

6 Мітохондрії
6.1 Які основні структурно-функціональні характеристики мітохондрій?
Мішечки округлої або витягнутої форми, стінка яких складається з двох мембран. Зовнішня мембрана гладка, має звичайну проникність. Внутрішня мембрана має виборчу проникність, в ній є випячування - крісти, в них вбудовани ферменти дихального ланцюга, ферментний комплекс АТФ-синтезу, транспортні білки.
Порожнина мітохондрії заповнена матриксом, який складається з безлічі ферментів (цикл Кребса, бета-окислення ліпідів та ін), рибосом, ДНК, РНК, проміжних продуктів розпаду жирних кислот і вуглеводів.
6.2 Які особливості будови мітохондрій?
Мітохондрії - найбільш відокремлені структурні елементи цитоплазми клітини, що володіють значною мірою самостійної життєдіяльністю. Існує навіть точка зору, що мітохондрії в історичному розвитку спочатку представляли собою самостійні організми, а потім проникали в цитоплазму клітин, де і ведуть сапрофітне існування. Про це свідчить, зокрема, той факт, що в мітохондріях є самостійний генетичний апарат (мітохондральной ДНК) і синтетичний апарат (мітохондріальні рибосоми). Однак зараз уже достовірно встановлено, що частина мітохондріальних білків синтезується в клітині.
6.2 Які функції мітохондрій?
Синтез молекул АТФ.
Джерелом утворення енергії в мітохондрії (її "паливом") є піровиноградна кислота (піруват), яка утворюється з вуглеводів, білків і ліпідів у гіалоплазме. Окислення пірувату відбувається в мітохондріальному матриксі в циклі трикарбонових кислот, а на крістах мітохондрій здійснюється перенесення електронів, фосфорилювання АДФ і утворення АТФ. АТФ, що утворюється в мітохондріях є єдиною формою енергії, яка використовується кліткою для виконання різних процесів.
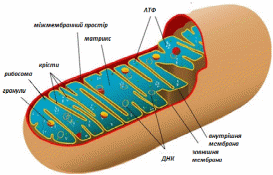
Малюнок 23 - мітохондрія (модель об'ємної реконструкції)
7 Лізосоми
7.1 Які основні структурно-функціональні характеристики лізосоми?
Мішечки, стінка яких складається з мембрани, всередині знаходяться гідролітичні ферменти (протеази, нуклеази, глікозідази, ліпази, фосфоліпази, сульфатази - більше 40 ферментів), що руйнують макромолекули - білки, вуглеводи і жири до низькомолекулярних продуктів, які можуть через мембрану дифундувати в цитозоль. Маркерним ферментом лізосом є кисла фосфатаза.
Усередині лізосом підтримується кисле рН, так як ферменти активні в кислому середовищі.
7.2 Назвіть види лізосом згідно функції, що вони виконують.
1. Первинні лізосоми (електроннощільні тільця) - неактивні;
2. Вторинні лізосоми фаголізосоми, в тому числі аутофаголізосоми (активні, що розщеплюють органічні речовини);
3. Третинні лізосоми або залишкові тільця, які містять неперетравлені продукти. Надалі видаляються з клітки екзоцитозу.
7.3 Опишіть травну функцію лізосом.
Травна функція лізосом починається тільки після злиття лізосоми з фагосомою (тобто фагоцитованої речовини, що оточена біліпідною мембраною). При цьому утворюється єдиний пухирець - фаголізосома, в якій змішуються фагоцитований матеріал і ферменти лізосоми. Після цього починається розщеплення (гідроліз) біополімерних з'єднань фагоцитованого матеріалу на мономірні молекули (амінокислоти, моносахара і так далі). Ці молекули вільно проникають через мембрану фаголізосоми в гіалоплазму і потім утилізуються клітиною, тобто використовуються або для утворення енергії, або на побудову біополімерних структур. Але не завжди фагоцитовані речовини розщеплюються повністю.
Доля решти речовин може бути різною. Деякі з них можуть бути виведені з клітини за допомогою екзоцитозу, за механізмом, який зворотній фагоцитозу. Деякі речовини (насамперед ліпідної природи) не розщеплюються лізосомальними гідролазами, а накопичуються і ущільнюються в фаголізосомах. Такі утворення називаються третинними лизосомами або залишковими тільцями. У процесі фагоцитозу і екзоцитозу здійснюється регуляція мембран в клітині: в процесі фагоцитозу частина плазмолеми відтшнуровиваєтся і утворює оболонку фагосоми, в процесі екзоцитозу ця оболонка знову вбудовується в плазмолему. Встановлено, що деякі клітини протягом години повністю оновлюють плазмолему.
Крім розглянутого механізму внутрішньоклітинного розщеплення фагоцитованих екзогенних речовин, таким же способом руйнуються ендогенні біополімери - пошкоджені або застарілі власні структурні елементи цитоплазми. Спочатку такі органели або цілі ділянки цитоплазми оточені біліпідною мембраною і утворюєть вакуоль аутофаголізосома, в якій здійснюється гідролітичне розщеплення біополімерних речовин, як і в фаголізосомах.
7.4 Які клітини містять багато лізосом?
Слід зазначити, що всі клітини містять в цитоплазмі лізосоми, але в різній кількості. Є спеціалізовані клітини (макрофаги), в цитоплазмі яких міститься дуже багато первинних і вторинних лізосом. Такі клітини виконують захисні функції в тканинах, вони спеціалізовані на поглинанні великої кількості екзогенних частинок (бактерій, вірусів), а також розпалися власних тканин.
7.5 Опишіть будову і функції пероксисом.
Пероксисоми - мікротільця цитоплазми, подібні за будовою з лизосомами, проте відрізняються від них тим, що в їх матриксі містяться кристалоподібні структури, а серед білків-ферментів міститься каталаза, що руйнує перекис водню, що утворюється при окисленні амінокислот.
7.6 Функція
Розщеплення фагоцитовані матеріалу.

Малюнок 24 - Загальна схема розщеплення фагоцитованого матеріалу

Рисунок 25 - Схема фагоцитозу
7.7 Що таке вакуолярная система?
Вакуолярная система, що складається з одномембранних різноманітних за будовою і функціями органел (ендоплазматичний ретикулум, апарат Гольджі, лізосоми, ендосоми, секреторні вакуолі), виконує загальну функцію синтезу, перебудови (модифікації), сортування та виведення (експорту) з клітки біополімерів, головним чином білків -глікопротеїдів, а також функцію синтезу мембран цієї системи і плазматичної мембрани.
Необхідно відзначити, що синтез основної маси клітинних білків протікає на полісомах в цитозолі. Особливістю білкового синтезу в цитозолі є те, що залежно від типу іРНК синтезуються різні білки, що прямують строго до своїх внутрішньоклітинних компонентів. Це пов'язано з тим, що різні за призначенням білки мають певні «сигнальні» послідовності амінокислот, як би адреси, за якими різні білки розподіляються в клітці. Так, ядерні білки мають NLS-сигнальну послідовність, білки мітохондрій, так само як білки цитозоля, цитоскелета, пластид і пероксисом, - свою сигнальну послідовність. Характерним є те, що всі типи перерахованих білків починають і закінчують синтез в цитозолі і потім посттрансляційної за допомогою внутрішньоклітинних білкових комплексів переносяться «за адресами».
На відміну від цих типів білків білки експортного призначення і білки мембран синтезуються на рибосомах, розташованих на мембранах ендоплазматичного ретикулума, і потрапляють всередину вакуолей, по мірі синтезу поліпептидного ланцюга, котрансляціонно. Потім ці білки вже всередині вакуолей або у складі мембран вакуолей транспортуються всередину клітини.
Загальна схема функціонування вакуолярної системи
На малюнку 26 представлені мембранні везикулярні компоненти, об'єднані в єдину функціональну систему. Всі вони мають загальну властивість: вони являють собою одномембранні компартменти, що мають єдине загальне джерело утворення (гранулярний ЕПР). Для всієї вакуолярної системи характерні кооперативность її функціонування, взаємозв'язок і послідовність етапів утворення, перебудови, транспорту та експорту синтезованих білків. Коротенько функції окремих компонентів полягають у наступному:
1. Гранулярний ЕПР: котрансляціонний синтез розчинних внутрівакуолярних білків (секреторні білки, гідролази лізосом та ін); котрансляціонний синтез нерозчинних білків, що входять до складу всіх мембран вакуолярної системи; первинна модифікація розчинних і нерозчинних (мембранних) білків, їх з'єднання з олігосахаридами - первинне гликозилирование синтезованих білків, освіта гликопротеидов; синтез мембранних ліпідів та їх вбудовування в мембрану - «збірка мембран».
2. Відділення вакуолей, що містять новостворені продукти, і їх перехід в цис-зону апарату Гольджі (ЕПР-АГ-комплекс).
3. Цис-зона апарату Гольджі: вторинна модифікація гликопротеидов; синтез полісахаридів (геміцелюлоза рослин) і гексозаміногліканов.
4. Проміжна зона апарату Гольджі: додаткові модифікації гликопротеидов, трансглікозілірованіе.
5. Транс-Гольджі-мережу: сортування секреторних і лізосомних білків; відділення вакуолей.
6. Екзоцитоз (секреція).
7. Екзоцитоз постійний.
8. Відділення первинних лізосом з гідролазами.
9. Ендоцитоз.
10. Вторинна лизосома.
11. Рециклизация рецепторів гидролаз.
12. Рециклизация рецепторів плазматичної мембрани.
13. Гладкий ендоплазматичнийретикулум: синтез і конденсація ліпідів, депонування іонів Са2 +, синтез і ресорбция глікогену та ін.
14. Транспорт в зону апарату Гольджі.
15. Транспорт від апарату Гольджі в ендоплазматичний ретикулум.

Малюнок 26 - Загальна схема вакуолярної системи клітини
1 - ядерна оболонка; 2 - шорсткогоЕПР (ЕПР); 3 - перехідна зона (ЕПР-АГ-комплекс); 4 - перенесення від ЕПР до апарату Гольджі (АГ); 5 - проксимальні ділянки (цис) АГ; 6 - середня частина (мед) АГ; 7 - дистальна (транс) частина АГ; 8 - транс-сетьАГ (TGN); 9 - поворотний шлях вакуолей АГ; 10 - відділення первинних лізосом; 11 - постійна екскреція (секреція); 12 - сигнальна секреція; 13 - ендоцитоз; 14 - ендосоми; 15 - вторинна лизосома; 16 - повернення лізосомних мембан в TGN; 17 - повернення рецепторів в плазматичну мембрану; 18 - гладкий ендоплазматичний ретикулум