

Лнтический
путь
ас.
Линейная
ДНК фага \t
проникающая
в клетку
Концы
цепей в одноцепочечных разрывах
сшиваются лигаэой
Двуна-^
праеленная
репликация
/
\\
ЛИЗО!
енный путь
Начало
синтеза ран них белков
Окончание
синтеза ранних- белков: начало синтеза
белков интеграции *
Им
let pdiиеная
рекомбинации
Хромосома
£.
coii
<5в
сМ Ы
т-г*г1-
С05
Gtt
blO
Начало
синтеза
поэдни*
белков
Окончание
син теза ранних
белкоь
]
'о
Катенирисанныв линейные геномы
Расщеплениэ
белком
А
/
Заполнение
г
о " о в к и
■
И
Ц Ь 
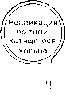


Конец ин let рации; 'переход к пизогенному состоянию
?
кой, необходимо функционирование как ранних так и пояттниу fphL 2?тия» который указан горизонтальной стрел- тикальной стрелкой, нужны функции лишь ранних генов. ' лиз°генного же пути, который указан вер-





in
t
Ей]
0
cl
m
о
T~
Q
Л
COS
Kb
26 27 28 29 30 31 32 33 34 35 36 37 30 39 40 4; 42 43 44 45 46 4-’ 46 49 5C
I
I I 1
I
I 1 I
Г
•
Т"Г"
~|—Г~1—г
I
I
Ш
Задержанные
ранние
-{
h
Немедпенные
ранние
©
Поздние
Установление
и
поддержание
лизогенного
состояния
'м
)
Репрессор
с I
Рис.
1.7. Схема регуляции генов бактериофага
К.
Прямоугольниками над картой указаны
те гены или сайты ДНК, которые участвуют
в регуляции активности генов фага К
(обсуждение функций этих генов см. в
тексте). ф
промоторы; >• продукты транскрипции;
tu
?ri и tn2~~
сайты
терминации, зависимой от фактора р; W
— продукт
гена
N,
который снимает терминацию в сайтах
tu
и Ы2\
Q
— продукт
гена Q, активатор
поздней транскрипции с промотора
pn'; —
транскрипты, которые кодируют генные
продукты, участвующие в установлении
и
поддержании лизогенного состояния.
SO
ГЛАВА 1
мым
выключает транскрипцию с промоторов
рь и рr.
К этому
времени, однако, образуется другой
регуляторный белок — Q,
причем
в количествах, достаточных для того,
чтобы началась транскрипция поздних
генов. Белок Q
действует
как положительный регулятор синтеза
РНК с промотора р\,
который используется для транскрипции
всей поздней области, содержащей
многочисленные гены, ответственные за
сборку головки и хвостового отростка
и за лизис клетки.
Сборка
Два
основных компонента зрелых частиц —
головка и хвостовой отросток —
собираются по отдельности.
Самым
ранним из выявленных при сборке головки
предшественником является «обнесенная
лесами» предголовка (рис. 1.8). Дальнейшее
созревание предголовок, в ходе которого
происходят удаление белка, выполняющего
функцию «лесов», и протеолити- ческое
расщепление других компонентов, зависит
от функционирования белка groE
бактерия-хозяина.
В результате получаются структуры,
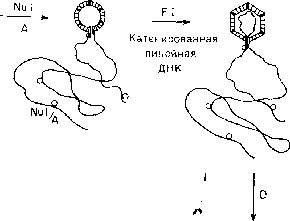
названные предголовками. На первом
этапе упаковки ДНК в прсдголовку
участвуют два белка, Nu\
и А,
которые связываются с катенированной
линейной ДНК вблизи левого сайта cos,
но не с самим этим сайтом. Затем такой
комплекс присоединяется к определенной
области предголовки. В присутствии
белка FI
ДНК укладывается внутри предголовки,
размер которой при этом увеличивается
примерно на 20%. Когда головка
оказывается заполненной, снаружи
капсида присоединяется белок D
(«decoration»
— белок «наружной отделки»), который
«обжимает» головку вокруг ДНК. В процессе
упаковки левый и правый сайты cos
на
катенированной линейной ДНК оказываются
рядом у входа в головку, где они и
расщепляются наискосок под действием
ter-функции
белка А
с образованием липких концов длиной
по 12 нуклеотидов. Наконец, заполненные
головки объединяются с независимо
образованными хвостовыми отростками. .
Лизис
Для
лизиса клетки-хозяина и высвобождения
потомства фага необходимы два фаговых
белка, R
и 5.
Особенно часто используются мутанты
по белку S,
так как
при этом внутри клеток накапливаются
большие количества фаговых частиц и
тем самым повышается выход фага.
Мутанты S~
можно
высвободить, лизируя клетки хлороформом.
Лизогенизация
В
небольшой части зараженных клеток не
развертывается литический цикл, а
активируется иной путь развития,
приводящий к интеграции ДНК фага К
в геном бактерии-хозяина. Вы-
f*
D=E
I
=B •
= Nu3
<5
=C
gro
e
«Обнесенная
лесами»
предголовка
(Белок
Сактерии-
хозяина)
/
••
I


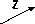


Предголовка
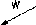

FH

ter
Функционирование белка А
Зрелая
фаговая
частица
Молекула
ДНК фага \
единичной
длины
Рис. 1.8. Схема сборки бактериофага к. Указаны белки, функционирующие на каждом этапе сборки (подробности см. в тексте и в работе Hohn, 1979). 4 F
о
•s
ч
м
£
Е
ю
п
>5
ч
о
V
X
О
со
33
X
со