

1. Ботаника в переводе с греческого растение, травы и представляет собой комплекс биологичских наук у растений. задача ботаники изучение строенияжизненных функций, распространения,происхождения, и эволюции растений.все люди и животные дышат кислородом и выделяют углекислый газ. Количество углекислого газа в воздухе также увеличивается от сжигания топлива. А растения в свою очередь поглощают углекислый газ из воздуха на свету и выделяют кислород.Кроме этого растения обогащают воздух кислородом, уменьшая количество углекислого газа. Ввиду того, что кислород является необходимым компонентом для жизни людей и животных, жизнь на Земле без зелёных растений была бы невозможна.Значение растений при образовании в них органических веществ играет также немаловажную роль. Зелёные растения создают органические вещества, а люди и животные для своего питания получают их готовыми от зелёных растений. Люди выращивают культурные растения, чтобы в дальнейшем собирать урожаи плодов фруктов, овощей, зёрен и т.д. и употреблять их в пищу, заготавливать на зиму. А для сельскохозяйственных животных собирают зерна, силос, которые также необходимые для жизни животных, потому что содержат питательные органические вещества. Животные не могли бы существовать без зелёных растений, так как питаются готовыми органическими веществами, которые в них образуются.значение растений в природе и жизни человека играет очень большую роль. Потому что зелёные растения дают людям пищу, сырьё для промышленности, а также корма для сельскохозяйственных животных.
2.Клетка - элементарная структурная, функциональная и генетическая единица всех живых организмов. наука о строении клетки, ее жихнедеятельности называют - цитологией.Представление о клетки возникло в связи с изобретением в 1590г голанд ученым Яном Янсенем микроскопа.Световой микроскоп остававшийся единственным инструментом изучения клетки на протяжении 350 лет. Впервые клеточное строение у растений наблюдал и описал Роберт Гук. В1831г Броун открыл ядро, а 1839 Ян Пуркинье рассматривает цитоплазму в качестве важнейшего компонента кл-ки. В 30-40х 19века немецкие ученые Шванн и Шлейден чформировали клеточную теорию суть которой состояла в тои, что клетка была признана основной структурной единицей всей живой природы. в 1858г Вирхов сформировал положение о том, что новые кл-ки образуются из материнских путем деления. в конце 19в ученый Флеммен открыл деление ядра и ввел точное понятие амитоз, митоз, кариогенез. 1880-1883 открыты хлоропласты 1886-1888 открыты хромосомы 1890- митохондрии 1898 аппарат Гольджи. 40е 20в создание электронного микроскопа в середине 20в позволило установить мембранное строение кл-ки.
3.В растительной клетке есть ядро и все органоиды, свойственные в животной клетке: эндоплазматическая сеть, рибосомы, митохондрии, аппарат Гольджи. Вместе с тем она отличается от животной клетки следующими особенностями строения:1) прочной клеточной стенкой значительной толщины; 2) особыми органоидами — пластидами, в которых происходит первичный синтез органических веществ из минеральных за счет энергии света — фотосинтез; 3) paзвитой системой вакуолей, в значительной мере обусловливающих осмотические свойства клеток.Растительная клетка, как и животная, окружена цитоплазматической мембраной, но, кроме нее, ограничена толстой состоящей из целлюлозы клеточной стенкой. Наличие клеточной стенки — специфическая Особенность растений. Она определила малую подвижность растений. Вследствие этого питание и дыхание организма стали зависеть от поверхности тела, контактирующей с окружающей средой, что привело в процессе эволюции к большей расчлененности тела, гораздо более выраженной, чем у животных. Клеточная стенка имеет поры, через которые каналы эндоплаэматической сети соседних клеток сообщаются друг с другом.Преобладание синтетических процессов над процессами освобождения энергии — одна из наиболее характерных особенностей обмена веществ растительных организмов. Первичный синтез углеводов из неорганических веществ осуществляется в пластидах.Различают три вида пластид: 1) лейкопласты — бесцветные пластиды, в которых из моносахаридов и дисахаридов синтезируется крахмал (есть лейкопласты, запасающие белки или жиры); 2) хлоропласты — зеленые пластиды, содержащие пигмент хлорофилл, где осуществляется фотосинтез — процесс образования органических молекул из неорганических за счет энергии света, 8) хромопласты, включающие различные пигменты из группы каротиноидов, обусловливающих яркую окраску цветков и плодов. Пластиды могут превращаться друг в друга. Они содержат ДНК и РНК, и увеличение их количества осуществляется делением надвое.Вакуоли окружены мембраной и рецэвиваются из эндоплазматичеокой сети. Вакуоли содержат в растворенном виде белки, углеводы, низкомолекулярные продукты синтеза, витамины, различные соли. Осмотическое давление, создаваемое растворенными в вакуолярном соке веществами, приводит к тому, что в клетку поступает вода, которая обусловливает тургор — напряженное состояние клеточной стенки. Толстые упругие стенки обеспечивают прочность растений к статическим и динамическим нагрузкам
4.цитоплазма это часть протопласта без ядра. основа цитоплазмы состовляет гиалоплазма- безцветнаяколойдная система обеспечивающая взаимодействие всех структур цитоплазмы. Гиалоплазма пронизана микротрубочками и микрофилоэлементами сборка и распад которых обеспечивающий из золя в гель. их совокупность состовляет цитоскелет.Цитоплазма (от греческого kytos — клетка и plasma — сформировавшееся) — это содержимое растительной или животной клетки, за исключением ядра (кариоплазмы). Цитоплазму и кариоплазму называют протоплазмой. В обычном микроскопе она выглядит как полужидкое вещество (основное вещество, или гиалоплазма), в котором взвешены разнообразные капельки, вакуоли, гранулы, палочковидные или нитевидные структуры. Под электронным микроскопом цитоплазма имеет еще более сложный вид (целый лабиринт мембран с заключенной между ними протоплазмой). Цитоплазма является сложной смесью белков, которые находятся в коллоидном состоянии, углеводов, жиров, нуклеиновых кислот и других органических соединений. Из неорганических соединений в цитоплазме присутствует вода, а также различные минеральные вещества.Снаружи каждая клетка окружена тончайшей плазматической мембраной (т. е. оболочкой), играющей важную роль в регуляции состава клеточного содержимого и являющейся производной цитоплазмы. Мембрана представляет собой трехслойную структуру (наружный и внутренний слои состоят из белка, между ними расположен слой фосфолипидных молекул) общей толщиной около 120 Å (ангстрем). Клеточная стенка пронизана мельчайшими отверстиями — порами, через которые протоплазма одной клетки может обмениваться с протоплазмой других, соседних клеток.В цитоплазме располагаются различные органоиды — специализированные структуры, выполняющие определенные функции в жизни клеток. Среди них важнейшую роль в обмене веществ играют митохондрии; в обычном микроскопе они видны в виде небольших палочек или зернышек. Данные электронной микроскопии указывают на их сложную структуру. Каждая митохондрия имеет оболочку, состоящую из трех слоев и внутренней полости. От оболочки в эту полость, заполненную жидким содержимым, вдаются многочисленные перегородки, не доходящие до противоположной стенки, называемые кристами. С митохондриями связаны дыхательные процессы. В цитоплазме имеется так называемая эндоплазматическая сеть (ретикулум) — разветвленная система субмикроскопических канальцев, трубочек и цистерн, ограниченных мембранами. Мембраны эндоплазматической сети двойные. На стороне, обращенной к основному веществу цитоплазмы, на каждой мембране расположены многочисленные гранулы, в состав которых входит рибонуклеиновая кислота, в соответствии с чем их стали называть рибосомами. При участии рибосом в эндоплазматической сети происходит синтез белков.Одним из компонентов цитоплазмы является сетчатый аппарат или «комплекс Гольджи», тесно связанный с эндоплазматической сетью и участвующий в процессах секреции. Имеются данные, показывающие, что мембраны ядра клетки (см.) без перерыва переходят в мембраны эндоплазматической сети и комплекса Гольджи. В цитоплазме некоторых животных клеток могут присутствовать фибриллы — тонкие нитевидные образования и трубочки, являющиеся сократительными элементами. Часто в цитоплазме видны зерна гликогена (у растений — крахмала), жировые вещества в виде мелких капель и другие структуры
5. Понятие об элементарной мембране. Классификация органелл по наличию мембран.
Понятие об элементарной биологической мембране Согласно современным представлениям, все биологические мембраны устроены сходным образом: основу мембраны составляет двойной молекулярный слой липидов (липидный бислой), по обе стороны и в толще которого находятся белки. Основные компоненты клетки подразделяют на мембранные и немембранные органеллы или органоиды (постоянные части клетки, имеющие определенное строение и выполняющие специфические функции). К мембранным органеллам относят цитоплазматическую сеть (эндоплазматический ретикулум), пластинчатый комплекс (аппарат Гольджи), митохондрии, лизосомы, пероксисомы. К немембранным относятся те органоиды, которые не имеют собственной замкнутой мембраны: рибосомы (полирибосомы), клеточный центр, элементы цитоскелета: микротрубочки и фибриллярные структуры. Биологическая мембрана - белково-липидная структура молекулярных размеров, расположенные на поверхности клетки или на поверхности внутриклеточных частиц ядра, митохондрий и др. Обладая избирательной проницаемостью, биологические мембраны регулируют в клетках концентрацию солей, сахаров, аминокислот и других продуктов обмена веществ. Модель строения элементарной биологической мембраны: 1 - молекулы липидов: а - гидрофильная "голова"б – гидрофобный "хвост"; 2 - молекулы белков: в - интегральная; г - периферическая;
Классификация органелл. Различают мембранные и немембраные органеллы. К мембранным органеллам относятся митохондрии, эндоплазматическая сеть, аппарат Гольджи, лизосомы. Немембранные органеллы: свободные рибосомы и полисомы, микротрубочки, центриоли и филаменты (микрофиламенты, промежуточные филаменты). Во многих клетках органеллы могут принимать участие в образовании особых структур, характерных для специализированных клеток. Так, реснички и жгутики образуются за счет центриолей и плазматической мембраны, микроворсинки — это выросты плазматической мембраны с гиалоплазмой и микрофиламентами, акросома спермиев — это производное элементов аппарата Гольджи, «эллипсоид» зрительных клеток — скопления митохондрий и пр.
6. Основные органеллы цитоплазмы, их строение и функции (гиалоплазма, плазмалемма, тонопласт, ЭПС, комплекс Гольджи, лизосомы, рибосомы, митохондрии).
Гиалоплазма - основное вещество цитоплазмы, заполняет все пространство между плазматической мембраной, оболочкой ядра и другими внутриклеточными структурами. Гиалоплазму можно рассматривать как сложную коллоидную систему, способную существовать в двух состояниях: золеобразном (жидком) и гелеобразном, которые взаимно переходят одно в другое. В процессе этих переходов осуществляется определенная работа, затрачивается энергия. Гиалоплазма лишена какой-либо определенной организации. Химический состав гиалоплазмы: вода (90%), белки (ферменты гликолиза, обмена сахаров, азотистых оснований, белков и липи-дов). Некоторые белки цитоплазмы образуют субъединицы, дающие начало таким органеллам, как центриоли, микрофиламенты.
Функции гиалоплазмы:
1) образование истинной внутренней среды клетки, которая объединяет все органеллы и обеспечивает их взаимодействие;
2) поддержание определенной структуры и формы клетки, создание опоры для внутреннего расположения органелл;
3) обеспечение внутриклеточного перемещения веществ и структур;
4) обеспечение адекватного обмена веществ как внутри самой клетки, так и с внешней средой.
Функции плазмолеммы:
· разграничивающая (барьерная);
· рецепторная или антигенная;
· транспортная;
· образование межклеточных контактов.
Основу строения плазмолеммы составляет двойной слой липидных молекулбилипидная мембрана, в которую местами включены молекулы белков, также имеется надмембранный слой гликокаликс, структурно связанный с белками и липидами билипидной мембраны, и в некоторых клетках имеется подмембранный слой.
ТОНОПЛАСТ — (от греч. tonos натяжение, напряжение и plastos оформленный, вылепленный), мембрана, ограничивающая вакуоль растит, клетки. Обладает избирательной проницаемостью, способна к активному транспорту веществ. Возможно участие Т. в образовании лизосом …
Эндоплазма-тическая сеть ЭС) |
Ультрамикроскопическая система мембран, образующих трубочки, канальцы, цистерны, пузырьки. Строение мембран универсальное (как и наружной), вся сеть объединена в единое целое с наружной мембраной ядерной оболочки и наружной клеточной мембраной. Гранулярная ЭС несет рибосомы, гладкая - лишена их |
Обеспечивает транспорт веществ как внутри клетки, так и между соседними клетками. Делит клетку на отдельные секции. в которых одновременно происходят различные физиологические процессы и химические реакции. Гранулярная ЭС участвует в синтезе белка. В каналах ЭС образуются сложные молекулы белка, синтезируются жиры, транспортируется АТФ |
Аппарат Гольджи |
Микроскопические одномембранные органеллы, состоящие из стопочки плоских цистерн, по краям которых ответвляются трубочки, отделяющие мелкие пузырьки |
В общей системе мембран любых клеток - наиболее подвижная и изменяющаяся органелла. В цистернах накапливаются продукты синтеза, распада и вещества, поступившие в клетку, а также вещества, которые выводятся из клетки. Упакованные в пузырьки, они поступают в цитоплазму: одни используются, .другие выводятся наружу. В растительной клетке участвует в построении клеточной стенки |
Лизосомы |
Микроскопические одномембранные органеллы округлой формы. Их число зависит от жизнедеятельности клетки и ее физиологического состояния. В лизосомах находятся лизирующие (растворяющие) ферменты, синтезированные на рибосомах |
Переваривание пищи, попавшей в животную клетку при фагоцитозе и пиноцитозе. Защитная функция. В клетках любых организмов осуществляют автолиз (саморастворение органелл), особенно в условиях пищевого или кислородного голодания. У животных рассасывается хвост. У растений растворяются органеллы при образовавании пробковой ткани. сосудов древесины |
Рибосомы |
Ультрамикроскопическис органеллы округлой или грибовидной формы, состоящие из двух частeй - субъединиц. Они не имеют мембранного строения и состоят из белка и рРНК. Субъединицы образуются в ядрышке. Объединяются вдоль молекулы иРНК в цепочки - полирибосомы - в цитоплазме |
Универсальные органеллы всех клеток животных и растений. Находятся в цитоплазме в свободном состоянии или на мембранах ЭС; кроме того, содержатся в митохондриях и хлоропластах. В рибосомах синтезируются белки по принципу матричного синтеза; образуется полипептидная цепочка - первичная структура молекулы белка |
Митохондрии |
Микроскопические органеллы, имеющие двухмембранное строение. Внешняя мембрана гладкая, внутренняя - образует различной формы выросты - кристы. В матриксе митохондрии (полужидком веществе) находятся ферменты, рибосомы, ДНК, РНК |
Универсальная органелла, являющаяся дыхательным и энергетическим центром. В процессе кислородного (окислительного) этапа диссимиляции в матриксе с помощью ферментов происходит расщепление органических веществ с освобождением энергии, которая идет на синтез АТФ (на кристах) |
|
|
|
7. Пластиды как органеллы, специфические для зеленых растений.
Пластиды являются основными цитоплазматическими органеллами клеток автотрофных растений. Название происходит от греческого слова «plastos», что в переводе означает «вылепленный».
Главная функция пластид – синтез органических веществ, благодаря наличию собственных ДНК и РНК и структур белкового синтеза. В пластидах также содержатся пигменты, обусловливающие их цвет. Все виды данных органелл имеют сложное внутреннее строение. Снаружи пластиду покрывают две элементарные мембраны, имеется система внутренних мембран, погруженных в строму или матрикс.
Классификация пластид по окраске и выполняемой функции подразумевает деление этих органоидов на три типа: хлоропласты, лейкопласты и хромопласты. Пластиды водорослей именуются хроматофорами.
Хлоропласты – это зеленые пластиды высших растений, содержащие хлорофилл – фотосинтезирующий пигмент. Представляют собой тельца округлой формы размерами от 4 до 10 мкм. Химический состав хлоропласта: примерно 50% белка, 35% жиров, 7% пигментов, малое количество ДНК и РНК. У представителей разных групп растений комплекс пигментов, определяющих окраску и принимающих участие в фотосинтезе, отличается. Это подтипы хлорофилла и каротиноиды (ксантофилл и каротин). При рассматривании под световым микроскопом видна зернистая структура пластид – это граны. Под электронным микроскопом наблюдаются небольшие прозрачные уплощенные мешочки (цистерны, или граны), образованные белково-липидной мембраной и располагающиеся в непосредственно в строме. Причем некоторые из них сгруппированы в пачки, похожие на столбики монет (тилакоиды гран), другие, более крупные находятся между тилакоидами. Благодаря такому строению, увеличивается активная синтезирующая поверхность липидно-белково-пигментного комплекса гран, в котором на свету происходит фотосинтез.
Хромопласты – пластиды, окраска которых бывает желтого, оранжевого или красного цвета, что обусловлено накоплением в них каротиноидов. Благодаря наличию хромопластов, характерную окраску имеют осенние листья, лепестки цветов, созревшие плоды (помидоры, яблоки). Данные органоиды могут быть различной формы – округлой, многоугольной, иногда игольчатой.
Лейкопласты представляют собой бесцветные пластиды, основная функция которых обычно запасающая. Размеры этих органелл относительно небольшие. Они округлой либо слегка продолговатой формы, характерны для всех живых клеток растений. В лейкопластах осуществляется синтез из простых соединений более сложных – крахмала, жиров, белков, которые сохраняются про запас в клубнях, корнях, семенах, плодах. Под электронным микроскопом заметно, что каждый лейкопласт покрыт двухслойной мембраной, в строме есть только один или небольшое число выростов мембраны, основное пространство заполнено органическими веществами. В зависимости от того, какие вещества накапливаются в строме, лейкопласты делят на амилопласты, протеинопласты и элеопласты.
Все виды пластид имеют общее происхождение и способны переходить из одного вида в другой. Так, превращение лейкопластов в хлоропласты наблюдается при позеленении картофельных клубней на свету, а в осенний период в хлоропластах зеленых листьев разрушается хлорофилл, и они трансформируются в хромопласты, что проявляется пожелтением листьев. В каждой определенной клетке растения может быть только один вид пластид.
8. Ядро. Физико-химические особенности. Ядерная оболочка, ядерный сок. Ядрышко, его образование, строение, функции. Форма, число и размеры ядер. Функции ядра.
Ядро (лат. nucleus) — это один из структурных компонентов эукариотической клетки, содержащий генетическую информацию (молекулы ДНК), осуществляющий основные функции: хранение, передача и реализация наследственной информации с обеспечением синтеза белка. Содержит различные ферменты, нуклеотиды, аминокислоты и другие вещества, необходимые для обеспечения синтеза нуклеиновых кислот и субъединиц рибосом, транспортируемых из ядра в цитоплазму. В ядерном соке находятся также нитевидные белковые молекулы, составляющие ядерный матрикс, который, подобно цитоскелету в цитоплазме,
От цитоплазмы ядро отделено ядерной оболочкой, образованной за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов. Полость ядерной оболочки называется люменом илиперинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. Основу ядерного сока, или матрикса, составляют белки. Ядерный сок образует внутреннюю среду ядра, в связи с чем он играет важную роль в обеспечении нормального функционирования генетического материала. В составе ядерного сока присутствуют нитчатые, или фибриллярные, белки, с которыми связано выполнение опорной функции: в матриксе находятся также первичные продукты транскрипции генетической информации
Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специальные участки, так называемые ядрышковые организаторы, содержащие генырибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации (NoLS, от англ. Nucleolus Localization Signal). Следует отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда неспецифически.
Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц.
Большинство клеток имеют одно ядро, изредка встречаются двухъадерные (клетки печени) и многоядерные (многие водоросли, грибы, млечные сосуды растений, поперечнополосатые мышцы). Некоторые клетки в зрелом состоянии не имеют ядра (например, эритроциты млекопитающих и клетки ситовидных трубок у цветковых растений).
Форма и размеры ядра клетки очень изменчивы и зависят от вида организма, а также от типа, возраста и функционального состояния клетки. Ядро может быть шаровидным (5—20 мкм в диаметре), линзовидным, веретеновидным и даже многолопастным (в клетках паутинных желез некоторых насекомых и пауков).
9. Клеточная стенка (клеточная оболочка) – характерный признак растительной клетки, отличающий ее от клетки животной. Клеточная стенка придает клетке определенную форму. Клеточная стенка придает клетке прочность и защищает протопласт, она уравновешивает тургорное давление и препятствует, таким образом, разрыву плазмалеммы. Совокупность клеточных стенок образует внутренний скелет, поддерживающий тело растения и придающий ему механическую прочность. Клеточная стенка бесцветна и прозрачна, легко пропускает солнечный свет. Обычно стенки пропитаны водой. По системе клеточных стенок осуществляется транспорт воды и растворенных в ней низкомолекулярных соединений (транспорт по апопласту). Клеточная стенка состоит в основном из полисахаридов, которые можно подразделить на скелетные вещества и вещества матрикса. Скелетным веществом клеточной стенки растений является целлюлоза (клетчатка), представляющая собой бета-1,4-D-глюкан. Это самое распространенное органическое вещество биосферы. Молекулы целлюлозы представляют собой очень длинные неразветвленные цепи, они располагаются параллельно друг другу группами по нескольку десятков и скреплены многочисленными водородными связями. В результате образуются микрофибриллы, которые создают структурный каркас стенки и обусловливают ее прочность. Микрофибриллы целлюлозы видны только в электронный микроскоп. Целлюлоза нерастворима и не набухает в воде. Микрофибриллы целлюлозы эластичны и очень прочны на разрыв (сходны со сталью . У грибов скелетным веществом клеточной стенки является хитин – полисахарид, построенный из остатков глюкозамина. Хитин еще более прочен, чем целлюлоза. Микрофибриллы погружены в аморфный матрикс, обычно представляющий собой насыщенный водой пластичный гель. Матрикс является сложной смесью полисахаридов, молекулы которых состоят из остатков нескольких различных сахаров и представляют собой более короткие, чем у целлюлозы, и разветвленные цепи. Матричные полисахариды определяют такие свойства клеточной стенки, как сильная набухаемость, высокая проницаемость для воды и растворенных в ней низкомолекулярных соединений. Полисахариды матрикса делят на две группы – пектиновые вещества и гемицеллюлозы.
10. Пектиновые вещества сильно набухают или растворяются в воде. Они легко разрушаются под действием щелочей и кислот. Простейшими представителями пектиновых веществ являются растворимые в воде пектовые кислоты – продукты полимеризации .Пектиновые кислоты, пектаты и пектинаты растворимы в воде в присутствии сахаров и органических кислот с образованием плотных гелей. Гемицеллюлозы представляют собой разветвленные цепи, построенные из остатков нейтральных сахаров, чаще встречаются глюкоза, галактоза, манноза, ксилоза; степень полимеризации 50-300. Гемицеллюлозы химически более устойчивы, чем пектиновые вещества, они труднее гидролизуются и слабее набухают в воде. Клеточные стенки растений часто подвергаются химическим видоизменениям. Одревеснение,происходит в том случае, если в матриксе откладывается лигнин – полимерное соединение фенольной природы, нерастворимое в воде. Одревесневшая клеточная стенка теряет эластичность, резко повышается ее твердость и прочность на сжатие, снижается проницаемость для воды. Опробковение, происходит в результате отложения с внутренней стороны клеточной стенки гидрофобных полимеров – суберина и воска. Суберин представляет собой смесь эфиров полимерных жирных кислот. Мономерами воска являются жирные спирты и восковые эфиры. Воск легко извлекается органическими растворителями и быстро плавится, образует кристаллы. Суберин – аморфное соединение, не плавится и не растворяется в органических растворителях. Суберин и воск, образуя чередующиеся параллельные слои, выстилают всю полость клетки с внутренней стороны в виде пленки. Субериновая пленка практически непроницаема для воды и для газов, поэтому после ее образования клетка обычно отмирает. Опробковение характерно для стенок клеток покровной ткани пробки. Кутинизации подвергаются наружные стенки клеток покровной ткани эпидермы. Кутин и воск откладываются чередующимися слоями на наружной поверхности клеточной стенки в виде пленки – кутикулы. Кутин представляет собой жироподобное полимерное соединение, близкое по химической природе и свойствам суберину. Кутикула предохраняет растение от излишнего испарения воды с поверхности растения. Окрасить ее можно реактивом судан III в оранжево-красный цвет. Минерализация клеточной стенки происходит вследствие отложения в матриксе большого количества минеральных веществ.Минеральные вещества придают стенке твердость и хрупкость.
Клеточная стенка является продуктом жизнедеятельности протопласта. Полисахариды матрикса, гликопротеид стенки, лигнин и слизи образуются в аппарате Гольджи. Синтез целлюлозы, образование и ориентация микрофибрилл осуществляются плазмалеммой. Большая роль в ориентации микрофибрилл принадлежит микротрубочкам, которые располагаются параллельно откладывающимся микрофибриллам вблизи плазмалеммы. Если микротрубочки разрушить, образуются только изодиаметрические клетки.
11
Вакуо́ль — одномембранный органоид, содержащийся в некоторых эукариотических клетках и выполняющий различные функции (секреция, экскреция и хранение запасных веществ, аутофагия, автолиз и др.). Вакуоли развиваются из мембранных пузырьков — провакуолей. Провакуоли являются производными эндоплазматического ретикулума и комплекса Гольджи, они сливаются и образуют вакуоли. Вакуоли и их содержимое рассматриваются как обособленный от цитоплазмы компартмент. Различают пищеварительные и сократительные (пульсирующие) вакуоли, регулирующие осмотическое давление и служащие для выведения из организма продуктов распада. Вакуоли особенно хорошо заметны в клетках растений: во многих зрелых клетках растений они составляют более половины объёма клетки, при этом они могут сливаться в одну гигантскую вакуоль. Одна из важных функций растительных вакуолей — накопление ионов и поддержание тургора (тургорного давления). Вакуоль — это место запаса воды.
Мембрана, в которую заключена вакуоль, называется тонопласт, а содержимое вакуоли — клеточный сок. Клеточный сок состоит из воды и растворенных в ней веществ, а также из моносахаридов, дисахаридов, танинов,углеводов, неорганических веществ (нитраты, фосфаты, хлориды и др.) и органических кислот. Тургор(позднелат. turgor — вздутие, наполнение, от лат. turgere — быть набухшим, наполненным)напряжённое состояние клеточной оболочки, зависящее от осмотического давления внутриклеточной жидкости , осмотическое давления внешнего раствора и упругости клеточной оболочки.
Плазмолиз-отделение протопласта отклеточной стенки в гипертоническом растворе.
Плазмолизу предшествует потеря тургора.
Плазмолиз возможен в клетках, имеющих плотную клеточную стенку (у растений, грибов, крупных бактерий[1]). Клетки животных, не имеющие жесткой оболочки, при попадании в гипертоническую среду сжимаются, при этом отслоения клеточного содержимого от оболочки не происходит. Характер плазмолиза зависит от ряда факторов:
от вязкости цитоплазмы;
от разности между осмотическим давлением внутриклеточной и внешней среды;
от химического состава и токсичности внешнего гипертонического раствора;
от характера и количества плазмодесм;
от размера, количества и формы вакуолей.
Различают уголковый плазмолиз, при котором отрыв протопласта от стенок клетки происходит на отдельных участках. Вогнутый плазмолиз, когда отслоение захватывает значительные участки плазмалеммы, и выпуклый, полный плазмолиз, при котором связи между соседними клетками разрушаются практически полностью. Вогнутый плазмолиз часто обратим; в гипотоническом растворе клетки вновь набирают потерянную воду, и происходит деплазмолиз. Выпуклый плазмолиз обычно необратим и ведет к гибели клеток.
Выделяют также судорожный плазмолиз, подобный выпуклому, но отличающийся от него тем, что сохраняются цитоплазматические нити, соединяющие сжавшуюся цитоплазму с клеточной стенкой, и колпачковый плазмолиз, характерный для удлиненных клеток.
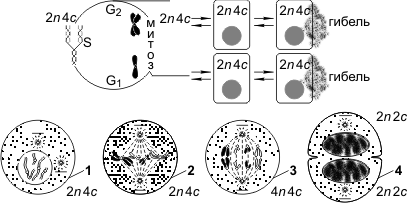
13. Митотический цикл
Митоз — основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала.Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют митотический цикл. Ниже приводится краткая характеристика фаз цикла.
Интерфаза состоит из трех периодов: пресинтетического, или постмитотического, — G1, синтетического — S, постсинтетического, или премитотического, — G2.
Пресинтетический период (2n 2c, где n — число хромосом, с — число молекул ДНК) — рост клетки, активизация процессов биологического синтеза, подготовка к следующему периоду.
Синтетический период (2n 4c) — репликация ДНК.
Постсинтетический период (2n 4c) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей.
Профаза (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом.
Метафаза (2n 4c) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза (4n 4c) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами).
Телофаза (2n 2c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки.
Митотический цикл, митоз
Митотический цикл, митоз: 1 — профаза; 2 — метафаза; 3 — анафаза; 4 — телофаза.
Биологическое значение
Биологическое значение митоза. Образовавшиеся в результате этого способа деления дочерние клетки являются генетически идентичными материнской. Митоз обеспечивает постоянство хромосомного набора в ряду поколений клеток. Лежит в основе таких процессов, как рост, регенерация, бесполое размножение и др.
Мейоз — это особый способ деления эукариотических клеток, в результате которого происходит переход клеток из диплоидного состояния в гаплоидное. Мейоз состоит из двух последовательных делений, которым предшествует однократная репликация ДНК.
Первое мейотическое деление (мейоз 1) называется редукционным, поскольку именно во время этого деления происходит уменьшение числа хромосом вдвое: из одной диплоидной клетки (2n 4c) образуются две гаплоидные (1n 2c).
Интерфаза 1 (в начале — 2n 2c, в конце — 2n 4c) — синтез и накопление веществ и энергии, необходимых для осуществления обоих делений, увеличение размеров клетки и числа органоидов, удвоение центриолей, репликация ДНК, которая завершается в профазе 1.
Профаза 1 (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом, конъюгация гомологичных хромосом и кроссинговер. Конъюгация — процесс сближения и переплетения гомологичных хромосом. Пару конъюгирующих гомологичных хромосом называют бивалентом. Кроссинговер — процесс обмена гомологичными участками между гомологичными хромосомами.
Профаза 1 подразделяется на стадии: лептотена (завершение репликации ДНК), зиготена (конъюгация гомологичных хромосом, образование бивалентов), пахитена (кроссинговер, перекомбинация генов), диплотена (выявление хиазм, 1 блок овогенеза у человека), диакинез (терминализация хиазм).
14. Классификация тканей.
Ткани делят на простые и сложные. Простыми называют ткани, состоящие из клеток более или менее одинаковых по форме и функциям. Сложные ткани состоят из клеток, разных по форме и функциям, но тесно взаимосвязанных в своих жизненных отправлениях. Пример первых — столбчатая хлоренхима, губчатая хлоренхима, колленхима, вторых — ксилема, флоэма.
Ткани делятся на образовательные (меристема) и постоянные.
Образовательными называются специализированные ткани, клетки которых сохраняют длительную способность к делению, обеспечивая рост растения и отдельных его органов. С учетом положения в теле растения их делят на апикальные (или верхушечные, находятся на апексах корня и побега), интеркалярные (или вставочные, свойственны побегу — стеблю и листьям, находятся в междоузлиях и черешках) и боковые (или латеральные, представлены главным образом в осевых органах — в корне и стебле голосеменных и двудольных покрытосемянных).
Постоянными называют ткани, клетки которых утратили способность к делению (полностью или сохраняют её потенциально) и специализируются на выполнении других функций: защитной, запасающей, механической, проводящей и т. д. С учетом происхождения, преобладающей функции и положения в теле растения постоянные ткани, в свою очередь, делят на покровные, проводящие и основные, начало которым при первичном росте дают соответственно протодерма, прокамбий и основная меристема.
Наряду с анатомо-физиологической существует и онтогенетическая классификация тканей, основанная на их происхождении и времени появления в процессе морфогенеза органа. По этой классификации ткани делят на первичные и вторичные.
Первичные меристемы ведут свое начало от первой клетки нового организма — зиготы, которым свойственна способность к делению. Они первыми формируются при заложении нового организма и обеспечивают его первичный рост. Это — верхушечные и вставочные меристемы. Те постоянные ткани, клетки которых дифференцируются из производных клеток первичной меристемы, называют первичными. К ним относят ткани: первичные покровные, первично проводящие и основные.
Вторичными называют меристемы, которые формируются в вегетативных органах позднее первичных и обеспечивают их вторичный рост. Это боковые меристемы — камбий и феллоген (пробковый камбий). Постоянные ткани, начало которым дали производные клетки вторичной меристемы, называют вторичными. К ним относятся вторичную покровную ткань, вторичные проводящие ткани.
Понятие о тканях.
Характерной особенностью высших растений является то, что их клетки объединяются в группы, образуя настоящие ткани. Ткань - это совокупность взаимосвязанных клеток, имеющих общее происхождение, сходные по строению и выполняемым функциям. В ткани клетки связаны структурно и функционально. Наука, изучающая ткани, их происхождения, характерные особенности, функции, называется гистологией (от гр. Сл. "Histos" - ткань).
Растительные ткани можно классифицировать по различным признакам - по происхождению, по плотности и компактностью расположения клеток, по сложности, по выполняемым функциям, по уровню жизнедеятельности, по характеру дифференциации и другим признакам. По сложности выделяют простые ткани, клетки которых однородные по строению и выполняют одну функцию, и комплексные ткани, является полифункциональными и состоят из разнородных элементов. Зависимости от выполняемых функций выделяют следующие типы тканей - образующие, покровные, основные, механические, ведущие, выделительные и другие. По происхождению ткани бывают первичными, происходящих от малодифференцированных клеток, и вторичные, образующиеся в результате разделения уже дифференцированных взрослых клеток. По плотности различают рыхлые (с хорошо выраженными межклетников) и плотные ткани. По характеру дифференциации ткани подразделяют на эмбриональные и постоянные.
Основной причиной возникновения тканей в процессе эволюции растений был их выход на сушу. При этом растения попали в совершенно другой, чем водное, среда обитания, характеризующейся более сложным влиянием внешних условий. В надземных условиях существования растениям необходимы были более надежны внешние покрытия для защиты от различных неблагоприятных и изменчивых условий. Закрепление растений в почвенном субстрате обусловило дифференциацию их тела на подземную часть, выполняющая функцию минерального питания, и на надземную часть, выполняющая функцию воздушного питания.
Это обусловило формирование различных типов покровной и основной тканей. На суше возникли осложнения с поступления различных веществ к органам растений. Поэтому развиваются специальные проводящие элементы. Для поддержания тела в пространстве, предоставление ему определенной формы и противодействия внешним нагрузкам необходима была специальная Укрепляющая ткань, функцию которой начала выполнять механическая ткань.
15. Меристемы различны по расположению в отдельпых органах. калъные, или верхушечные, меристемы расположены в окончаниях (верхушках) стебля, растущих побегов и корня п обеспечивают верхушечный тип роста. Вставочные, или интеркалярные, меристемы, расположенные между двумя закончившими рост тканями, характерны для стебля (рост междоузлий у злака) и для некоторых листьев. Этим меристемам соответствует интеркалярный тип роста. Вааалъ-ные- меристемы расположены у основания органа и характерны для листьев, которые растут основанием. Наконец, существуют меристемы, обеспечиваюш ие рост стебля в толщину, расположенные в стебле между ксилемой и флоэмой (камбий).
В онтогенезе растения растущим и запасающим тканям принадлежит ведущая роль в ориентации потоков ассимилятов нз листьев, В растении возникает несколько аттрагирующих (потребляющих) зон верхушечная меристема стебля, кончики корней, участки интеркалярного роста, плоды и запасающие паренхимные ткани — это главные центры, между которыми распределяется большая часть подвижных ассимилятов. Указанные зоны возникают в определенной последовательности соответственно программе онтогенеза растения. Потребность в ассимилятах у каждого из таких центров в онтогенезе неодинакова.
Растения в отличие от животных в течение всей жизни растут, образуя новые ткани и органы, которые закладываются в эмбриональных зонах — меристемах, где все клетки делятся. Существование меристем поддерживается инициальными клетками инициалями), способными к делению неопределенно длительное время. Апикальные верхушечные) меристемы расположены на концах побегов и корней. Латеральные боковые) меристемы образуют слои клеток вдоль каждого побега и корня (у двудольных растений). К ним относятся первичные латеральные меристемы — прокамбий и перицикл и вторичные — камбий и феллоген. В основании молодых междоузлий и листьев функционируют интеркалярные вставочные) меристемы .При повреждении у растений активируются меристемы, за счет которых регенерируют ткани, органы и целые организмы.
Сначала происходит первичный рост. В результате первичного роста может сформироваться целое растение, и у большинства однодольных и травянистых двудольных это единственный тип роста. В нем участвует апикальная, а иногда и интеркалярная меристема. Строение зрелых первичных корней и стеблей описано в разд.
Физиология и биохимия действия. Действие ГА на растения наиболее типично проявляется в удлинении их стебля. Особенно наглядно это прослеживается у карликовых растений и у длиннодневных растений, находящихся в фазе розетки. У последних гиббереллин способствует образованию цветоноса, а часто и зацветанию. Места действия ГА — апикальные и интеркалярные меристемы, где под влиянием этого фитогормона активируется деление клеток. Ускоряется также растяжение клеток, однако непосредственно на этот процесс ГА действует слабо. Гиббереллины не стимулируют рост корня, а в повышенных концентрациях даже ухудшают его состояние. Как правило, добавки ГА не требуется и при выращивании культуры тканей. Обработка гиббереллином выводит семена и клубни некоторых растений из состояния покоя.
Интеркалярная меристема находится не непосредственно над морфологически нижним узлом и отделена от него участком (5—7 мм) немеристематических тканей, который состоит из двух зон растяжения и остаточной дифференциации. Кпизу от зоны интерка-лярной меристемы растяжение клеток сопровождается значительным повышением активности оксидоредуктаз и накоплением крахмала, а кверху от нее — незначительным повышением активности этих ферментов и отсутствием изменений в содержании крахмала.
Кроме деления меристем на детерминированные и недетерминированные существуют и другие классификации. Например, апикальные меристемы стеблей и корней отличаются от латеральных меристем, включающих камбий и феллоген (пробковый камбий). У некоторых растений имеются интеркалярные меристемы, расположенные между участками дифференцированных тканей. Прекрасным примером наличия такого типа.
Представителем этого порядка может служить род эктокарпус E to arpus). Многочисленные виды его распространены во всех морях, особенно холодных, растут на различных подводных объектах и на других более крупных водорослях. Таллом (как спорофита, так и гаметофита) имеет вид желтовато-бурых кустиков, достигающих в длину нескольких сантиметров. Он состоит из стелющихся по субстрату нитей, от которых отходят вертикальные нити из одного ряда клеток, обильно ветвящиеся. Вегви вертикальных нитей часто оканчиваются многоклеточными бесцветными волосками. Стелющиеся по субстрату нити обнаруживают верхушечный рост. Рост вертикальных нитей у разных видов неодинаков, У большинства видов рост диффузный и лищь у некоторых видов дифференцируется ясно выраженная интеркалярная вставочная меристема у основания волосков еще реже наблюдается верхушечный рост.
В ходе эвокации под действием флорального стимула в апикальной меристеме стебля индуцируется экспрессия генов, ответственных за морфогенетическую программу цветообразова-ния. Увеличивается число клеточных делений, изменяется форма апекса. Первыми закладываются чашелистики, появляются примордии лепестков, затем последовательно закладываются тычинки и плодолистики. Прежде всего, как и при формировании примордиев листа, физиологически и морфологически выделяются группы инициальных клеток, особенности деления которых определяют дальнейший ход морфогенеза каждой части цветка. Например, тычиночная нить образуется за счет апикального, а затем интеркалярного деления и роста клеток примордия. В дистальной части инициальные клетки дают начало тканям пыльника.
Эумеристемы, дающие начало всем тканям растения, кроме проводящих, состоят из изодиаметрических, в очертании многоугольных клеток. Каждая клетка эумеристемы имеет крупное ядро, занимающее около половины объёма клетки, а ядерная оболочка имеет много пор. В цитоплазме много диффузно расположенных рибосом. Имеются пропластиды, митохондрии и диктиосомы. Вакуоли немногочисленные и мелкие. Соседние клетки соединены более или менее диффузно расположенными плазмодесмами.
Клетки меристем, образующих проводящие ткани (камбий и прокамбий), прозенхимные, снабжены крупными вакуолями.
Клеткам меристем свойственно неравное (дифференцированное) деление. Клетка делится митозом на 2: одна остаётся клеткой меристемы, а другая, поделившись один или несколько раз, приступает к дифференцировке.
Митотическая активность клеток меристемы различна. Наиболее активные в этом отношении клетки — инициальные клетки, дающие начало производным клеткам. Производные могут отличаться от инициальных формой, размерами, количеством вакуолей.
Увеличение общего объёма меристемы может быть следствием только иницииальных клеток (так, например, нарастают талломы некоторых бурых водорослей), либо чередованием делений инициальной клетки и производных, причём большее количество делений приходится на производные клетки. Такая периодичность свойственна конусам нарастания большинства высших споровых. У семенных же инициальная клетка и производные морфологически неразличимы.
Классификация меристем и их производных
Морфологическая классификация
Морфологическая классификация основана на ориентации перегородок в делящихся клетках. Выделяется три морфологических типа меристем:
Пластинчатые меристемы однослойные, клетки, таблитчатые на поперечном срезе, делятся антиклинально (перпендикулярно плоскости органа). Участвует в образовании эпидермы.
Колончатые, или стержневые меристемы состоят из кубических или призматических клеток, располагающихся продольными рядами. У некоторых растений такие меристемы образуют сердцевину стебля.
Массивные меристемы состоят из многоугольных в очертании клеток, делящихся в разных направлениях и обеспечивающих более или менее равномерное увеличение объёма ткани. Из массивных меристем развивается, например, спорогенная ткань спорангия.
Онтогенетическая классификация
Все меристемы, функционирующие на протяжении всей жизни растения, можно подразделить на общие и специальные. К общим меристемам относится меристема зародыша, из которой развиваются апикальные (верхушечные) меристемы, а они, в свою очередь, дают начало третьей группе общих меристем — интеркалярным.
От апикальных меристем берут начало:
протодерма, образующая эпидерму;
основная меристема, дающая начало системе тканей основной паренхимы;
прокамбий.
Клетки этих трёх тканей более вакуолизированы, чем апикальные меристемы, и обладают ограниченной способностью к делению, поэтому часто их называют полумеристемами.
Часть клеток прокамбия даёт начало первичным проводящим тканям — первичным ксилеме и флоэме; из других его клеток развивается камбий, который, в свою очередь, образует вторичные проводящие ткани — вторичные ксилему и флоэму. Вторичные проводящие ткани образует и т. н. добавочный камбий, развивающийся не из прокамбия, а из постоянных тканей, живые клетки которых при этом дедифференцируются, то есть возвращаются из специализированного состояния к состоянию, близкому к эмбриональному, и вновь приобретают способность к делению.
Таким же образом в растении возникают и раневые меристемы, восстанавливающие повреждённые участки растения. На возможности новообразования меристем основано вегетативное размножение растений.
Кроме добавочного камбия, постоянные ткани также участвуют в образовании феллогена (пробкового камбия). Делясь периклинально (то есть параллельно поверхности органа), клетки феллогена отделяют наружу будущие клетки пробки (феллемы), а внутрь — феллодерму.
Топографическая классификация
В соответствии с расположением, помимо апикальных меристем, локализованных на кончиках осевых органов — стебля и корня, выделяют меристемы:
латеральные (боковые) меристемы расположены внутри осевых органов и осуществляют их утолщение. К латеральным меристемам относят камбий, добавочный камбий и феллоген.
краевая (маргинальная) меристема даёт начало листовой пластинке.
интеркалярные меристемы осуществляют вставочный рост. За счёт их деятельности удлиняются междоузлия на ранних этапах развития побега, развиваются черешки листьев. Принципиально они отличаются от прочих меристем тем, что, помимо недифференцированных клеток, в них существуют и клетки на разных стадиях дифференциации. Поэтому корректнее здесь говорить не об интеркалярных меристемах, а о зонах интеркалярного (вставочного) роста.
Все эти меристемы, кроме феллогена, являются производными апикальных меристем.
Генетическая классификация
Согласно генетической классификации выделяют первичные и вторичные меристемы. Первичные меристемы связаны с меристемами зародыша и апикальными меристемами, а вторичные развиваются позднее, когда начинается утолщение осевых органов и заканчивается верхушечный рост. Однако такое разделение условно, так как, например, камбий закладывается до окончания верхушечного роста.
Основная статья: Апикальная меристема
Развивающийся зародыш любого растения состоит из эумеристемы. По мере формирования зачатков вегетативных органов у зародыша эумеристема остаётся только в двух местах: в верхушке зачаточного побега (будущий конус нарастания). Эти меристемы и называют апикальными меристемами (от лат. apex — вершина). Они осуществляют рост побегов и корней в длину и дают начало большинству других меристем.
Когда растение вступает в фазу цветения, верхушечная меристема стебля преобразуется в меристему соцветия. От неё берёт начало флоральная меристема, образующая структуры цветка: лепестки, чашелистики, плодолистики и тычинки.
16. Покровные ткани
Покро́вные тка́ни — наружные ткани растения.
Покровные ткани предохраняют органы растения от высыхания, от температурных воздействий, механических повреждений, гиф грибов, болезнетворных бактерий и вирусов и других неблагоприятных воздействий окружающей среды. Осуществляют всасывание и выделение воды и других веществ.
Через покровные ткани стебля осуществляется газообмен. В эпидерме он происходит через устьица. После образования перидермы эпидерма отмирает и слущивается, и газообмен идёт через чечевички.
Часто эпидерма растений несёт различные образования: эмергенцы, кроющие и железистые волоски (трихомы), составляющие опушение растения.
Эпидерма, образование и функции
Эпиде́рма (эпиде́рмис, ко́жица) — внешняя первичная покровная ткань растений, обычно однослойная[1], покрывающая молодые стебли и остальные наземные органы (листья, лепестки, плоды и др.). Представляет собой наружный слой клеток, образующийся из протодермы конуса нарастания.
Эпидерма — многофункциональная ткань, но её основными функциями являются защита внутренних тканей растения, а также осуществление газообмена и транспирации. Эти процессы осуществляются через устьица.
Продолжительность существования эпидермы у различных растений и их органов неодинакова. На листьях и стеблях травянистых растений эпидерма сохраняется до конца их жизни, а в стеблях древесных растений, характеризующихся вторичным утолщением, эпидерма заменяется вторичной покровной тканью — пробкой (феллемой). Эпидерма при этом отмирает и слущивается, и вместо устьиц газообмен осуществляют чечевички.
Важнейшие функции — защита растений от неблагоприятных внешних факторов и регуляция газо- и парообмена. Кроме того, ткань кожицы может выделять наружу различные вещества (соли, воду, эфирные масла), принимать участие в фотосинтезе, поглощении воды и питательных веществ, синтезе различных соединений, в движении листьев, воспринимать раздражение и т.д. Полифункциональность эпидермиса обусловливает его строение. Эпидермис — сложная ткань, так как состоит из морфологически разнородных элементов.
Регуляция транспирации в большей степени обусловливается наличием жирового вещества кутина, часто в комплексе с воском. Эти вещества инкрустируют наружную стенку или образуют самостоятельный слой — кутикулу — на поверхности эпидермиса.
Кутикула может достигать значительной толщины, особенно у растений засушливых местообитаний. Комплекс кутикулы и кутинизированной оболочки представляет покров, не только защищающий растение от иссушения, она предохраняет растение от заражения всевозможными грибами-паразитами, бактериями, вирусами, которые в изобилии находятся на его поверхности.
Основные клетки
В наружной стенке основных клеток могут быть поры. Как правило, основные клетки живые, они содержат пластиды, обычно хлоропласты. Правда, хлоропласты эпидермы имеют слабо развитую систему внутренних мембран и, в отличие от хлоропластов хлоренхимы, фотосинтетически менее активны. Иногда основные клетки могут содержат лейкопласты (подорожник большой). Они также имеют слабо развитую систему внутренних мембран и обычно не содержат крахмала. У многих растений эпидермальные клетки лепестков и плодов содержат хромопласты, обусловливающие их окраску.
Для основных клеток эпидермы характерна крупная центральная вакуоль, накапливающая различные вещества: оксалат кальция, таннины, алкалоиды, пигменты (обычно либо антоциан, либо антохлор). Короткие клетки эпидермы злаков содержат кремниевые тельца.
Устьица в ботанике это по́ра, находящаяся на нижнем или верхнем слое эпидермиса листа растения, через которую происходит испарение воды и газообмен с окружающей средой.
Размеры устьица (длина) колеблются в пределах 0,01–0,06 мм (крупнее бывают устьица полиплоидных растений и у листьев, растущих в тени. Самые крупные устьица были обнаружены у вымершего растения Zosterophyllum, 0,12 мм (120 мкм). Пора состоит из пары специализированных клеток, называемых замыкающими (cellulae claudentes), которые регулируют степень открытости поры, между ними располагается устьичная щель (porus stomatalis). Стенки замыкающих клеток утолщены неравномерно: направленные к щели (брюшные) толще стенок, направленных от щели (спинных). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель. Под щелью расположена подустьичная (воздушная) полость, окружённая клетками мякоти листа, через которую непосредственно и происходит газообмен. Воздух, содержащий диоксид углерода (углекислый газ) и кислород, проникает внутрь ткани листа через эти поры, и далее используется в процессе фотосинтеза и дыхании. Избыточный кислород, произведённый в процессе фотосинтеза внутренними клетками листа, выходит обратно в окружающую среду через эти же поры. Также, в процессе испарения через поры выделяются пары воды. Клетки эпидермиса, примыкающие к замыкающим, получили название сопровождающих (побочных, соседних, околоустьичных). Они участвуют в движении замыкающих клеток. Замыкающие и сопровождающие клетки образуют устьичный комплекс (устьичный аппарат). Наличие или отсутствие устьиц (видимые части устьиц называют устьичными линиями) часто используют при классификации растений.
Так как углекислый газ является одним из ключевых реагентов в процессе фотосинтеза, у большинства растений устьица в дневное время открыты. Проблема состоит в том, что при входе воздух смешивается с парами воды, испаряющимися из листа, и поэтому растение не может получить углекислый газ, одновременно не потеряв некоторое количество воды. У многих растений существует защита от испарения воды в виде закупоривающих устьица восковых отложений.
Трихо́мы, или волоски́ (от греч. τρίχωμα — волос) — клетки эпидермы или выросты, образующие опушение на поверхностных органах растений, могут присутствовать на всех наземных органах растения[1]. Трихомы следует отличать от эмергенцев — межклеточных выростов, в образованиях которых участвует не только эпидерма, но и субэпидермальные ткани.
Всё многообразие трихом делят на два функциональных типа: кроющие и железистые. Первые образуются из покровных тканей и служат для защиты растения от неблагоприятного воздействия внешней среды, вторые — принадлежат к выделительным тканям наружной секреции и участвуют процессах накопления и выделения веществ различного функционального назначения.
Трихомы бывают одноклеточными и многоклеточными, мертвыми и живыми. Мертвые заполнены воздухом и придают растению белый цвет. Форма трихом может быть разнообразной (головчатые, звездчатые, крючковатые и др.). Часто трихомы минерализованы — пропитаны кремнеземом и кальцием.
Размеры трихом варьируются в значительных пределах. Чаще отдельный волосок, чешуйка или желёзка хорошо различимы в сильную лупу или микроскоп. Наиболее длинные трихомы (до 5—6 см) покрывают семена хлопчатник .
Трихомы выполняют разнообразные функции, которые до конца ещё не исследованы. Принято считать, что они защищают физиологически ткань листа — хлоренхиму от перегрева, механически всё растение от поврежедения насекомыми и животными, способствуют уменьшению испарения влаги, выведению солей из тканей листа и осуществляют химическую защиту растений.
Удивительно, базальные клетки трихом обоих типов соединены с субэпидермальными клетками значительно большим числом плазмодесм, чем с соседними эпидермальными клетками. Таким образом трихомы, генетически и структурно относясь к эпидерме, физиологически теснее связаны с субэпидермальными тканями. Функции редких рассеянных трихом неизвестны
17. Покровные комплексы - перидерма и корка. На смену перидерме в стебле закладывается вторичная покровная ткань – пробка. Её образование связано с заложением вторичной меристемы пробкового камбия. Клетки пробкового камбия делятся и дифференцируются к наружи – в пробку, а внутрь – в феллодерму. Комплекс, состоящий из феллогена, феллемы и феллодермы называется перидермой. Для газообмена и транспирации в пробке есть особые образования – чечевички.
18. Эпиблема (ризодерма) - первичная однослойная поверхностная ткань корня. Основная её функция – всасывание, избирательное поглощение из почвы воды с растворёнными в ней элементами минерального питания. Через эпиблему выделяются некоторые вещества, например кислоты, действующие на субстрат и преобразующие его. Это тонкостенные клетки, лишённые кутикулы, с вязкой цитоплазмой, с большим числом митохондрий (активное поглощение веществ происходит с затратой энергии). Поглощающая поверхность эпиблемы увеличивается в 10 раз и более за счёт образования корневых волосков (выростов клетки длиной 1…2(3) мм).
19. Основные ткани занимают наибольший объём в растению. Они состоят из живых паренхимных клеток, тонкостенных, с простыми порами. В зависимости от выполняемой функции различают основную (типичную), ассимиляционную, запасающую и воздухоносную основные ткани. Основная паренхима не имеет специфических, строго определённых функций, располагается внутри тела растения достаточно крупными массивами. Типичная основная паренхима заполняет сердцевину стебля, внутренние слои коры стебля и корня. Её клетки образуют вертикальные и горизонтальные тяжи (лучи), по которым в радиальном направлении перемещаются вещества. Из неё могут возникать вторичные меристемы. Ассимиляционная паренхима (хлоренхима) характеризуется наличием в её клетках хлорофилловых зёрен. Функция – процесс фотосинтеза. Хлоренхима расположена под эпидермисом листьев и зелёных стеблей, различают столбчатую и губчатую ассимиляционную паренхиму. Столбчатая паренхима располагается под верхним эпидермисом листа, участвует в процессе фотосинтеза. Губчатая, наряду с фотосинтезом, осуществляет газообмен и транспирацию благодаря крупным межклетникам. Запасающая паренхима. Вещества, синтезированные растением, могут откладываться в виде запасов. К накоплению запасных веществ способны все живые клетки, но если запасающая функция выступает на 1 место, то говорят о запасающих тканях. Запасающие ткани широко распространены в самых различных органах. У однолетних растений запасные вещества – в семенах, плодах. У многолетних запасы в специализированных органах (клубнях, корневищах, луковицах). Запасные вещества накапливаются в твёрдом и растворённом виде. У некоторых растений запасным веществом служат гемицеллюлозы, входящие в состав клеточных оболочек (клеточная оболочка финиковой пальмы). При прорастании семян гемицеллюлозы под действием ферментов превращаются в сахара, которые используются проростком. В растворённом виде накапливаются сахара в корнях свёклы, моркови. Воздухоносная паренхима (аэренхима) её отличительной чертой являются крупные межклетники. Особенно сильного развития достигает у водных растений и растений, растущих на болотной почве. Её функция – облегчить дыхание погружённых в воду частей растений.
20. Механические ткани. Основным признаком механических тканей является утолщение стенок клетки. Механические ткани подразделяются на: -склеренхиму, -колленхиму, -склереиду. Колленхима – первичная механическая ткань с неравномерно утолщёнными не одревесневающими живыми клетками, располагается под эпидермой, встречается чаще всего у двудольных растений. Колленхима возникает в молодых побегах, когда продолжается их рост в длину. Благодаря своему строению, она сама способна растягиваться по мере растягивания окружающих клеток. Особенностью колленхимы является то, что она выполняет свою функцию только в состоянии тургора. Колленхима бывает уголковая, пластинчатая, рыхлая (с межклетниками). Склеренхима – резко прозенхимные клетки, их длина достигает 50см. Клеточные стенки одревесневающие, утолщены. Выполняет функцию уже после отмирания протопласта. По происхождению склеренхима бывает как первичная, так и вторичная у однодольных и двудольных растений. Может быть представлена лубяными волокнами с целлюлозными клеточными стенками. Волокна стеблей двудольных используют для изготовления различных тканей. Склереиды – форма разнообразная, по происхождению первичны, клеточная стенка одревесневающая, равномерно утолщена, пронизана порами в поперечном направлении. Встречаются в плодах, семенах, листьях и стеблях. Механические ткани в стебле занимают периферическое положение, а в корнях – центральное.
21. Проводящие ткани и комплексы, их функции. Строение трахеальных элементов — трахеид, сосудов, ситовидных элементов — ситовидных клеток,ситовидных трубок. Образование тил и каллезы.
Проводящие ткани образуют в теле растения непрерывную разветвленную систему,соединяющую все органы. Провод.ткани служат для транспорта веществ в растении. Могут быть как первичны, так и вторичны. Первич. образуются из прокамбия,вторич.-из камбия. Проводящ.ткани подразделяются на ситовидные трубки,сосуды(трахеи), трахеиды.Ситовид.трубки- вертикальный ряд живых клеток,у которых первичные стенки перфорированы. Рядом с трубкой располежены одна или несколько клеток-спутниц. По ситовид.трубкам транспортируются растворы органических веществ. На зиму ситовидные пластинки у многих растений закупориваются каллезой, а весной это вещество растворяется, и трубки вновь начинают функционировать или же весной ситовидные трубки отмирают и заменяются новыми, возникающими вследствие деятельности камбия.Ситовид.клетки— вытянутые клетки,ситовидные поля которой рассеяны по боковым стенкам. В зрелых ситовид.клетках сохраняется ядро. Сосуды — это трубки, у которых утолщаются и одревесневают боковые стенки, отмирает протопласт,а в поперечных стенках образуется один или несколько перфораций. Сосуды бывают кольчатые,спиральные, сетчато-пористые и др. Сосуды функционируют ограниченное время. Прекращение их деятельности связано с закупоркой тилами.Тилы- это выросты соседних клеток,проникающие в полость через поры. Здесь они разрастаются,лигнифицируются,накапливаются смолы,камеди,танины и закупоривают сосуды.Деятельность сосудов прекращается,но они продолжают выполнять механические функции. Трахеиды— мертвые прозенхимные клетки, в их стенках имеются окомлённые поры. По сосудам и трахеидам транспортируются вода и растворенные в ней минеральные вещества. Фукциипроводящих тканей:1) передвижение воды и минеральных,органических веществ, поглощенных корнями из почвы 2)передвижение продуктов фотосинтеза в места использования и запасания: в корни, стебли, плоды и семена 3)повышают устойчивость органов растений к деформирующим нагрузкам 4)образуют непрерывную разветвленную систему, связывающую органы растений в единое целое.
22,Проводящие комплексы-ксилема, флоэма, их гистологический состав. Проводящие пучки и их типы.
Проводящие ткани располагаются в органах растений в виде продольных тяжей, образуя проводящие пучки. Проводящие пучки-комплекс проводящих тканей. В нем различают 2 зоны:ксилему(древесину), служащюю для транспорта минеральных вещест; ифлоэму(луб),для транспорта органических веществ. Основная часть ксилемы-сосуды и трахеиды,им сопутствует древесные паренхима и волокна;Основная часть флоэмы-ситовидные трубки,им сопутсвуют лубяные паренхима и волокна.Ксилема и флоэма образуются в результате работы специальных меристем-прокамбия и камбия. Ксилема, флоэма, возникшие из прокамбия-первичные,а из камбия — вторичные. Если между флоэмой и ксилемой есть камбий,то пучок открытый,если нет-закрытый. Проводящие пучки бывают коллатеральные(флоэма лежит к наруже от ксилемы),биколлатеральные(флоэма расположенна с обеих сторон ксилемы;открытые),концентрические(пучки,в которых или ксилема окружает флоэму,или флоэма ксилему,всегда закрытые), радиальные(в них ксилема и флоэма расположены по радиусам)
23.Выделительные ткани и их функции у растений. Жизнь растений представляет собой совокупность биохимических реакций, скорость и интенсивность которых в значительной мере модифицируется условиями среды произрастания. В этих реакциях образуется большое разнообразие побочных продуктов, не используемых растением для построения тела или для регулирования обмена веществами, энергией и информацией с окружающей средой. Такие продукты могут удаляться из растения разными способами: при отмирании и отделении ветвей и участков корневищ, при опадании листьев, в результате деятельности специализированных структур внешней и внутренней секреции. В совокупности эти приспособления образуют выделительную систему растений. В отличие от животных специальной выделительной системы у растений нет, однако экскреторные вещества из организма удаляются или скапливаются в отдельных вместилищах. Выделительная система растений многофункциональна. В её структурах осуществляются: синтез, накопление, проведение и выделение продуктов метаболизма. Выделительная ткань бывает внутренней и наружной секреции,в зависимости от того выделяются вещества наружу или остаются внутри растения. Структуры внешней секрециирасполагаются на поверхности органов растений и выделяют свои продукты, или секреты, во внешнюю среду. К ним относятся железистые волоски, желёзки, нектарники, гидатоды, осмофоры, переваривающие желёзки. К структурам внутренней секрецииотносятся млечники, или млечные сосуды, секреторные вместилища,секреторные клетки и идиобласты. Их основными функциями является образование, транспорт и накопление смол, эфирных масел, дубильных веществ, млечного сока, кристаллов солей.
24.Ткани внешней секреции — железистые волоски,секреторные желёзки,нектарники, осмосфоры, гидатоды.
Структуры внешней секреции располагаются на поверхности органов растений и выделяют секреты во внешнюю среду.Железистые волоски являются трихомами, т.е. выростами эпидермиса. Они характерны для растений семейств Астровые, Паслёновые, Яснотковые и др. Железистые волоски бывают простыми и сложными. У простых волосков одна (томат) или несколько (табак) вытянутых клеток образуют длинную ножку, клетки которой имеютхлоропласты. На ножке располагается одно- или многоклеточная головка с густой цитоплазмой, но не содержащая хлоропластов. У сложных волосков клетки ножки и головки не содержат хлоропластов и обеспечивают выделение веществ. По мере увеличения объёма секрета кутикула растягивается и лопается, что обеспечивает выход выделяемых веществ наружу. После этого образуется новый слой кутикулы и начинается формирование новой капли секрета. Желёзки имеют короткую многоклеточную ножку и многоклеточную округлую или уплощенную щитовидную головку и весьма часто располагаются на почечных чешуях древесных растений. Они известны также у астровых, крыжовниковых и в других семействах. Нектарники- образуются в цветке;они представлены отдельными клетками или находятся в ямках,шпорцах. Нектар-водный раствор сахаров с небольшой примесью белков,спиртов и ароматических средств. Важнейшими нектароносными растениями являются плодовые семечковые и косточковые породы, ягодные кустарники, липа, акация, вереск, клевер и др. Осмосферы-специализированные клетки эпидермы или особые желёзки,где вырабатываются ароматические вещества. Гидатоды-водяные устьица, которые выделяют воду. Приэтом вместе с водой могут выделяться соли, сахара и другие органические вещества. Такое явление называется гуттацией;встречаются у представителей семейств Капустные, Розовые, Первоцветные, Мятликовые, Толстянковые