса |
|
|
|
|
|
|
||||
служатоднимизосновныхисточников |
|
|
|
|
|
|
||||
энергиидлясинтезаАТФпомеханизму |
|
|
аэробных |
|
|
|
||||
β |
|
|
|
|
|
|
|
|
|
|
окислительногофосфорилирования. |
|
|
|
|
условиях. |
|||||
|
-Окисление жирных кислотпроисходиттольков |
|
||||||||
Жирные кислоты поступаютвцитозольиздвухисточников: некоторыеа бумину. |
||||||||||
доставляются клеткамкровью,будучиприсоединеныксывороточному |
||||||||||
Отделившисьотнего онипроходятсквозьклеточные мембраныв цитозоль. |
||||||||||
Вторымисточником жирныхкислотслужатсодержавшиесяв самомцитозоле |
||||||||||
|
|
не способны |
|
|
|
|
|
|
|
|
триглицериды,расщепляющиесяпод действиемлипаз.Однако свободные жирные |
||||||||||
кислоты |
|
пройтичерезмитохондриальные мембраны. Онимогут |
||||||||
попастьв митохондриальныйматрикс, вкотором происходитих окисление,после тогокакподвергнутся трехэтапнымпревращениям:
1. Активация жирныхкислот |
|
|
Перед тем,каквступитьв различные реакции,жирные кислоты должныбыть |
||
активированы,т.е.связанымакроэргическойсвязью скоферментомА: |
||
- |
+ АМФ + PPi |
. |
R-COOH + HS-KoA + АТФ → R-CO- КоА |
|
|
Реакцию катализируетфермент .Выделившийсявходе
(ацильнаяпирофосг уппа -остаток жирнойкислоты)
реакции фат(PPi) гидролизуетсяацил-КоАферментомсинтетазапирофосфатазой:
Н4Р207 +Н20 2Н3Р04.
Выделение энергии→ пригидролизе макроэргическойсвязипирофосфатасмещает равновесие реакциивправоиобеспечивает полноту протеканияреакцииактивации.
Жирные кислоты с длинной цепью,которыепреобладаютв организме человека(от 12до 20атомов углерода),активируютсяацил-КоАсинтетазами,расположенными навнешнеймембране митохондрий.Жирные кислоты скороткойисреднейдлиной цепи(от4до 12атомов углерода)могутпроникатьв матрикс митохондрийпутём
диффузии.Активация этихжирныхкислотпроисходитв матриксе митохондрий. |
|||
2.Транспорт жирныхкислот с длиннаружнойуглеводородной ц пьювнутреннююмитохондриях. |
|||
Образовавшеесявещество,ацил-КоА,не можетпроходить через |
|||
мембрану митохондрий.Однакона |
поверхностиэтой внутренней |
||
мембранынаходится ферменткарнитин |
|
,катализирующий |
|
переносацильнойгруппына |
|
,собразованиемацилкарнитина. |
|
|
карнитин-ацилтрансфераза I |
|
|
ацил-SКoA + карнитин →ацил-карнитин+ КoA-SH
карнитин -аминокислота,природное вещество,родственное витаминамгруппы В. В отличие отвитаминов,карнитинможетсинтезиоватьсяв организмеиз незаменимыхаминокислотлизина иметионина. В медицине используетсядля коррекцииметаболическихпроцессов.Оказываетанаболическое,антигипоксическое
иантитиреоидное действие,активируетжировойобмен,стимулирует
Образоварегенерациювшийся,повышаетацилкарнитинспособенпроходитьчеаппетит. резмежмембранное пространство итранспортируется спомощью карнитинацилтранслоказына внутреннюю поверхностьвнутреннеймембраны митохондрий.
3. Образованиевнутримитохондриальной ацил-КоА.
Напоследнейстадии остатокжирнойкислоты(ацильнаягруппа) переноситсяот карнитина навнутримитохондриальныйКoA приучастии фермента
катализируетпереносацилана внутримитохондриальныйкарнитинацилтр нсферазаКоАII,.Такимкоторыйобразом,ацил-КоА становитсядоступным дляферментов β-окисления.
ацил-карнитин+ КoA-SH →ацил-SКoA + карнитин
ИТАК:
жирные кислотысдлинной углеводороднойцепью переносятся черезплотную внутреннюю мембрану митохондрийспомощью
карнитина
β-Окисление жирныхкислот — |
|
|
|
|
|
|
|
|
|
|
|
||
Этоспецифическийпутькатаболизма жирныхкислот, протекающийвматриксе |
|
||||||||||||
митохондрийтолько ваэробныхусловиях изаканчивающийсяобразованием |
|
||||||||||||
ацетил-КоА.Водород изреакций -окисленияпоступаетвЦПЭ,(цепь переноса |
|
||||||||||||
электронов), аацетил-КоАβокисляетсявцитратномцикле,также поставляющем |
|
||||||||||||
|
|
β |
|
|
|
|
|
|
|
|
|
|
|
водород дляЦПЭ. Поэтому -окисление жирныхкислот — важнейший |
|
|
|||||||||||
метаболическийпуть, обеспечивающийсинтезАТФвдыхательнойцепи. |
|
||||||||||||
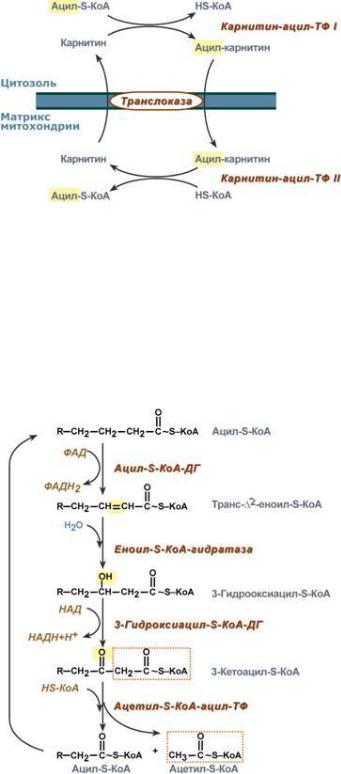
Окисление насыщенныхβ α |
жирныхкислот состоитих4 ферментативных реакций: |
|
|||||||||||
|
|
|
1 |
дег |
-Окисление начинаетсяс |
, |
|||||||
|
|
|
зависимой ацил-Кацил-КоА FAD- |
||||||||||
|
|
|
|
1. β |
дрирования |
|
|
||||||
|
|
|
|
сαобразованием двойнойсвязимежду |
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
Адегидрогеназой |
|
|
|
|
|
|
-и (IIиIII)атомамиуглерода. |
|
||||||||
|
|
|
|
Продуктреакции —еноил-КоА. |
|
||||||||
|
|
|
|
|
|
|
β- |
|
|
|
|
|
|
|
|
|
|
Восстановленныйвэтойреакции |
|
||||||||
|
|
|
2 |
кофермент FADH2 передаётпару |
|
||||||||
|
|
|
|
атомовводородаубихинону, |
|
||||||||
|
|
|
|
являющемусясоставнойчастью |
|
||||||||
|
|
|
|
митохондриальной дыхательной |
|
||||||||
|
|
|
3 |
цепи. |
|
|
|
|
|
|
|||
|
|
|
|
|
|
Вследующейреакции -окисления |
|
||||||
|
|
|
|
поместу двойнойсвязи |
β |
|
|||||||
|
|
|
|
2. |
|
|
|
|
|
|
|
||
|
|
|
|
присоединяетсямолекулаводы |
|
||||||||
|
|
|
|
такимобразом,чтоОН-группа |
|
||||||||
|
|
|
4 |
находится у -углеродногоатома |
|
||||||||
|
|
|
ацила,образуя -гидроксиацил-КоА. |
|
|||||||||
|
|
|
|
|
|
|
|
|
β |
|
|
|
|
|
|
|
|
|
. Затем -гидроксиацил-КоА |
|
|||||||
|
|
|
|
|
|
|
|
|
|
β |
|
|
|
|
|
|
|
окисляетсяNAD+-зaвисимой |
|
||||||||
|
|
|
|
3 |
|
|
|
β |
|
|
|
|
|
|
|
|
|
дегидрогеназой. Образуется β- |
|
||||||||
|
|
|
|
кетоацил-CoA и NADH+H+,который |
|
||||||||
передает восстановительные эквиваленты NADH -дегидрогеназе дыхательнойцепи. |
|
||||||||||||
|
.Образовавшийся -кетоацил-КоА подвергается тиолитическомурасщеплению |
|
|||||||||||
ферментом тиолазой, таккакпоместу разрывасвязи С—Счерез атомсеры |
|
||||||||||||
4 |
β |
|
|
|
|
|
|
|
|
|
|
|
|
присоединяетсямолекулакофермента-А.В результате этой последовательностииз |
|
||||||||||||
4реакцийотацил-КоА отделяетсядвухуглеродныйостаток —ацетил-КоА.Жирная |
|
||||||||||||

кислота,укороченнаяна2атомауглерода,опять проходит реакциидегидрирования, гидратации, дегидрирования,отщепленияацетил-КоА.Этупоследовательность реакцийобычноназывают«циклом -окисления»,имеяввиду,чтоодниите же реакцииповторяются срадикаломжирнойβ кислоты дотех пор, покавсякислотане превратится вацетил-Ко-А. . ХотяПродуктамиреакциикаждоговкаждомцикла«циклеβ»-окисленияодниите жеявля,остатокютсякислотыFADH , NADH,которыйи ацетилвходит-КоАв каждый последующийцикл, короче на2 углеродныхатома.В2 последнемцикле окисляетсяжирнаякислотаиз4атомовуглерода,поэтомуобразуются2молекулы ацетил-КоА,ане 1,каквпредыдущих.
например, дляокисления16углеродной пальмитиновойкислотытребуется7таких повторяющихсяциклов:
пальмитоил-КoA + 7CoA + 7FAD+ 7NAD+ + 7H 0 → 8 ацетил-КoA + 7FADH +
Из7NADHкаждой+ 7Hмолекулы+ FADH2 вдыхательной цепи2 образуется 2молекулыАТФ2 (14) КаждаямолекулаNADH образуетв дыхательнойцепи 3 молекулыАТФ (21) Каждаямолекулаацетил-КoA окисляетсявцикле кребсадоСО2 и Н2О иобразует12 молекулАТФ(96). Всего: приокислении. 16 углеродной пальмитиновойкислоты
образуется 131 молекул АТФ
Регуляцияскорости β-окисления
β-Окисление —метаболическийпуть,прочно связанныйс работойЦПЭиобщего путикатаболизма.Поэтомуегоскоростьрегулируетсяпотребностьюклеткив энергии,т.е.соотношениямиАТФ/АДФиNADH/NAD,так же,как искоростьреакций ЦПЭиобщего путикатаболизма.Скоростьβ-окисленияв тканяхзависитот доступностисубстрата,т.е.
отколичества жирныхкислот,поступающихв митохондрии. Концентрация свободныхжирныхкислотв кровиповышаетсяприактивации липолизав жировой тканиприголодании поддействиемглюкагонаи при физическойработепод действиемадреналина.В этихусловияхжирные кислотыстановятся преимущественнымисточникомэнергиидлямышципечени,так как в результате β- окисленияобразуютсяNADHиацетил-Ко А,ингибирующие пируватдегидрогеназный комплекс.Превращение пирувата,образующегося из глюкозы,в ацетил-КоА замедляется.Накапливаютсяпромежуточные метаболиты гликолиза и,в частности,глюкозо-6-фосфат.Глюкозо-6-фосфатингибируетгексокиназуи, следовательно,препятствуетиспользованиюглюкозыв процессегликолиза.Таким образом,преимущественное использование жирныхкислоткак основного источника энергиив мышечнойткании печенисберегаетглюкозудлянервнойткани и эритроцитов.
Скорость β-окислениязависиттакже отактивностифермента карнитинацилтрансферазы I.В печениэтотфермент ингибируетсямалонил-КоА, веществом,образующимсяприбиосинтезе жирныхкислот.В абсорбтивныйпериодв печениактивируетсягликолиз иувеличиваетсяобразование ацетил-КоАиз пирувата.Перваяреакциясинтезажирныхкислот — превращение ацетил-КоАв малонил-КоА.Малонил-КоАингибирует β-окисление жирныхкислот,которые могут использоватьсядлясинтезажира.
Окисление ненасыщенныхжирныхкислот
Около половины жирныхкислотв организме человеканенасыщенные, β-Окисление этихкислотидётобычнымпутёмдо техпор,покадвойнаясвязьне окажется
междутретьимичетвёртыматомамиуглерода.Затем ферментеноил-КоА изомеразаперемещаетдвойнуюсвязьиз положения3—4в положение 2-3иизменяет цис-конформациюдвойнойсвязинатранс-,котораятребуетсядля β-окисления.В этомцикле β-окисленияперваяреакциядегидрированияне происходит,так как двойнаясвязьв радикале жирнойкислоты уже имеется.Далее циклы β-окисления продолжаются,не отличаясьотобычного пути.
α-Окисление жирныхкислот.В липидахмозгаидругихотделахнервнойткани преобладаютжирные кислоты с оченьдлиннойцепью — более 20 углеродных атомов.Ониокисляютсяпо типу α-окисления,прикоторомотжирнойкислоты отщепляетсяпо одномуатомууглерода,выделяющемусяв виде С02. Этотпуть катаболизма жирных кислотне связанссинтезомАТФ.
ОБМЕНКЕТОНОВЫХТЕЛ |
|
|
|
|
|
|
|
|
|
Приголодании,длительнойфизическойработеивслучаях,когдаклеткинеполучают |
|||||||||
достаточногоколичестваглюкозы, жирные кислотыиспользуютсямногимитканямикак |
|||||||||
основнойисточникэнергии.Вотличиеотдругих тканей |
качествеисточникаэнергии. |
||||||||
Впеченичастьжирных кислотпревращаетсяв |
|
|
|
мозги другие отделынервной |
|||||
|
достаточное |
|
,которыеокисляются |
||||||
ткани практически не используют жирные кисло ыв |
|
количествоэнергиидля |
|||||||
мозгом,нервнойтканью,мышцами,обеспечивая |
|
|
|
||||||
|
|
|
кетоновые |
ла |
|
|
|||
синтезаАТФи уменьшаяпотреблениеглюкозы. Ккетоновымтеламотносят |
|||||||||
-гидроксибутират,ацетоацетатиацетон.Первыедве могутокислятьсявтканях, |
|||||||||
обеспечиваясинтезАТФ.Ацетонобразуетсятолькопривысоких концентрациях кетоновых |
|||||||||
β |
|
|
|
|
|
|
|
|
|
телвкровии,выделяясьсмочой,выдыхаемымвоздухомипотом,позволяеторганизму |
|||||||||
избавлятьсяотизбытка кетоновых тел. |
|
|
|
|
|
|
|
|
|
Смысл образованиякетоновых телзаключаетсявтом,чточастьацетил-КоА,полученного |
|||||||||
приокислении жирных кислотв клетках печени,избегает здесь дальнейшегоокисленияи |
|||||||||
направляется –вформекетоновых тел –вдругиеткани,гдеподвергаетсяокислениюдо |
|||||||||
СО2 иН2О.Образованиекетоновых тел - своегорода «перепускной»путь,которыйпечень |
|||||||||
используетдлятого,чтобынаправлятьклеточноетопливовдругиеобластитела. |
|||||||||
Прибольшомих накопленииразвивается |
|
|
|
,тяжелаяформакоторого |
|||||
представляетсерьезнуюугрозу дляжизни человекаиживотных.Рассматриватьэтот |
|||||||||
|
|
к т ацидоз |
|
|
|
|
|
||
процессможнокаку людей,такиуживотных. |
|
|
|
|
|
|
. |
||
Синтезкетоновых телпроисходит |
тольковмитохондрияхпечени |
||||||||
|
|
||||||||