

8.Ядро клетки в интерфазе. Хроматин: уровни организации (упаковки) наследственного материала (эухроматин, гетерохроматин).
В организме человека содержатся только эукариотические (ядерные) типы клеток. Безъядерные структуры (эритроциты, тромбоциты, роговые чешуйки) являются вторичными (постклеточными) образованиями, так как они образуются из ядерных клеток в результате их специфической дифференцировки. В подавляющем большинстве клеток содержится одно ядро, но встречаются двуядерные и даже многоядерные клетки. Форма ядра в большинстве клеток круглая (сферическая) или овальная. В некоторых клетках ядра имеют вытянутую или палочковидную форму. В зернистых лейкоцитах ядро подразделяется на сегменты (сегментоядерные лейкоциты). Локализуется ядро обычно в центре клетки, но в клетках эпителиальных тканей ядра нередко сдвинуты к базальному полюсу.
Структурные элементы ядра бывают четко выражены только в определенный период клеточного цикла в интерфазе. В период деления клетки (в период митоза или мейоза) одни структурные элементы исчезают, другие существенно преобразуются.
Классификация структурных элементов интерфазного ядра:
• хроматин;
• ядрышко;
• кариоплазма;
• кариолемма.
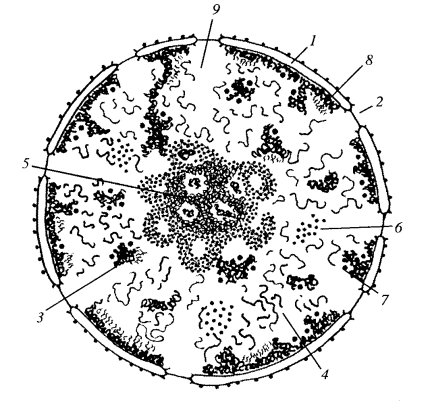
Хроматин представляет собой вещество, хорошо воспринимающее краситель (хромос), откуда и произошло его название. Хроматин состоит из хроматиновых фибрилл, толщиной 20-25 нм, которые могут располагаться в ядре рыхло или компактно. На этом основании различают два вида хроматина: • эухроматин - рыхлый или деконденсированный хроматин, слабо окрашивается основными красителями;
• гетерохроматин - компактный или конденсированный хроматин, хорошо окрашивается этими же красителями.
При подготовке клетки к делению в ядре происходит спирализация хроматиновых фибрилл и превращение хроматина в хромосомы. После деления в ядрах дочерних клеток происходит деспирализация хроматиновых фибрилл и хромосомы снова преобразуются в хроматин. Следовательно, хроматин и хромосомы представляют собой различные фазы одного и того же вещества.
По химическому строению хроматин состоит из:
• дезоксирибонуклеиновой кислоты (ДНК) 40 %;
• белков около 60 %;
• рибонуклеиновой кислоты (РНК) 1 %.
Ядерные белки представлены формами:
• щелочными или гистоновыми белками 80-85 %;
• кислыми белками 15-20 %.
Гистоновые белки связаны с ДНК и образуют полимерные цепи дезоксирибонуклеопротеида (ДНП), которые и представляют собой хроматиновые фибриллы, отчетливо видимые при электронной микроскопии. На определенных участках хроматиновых фибрилл осуществляется транскрипция с ДНК различных РНК, с помощью которых осуществляется затем синтез белковых молекул. Процессы транскрипции в ядре осуществляются только на свободных хромосомных фибриллах, то есть в эухроматине. В конденсированном хроматине эти процессы не осуществляются и потому гетерохроматин является неактивным хроматином. Соотношение эухроматина и гетерохроматина в ядре является показателем активности синтетических процессов в данной клетке. На хроматиновых фибриллах в S-периоде интерфазы осуществляется также процессы редупликации ДНК. Эти процессы происходят как в эухроматине, так и в гетерохроматине, но в гетерохроматине они протекают значительно позже. Ядрышко - сферическое образование (1-5 мкм в диаметре) хорошо воспринимающее основные красители и располагающееся среди хроматина. В одном ядре может содержаться от 1 до 4-х и даже более ядрышек. В молодых и часто делящихся клетках размер ядрышек и их количество увеличены. Ядрышко не является самостоятельной структурой. Оно формируется только в интерфазе в определенных участках некоторых хромосом - ядрышковых организаторах, в которых содержатся гены, кодирующие молекулу рибосомальной РНК. В области ядрышкового анализатора осуществляется транскрипция с ДНК рибосомальной РНК. В ядрышке происходит соединение рибосомальной РНК с белком и образование субъединиц рибосом.
Микроскопически в ядрышке различают:
• фибриллярный компонент - локализуется в центральной части ядрышка и представляет собой нити рибонуклеопротеида (РНП);
• гранулярный компонент - локализуется в периферической части ядрышка и представляет скопление субъединиц рибосом.
В профазе митоза, когда происходит спирализация хроматиновых фибрилл и образование хромосом, процессы транскрипции РНК и синтеза субъединиц рибосом прекращаются и ядрышко исчезает. По окончании митоза в ядрах вновь образованных клеток происходит деконденсация хромосом и появляется ядрышко.
Кариоплазма (нуклеоплазма) или ядерный сок состоит из воды, белков и белковых комплексов (нуклеопротеидов, гликопротеидов), аминокислот, нуклеотидов, сахаров. Под световым микроскопом кариоплазма бесструктурна, но при электронной микроскопии в ней определяются гранулы (15 нм), состоящие из рибонуклеопротеидов. Белки кариоплазмы являются в основном белками-ферментами, в том числе ферментами гликолиза, осуществляющих расщепление углеводов и образование АТФ. Негистоновые (кислые) белки образуют в ядре структурную сеть (ядерный белковый матрикс), которая вместе с ядерной оболочкой принимает участие в создание внутреннего порядка, прежде всего в определенной локализации хроматина. При участии кариоплазмы осуществляется обмен веществ в ядре, взаимодействие ядра и цитоплазмы.
Кариолемма (нуклеолемма) - ядерная оболочка отделяет содержимое ядра от цитоплазмы (барьерная функция), в то же время обеспечивает регулируемый обмен веществ между ядром и цитоплазмой. Ядерная оболочка принимает участие в фиксации хроматина.
Кариолемма состоит из двух билипидных мембран - внешней и внутренней ядерной мембраны, разделенных перинуклеарным пространством, шириной от 25 до 100 нм. В кариолемме имеются поры, диаметром 80-90 нм. В области пор внешняя и внутренняя ядерные мембраны переходят друг в друга, а перинуклеарное пространство оказывается замкнутым. Просвет поры закрыт особым структурным образованием - комплексом поры, который состоит из фибриллярного и гранулярного компонента. Гранулярный компонент представлен белковыми гранулами диаметром 25 нм, располагающимися по краю поры в три ряда. От каждой гранулы отходят фибриллы и соединяются в центральной грануле, располагающейся в центре поры. Комплекс поры играет роль диафрагмы, регулирующей ее проницаемость. Размеры пор стабильны для данного типа клеток, но число пор может изменяться в процессе дифференцировки клетки. В ядрах сперматозоидов ядерные поры отсутствуют. На наружной ядерной мембране могут локализоваться прикрепленные рибосомы. Кроме того, наружная ядерная мембрана может продолжаться в канальцы эндоплазматической сети.
9. Хромосомный и геномный уровни организации наследственного материала во время митотического деления клетки
I. Геномный уровень
Каждый биологический вид характеризуется определенным числом и строением хромосом, совокупность которых составляют хромосомный набор, или кариотип (полный парный набор хромосом, диплоидный набор). Данный набор хромосом содержится в соматических клетках, и поэтому для изучения кариотипа используются только соматические клетки. Все соматические клетки, независимо от их происхождения и строения (за исключением дифференцированных безъядерных клеток или полиплоидных клеток), имеют не только одинаковое числохромосом, но и идентичный набор генов.
Этому способствует то, что, во-первых в S-период интерфазы происходит репликация ДНК клетки, во-вторых, клетки делятся митозом, который является механизмом точного распределения наследственной информации между дочерними клетками.
Характерной особенностью кариотипа является наличие в нем пар гомологичных хромосом, в каждой паре одна хромосома имеют отцовское, другая - материнское происхождение. Гомологичные хромосомы характеризуются одинаковыми размерами и формой, а также специфичностью строения при дифференциальном окрашивании.
В диплоидном наборе различают аутосомы (для человека - хромосомы 1-22 пар) и половые хромосомы. В клетках мужских и женских организмов аутосомы имеют одинаковое морфологическое строение, но при этом следует помнить, что генотипы разных особей различны. Половые хромосомы имеют различное морфологическое строение и содержат негомологичные участки, характерные только для определенной хромосомы. Комбинации половых хромосом определяют генетический пол организма.
Существуют ряд особенностей наследования признаков в зависимости от расположения генов в хромосомах. В связи с этим различают следующие типы наследования признаков:
- аутосомное (доминантное или рецессивное)
- сцепленное с Х-хромосомой (доминантное или рецессивное)
- сцепленное с У-хромосомой (голандрическое)
В отличие от соматических половые клетки содержат гаплоидный набор хромосом, в котором содержится только одна из каждой пары хромосом, и в генетическом плане половые клетки значительно отличаются от соматических:
1. При сперматогенезе образуются два типа сперматозоидов
2. Половые клетки одного организма содержат различный геном - совокупность генов в гаплоидном наборе. В основе этого лежат два механизма: - в результате кроссинговера возникают новые комбинации неаллельных генов в хромосоме
- независимое расхождение пар хромосом (анафаза I мейоза) приводит к возникновению различных комбинаций негомологичных хромосом в гаметах.
Нарушения геномного уровня организации наследственного материала, т.е. изменения числа хромосом в диплоидном или гаплоидном наборе, называются геномные соматические или генеративные мутации. Механизмами возникновения геномных мутациях являются:
1. Соматические мутации возникают в результате нарушения расхождения хроматид в анафазу митоза (гетероплоидии) либо нарушений кариокинеза (возникают полиплоидные клетки) или цитокинеза (возникают многоядерные клетки).
2. Генеративные мутации возникают в результате нарушения расхождения хромосом (анафаза I мейоза) или хроматид (анафаза II мейоза) при образовании половых клеток. При этих нарушениях сформировавшиеся гаметы содержат измененный гаплоидный набор хромосом.
II. Хромосомный уровень
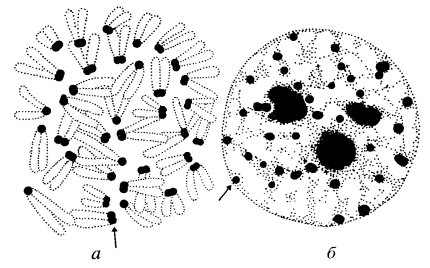
Рассматривает морфологическое строение и структурную организацию отдельных хромосом либо хроматиновых нитей. Такое разделение связано с тем, в какую стадию жизненного цикла клетки изучается хромосомный уровень: хромосомы определяются в клетке во время митоза, а хроматин - во время интерфазы. Структурная организация метафазной хромосомы. Препараты хромосом можно приготовить из любых тканей, содержащих делящиеся клетки. Клетки культивируют в питательной среде, затем останавливают митозы на стадии метафазы и окрашивают хромосомы специальными красителями. На ранних стадиях изучения хромосом использовали простые способы окрашивания (краситель Гимза или ацетоорсеин), при этом хромосомы окрашивались целиком и равномерно. Такой способ окрашивания позволил выявить морфологические особенности строения хромосом: размеры и форму хромосом.
Согласно Денверской классификации (1960) хромосомы располагаются и нумеруются в зависимости от их длины и расположения центромеры. Предложено нумеровать пары хромосом от 1 до 23. С 1-й по 22 пары - аутосомы, 23 пара - Х-хромосома и У-хромосома. По указанным признакам хромосомы разбиты на 8 групп (A - G). Однако существенным недостатком простого способа окрашивания является невозможность идентификации отдельных хромосом внутри группы.
В 70-х годах ХХ века ученые-генетики разработали новые методы окрашивания хромосом - методы дифференциального окрашивания. В настоящее время существуют несколько модификаций метода дифференциального окрашивания хромосом, которые отличаются использованием определенного флюоресцентного красителя или дополнительными процедурами перед окраской хромосом (тепловая обработка, использование солевых растворов, ферментов). Во всех методах наблюдается неравномерность окрашивания хромосом, при этом каждую хромосому можно надежно идентифицировать. На Парижской конференции по стандартизации и номенклатуре хромосом (1971) было предложено дополнить классификацию хромосом особенностями их сегментарной окраски. Каждая хромосома рассматривается как непрерывная совокупность сегментов. Хромосомные плечи (p - короткое,q - длинное плечо) подразделяются на сегменты, которые в свою очередь нумеруются от центромеры. Например, 1p22.
Структурная организация хроматина.
Хроматин (хроматиновая нить) представляет собой интерфазное состояние хромосомы и отличается от последних степенью спирализации и, соответственно, длиной. Поэтому число хроматиновых нитей в соматических клетках должно соответствовать диплоидному набору хромосом. Хроматин - это функционально активное состояние хромосом.
Хроматин, также как и хромосома, неоднороден по своей структуре. Различают два типа хроматина: эухроматин и гетерохроматин, которые морфологически и функционально отличаются друг от друга. Эухроматин - это деспирализованные и функционально активные участки хроматина, в этих участках интенсивно происходят процессы транскрипции. Гетерохроматин - более спирализованные и функционально неактивные участки хроматина. Эти участки содержат незначительное количество структурных генов и, по существу, представляют собой участки хроматина временно или постоянно выключенные из процессов регуляции клеточной активности. В разных типах тканей и на различных этапах индивидуального развития чередование и расположение участков эухроматина и гетерохроматина определенной хроматиновой нити могут существенно отличаться. Возможно это является одним из механизмов клеточной дифференцировки. Нарушения хромосомного уровня организации наследственного материала связаны с изменениями структуры отдельной хромосомы в результате воздействия мутагенного фактора. При этом могут возникать как внутрихромосомные (делеция, инверсия), так и межхромосомные перестройки (транслокация, транспозиция).
10.Химический состав, особенности морфологии хромосом. Динамика их структур в клеточном цикле (интерфазные и метафазные хромосомы).
Морфология хромосом
При микроскопическом анализе хромосом прежде всего видны различия их по форме и величине. Строение каждой хромосомы сугубо индивидуальное. Можно заметить также, что хромосомы обладают общими морфологическими признаками. Они состоят из двух нитей — хроматид, расположенных параллельно и соединенных между собой в одной точке, названной центромерой или первичной перетяжкой. На некоторых хромосомах можно видеть и вторичную перетяжку. Она является характерным признаком, позволяющим идентифицировать отдельные хромосомы в клетке. Если вторичная перетяжка расположена близко к концу хромосомы, то дистальный участок, ограниченный ею, называют спутником. Хромосомы, содержащие спутник, обозначаются как АТ-хромосомы. На некоторых из них в тело-фазе происходит образование ядрышек. Концевые участки хромосом имеют особую структуру и называются теломерами. Теломерные районы обладают определенной полярностью, препятствующей их соединению друг с другом при разрывах или со свободными концами хромосом.
Участок хрома-тиды (хромосомы) от теломеры до центромеры называют плечом хромосомы. Каждая хромосома имеет два плеча. В зависимости от соотношения длин плеч выделяют три типа хромосом: 1) мета-центрические (равноплечие); 2) субметацентрические (неравноплечие); 3) акроцентрические, у которых одно плечо очень короткое и не всегда четко различимо.
На Парижской конференции по стандартизации кариотипов вместо морфологических терминов «метацентрики» или «акро-центрики» в связи с разработкой новых методов получения «полосатых» хромосом предложена символика, в которой всем хромосомам набора присваивается ранг (порядковый номер) по порядку убывания величины и в обоих плечах каждой хромосомы (р — короткое плечо, q — длинное плечо) нумеруются участки плеч и полосы в каждом участке по направлению от центромеры. Такая система обозначений позволяет детально описывать аномалии хромосом.
Наряду с расположением центромеры, наличием вторичной перетяжки и спутника важное значение для определения отдельных хромосом имеет их длина.
Для каждой хромосомы определенного набора длина ее остается относительно постоянной. Измерение хромосом необходимо для изучения их изменчивости в онтогенезе в связи с болезнями, аномалиями, нарушением воспроизводительной функции.
Тонкое строение хромосом. Химический анализ структуры хромосом показал наличие в них двух основных компонентов: дезоксирибонуклеиновой кислоты (ДНК) и белков типа гистонов и протомите (в половых клетках). Исследования тонкой субмолекулярной структуры хромосом привели ученых к выводу, что каждая хроматида содержит одну нить — хромонему. Каждая хро-монема состоит из одной молекулы ДНК. Структурной основой хроматиды является тяж белковой природы. Хромонема уложена в хроматиде в форму, близкую к спирали. Доказательства этого предположения были получены, в частности, при изучении мельчайших обменных частиц сестринских хроматид, которые располагались поперек хромосомы.
Химический состав хромосом
Изучение химической организации хромосом эукариотических клеток показало, что они состоят в основном из ДНК и белков, которые образуют нуклеопротеиновый комплекс—хроматин, получивший свое название за способность окрашиваться основными красителями.
Как было доказано многочисленными исследованиями (см. § 3.2), ДНК является материальным носителем свойств наследственности и изменчивости и заключает в себе биологическую информацию — программу развития клетки, организма, записанную с помощью особого кода. Количество ДНК в ядрах клеток организма данного вида постоянно и пропорционально их плоидности. В диплоидных соматических клетках организма ее вдвое больше, чем в гаметах. Увеличение числа хромосомных наборов в полипловдных клетках сопровождается пропорциональным увеличением количества ДНК в них.
Белки составляют значительную часть вещества хромосом. На их долю приходится около 65% массы этих структур. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки. Гистоны представлены пятью фракциями: HI, Н2А, Н2В, НЗ, Н4. Являясь положительно заряженными основными белками, они достаточно прочно соединяются с молекулами ДНК, чем препятствуют считыванию заключенной в ней биологической информации. В этом состоит их регуляторная роль. Кроме того, эти белки выполняют структурную функцию, обеспечивая пространственную организацию ДНК в хромосомах.
Число фракций негистоновых белков превышает 100. Среди них ферменты синтеза и процессинга РНК, редупликации и репарации ДНК. Кислые белки хромосом выполняют также структурную и регуляторную роль. Помимо ДНК и белков в составе хромосом обнаруживаются также РНК, липиды, полисахариды, ионы металлов.
РНК хромосом представлена отчасти продуктами транскрипции, еще не покинувшими место синтеза. Некоторым фракциям свойственна регуляторная функция.
Регуляторная роль компонентов хромосом заключается в «запрещении» или «разрешении» списывания информации с молекулы ДНК. Массовые соотношения ДНК: гистоны: негистоновые белки: РНК: липиды — равны 1:1:(0,2—0,5):(0,1—0,15):(0,01—-0,03). Другие компоненты встречаются в незначительном количестве.
Функции у интерфазной и метафазной хромосомы (ХР) разные. Интерфазная хромосома - это раскрученная двойная нить ДНК, в таком состоянии с нее считывается информация, необходимая для жизнедеятельности клетки. То есть функция интерфазнойХР - передача информации с генома, последовательности нуклеотидов в молекуле ДНК, для синтеза необходимых белков, ферментов и т. д.
Когда приходит время деления клетки необходимо сохранить всю имеющуюся информацию и передать ее в дочерние клетки. В состоянии "раздрая" ХР этого сделать не может . Поэтому хромосоме приходится структурироваться - скручивать нить своей ДНК в компактную структуру. ДНК к этому времени уже удвоена и каждая нить скручивается в свою хроматиду. 2 хроматиды образуют хромосому. В профазе под микроскопом в ядре клетки становятся заметны маленькие рыхлые комочки - это будущие ХР. Они постепенно укрупняются и формируют видимые хромосомы, которые к середине метафазы выстраиваются по экватору клетки. В норме в телофазе равное количество хромосом начинает двигаться к полюсам клетки.
Однако случается иногда, что хроматиды цепляются друг за друга, переплетаются, кусочки отрываются - а результате две дочерние клетки получают немного неравную информацию. Такая штука называется патологический митоз. После него дочерние клетки будуи работать неправильно. При сильном повреждении хромосом клетка погибнет, при более слабом не сможет разделиться еще раз или даст череду неправильных делений. Такие вещи приводят к возникновению заболеваний, от нарушений биохимической реакции в отдельной клетке, до заболевания раком какого-то органа. Клетки делятся во всех органах, но с разной интенсивностью, поэтому у разных органов - разная вероятность заболеть раком.

11.Кариотип человека.
Американский генетик Томас Морган в 1911 г. сформулировал хромосомную теорию наследственности, в которой впервые показал, что передача признаков по наследству связана с хромосомами.
Хромосомы — это самовоспроизводящиеся структурные элементы клеточного ядра, содержащие гены, предназначенные для хранения наследственной информации и правильного ее распределения в ходе митоза.
По форме хромосомы бывают в виде палочек, нитей, петель. В зависимости от расположения первичной перетяжки —центромеры, различают три типа хромосом: метацентрические, субметацентрические и акроцентрические.
В метацентрических—центромера расположена посередине.
Для субметацентрических— характерно наличие плечей разной длины.
Акроцентрические—центромера находится на конце хромосомы.
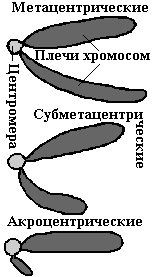
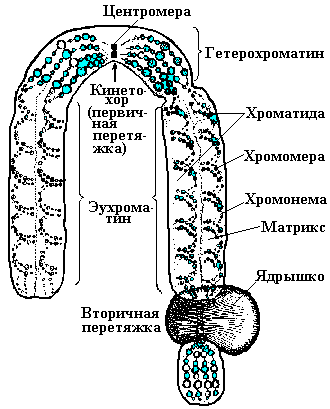
Концевые участки хромосомы называют теломерами. Особенность их состоит в том, что они не способны к соединению с другими участками хромосом.
Основными химическими компонентами хромосом эукариот являются ДНК, белки и небольшое количество РНК. Хромосомы способны менять свою структуру и длину на протяжении клеточного цикла. Так, в период интерфазы они находятся в деконденсированном состоянии и выполняют функции репликации и транскрипции. Максимальная конденсация хромосом характерна для делящейся клетки, особенно в метафазе. В период деления клетки хромосомы выполняют функцию перемещения и распределения наследственной информации.
Каждому виду характерен свой кариотип, т.е. определенное постоянное число, форма и размеры хромосом. В диплоидном наборе хромосом соматической клетки (его условно обозначают 2n) следует различать гомологичные хромосомы, которые имеют одинаковую морфологию, но происходят из разных геномов: одна от материнской гаметы, другая — от отцовской. Если пары гомологичных хромосом расположить в порядке убывания их размеров, то получится так называемаяидиограмма (рис. 6 Б).. У человека кариотип состоит из 46 хромосом. Причем различают 44 аутосомы и 2 половые хромосомы, которые отличаются у мужчин (ХУ) и у женщин (XX). Следовательно, кариотип мужского организма — 46,ХУ, женского — 46, XX.
В генетике человека широко используют цитогенетичеокий метод, с помощью которого изучают строение отдельных хромосом, а также особенности набора хромосом клеток человека в норме и патологии. По половому хроматину интерфазных ядер можно судить о состоянии половых хромосом, что позволяет провести экспресс-диагностику некоторых наследственных болезней.
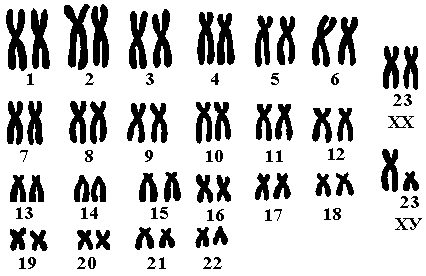
Денверская система классификации хромосом. Обычно классификация хромосом строиться на учете размера каждой из хромосом в кариотипе, по положению центромеры и по другим особенностям. Решениями конференций по хромосомам человека в Денвере США (Denverconference, 1960), в Лондоне (Londonconference, 1966) сведены обширные материалы из многочисленных литературных источников в систему, имеющую в настоящее время общепризнанный характер. Согласно этой системе, 22 пары аутосом были перенумерованы от 1 до 22-й номере уменьшения их длинны, пара половых хромосом обозначена символами Х и У. Кариотип мужчины - ХУ, женщины - ХХ. 22 пары аутосом разделены на семь групп, обозначаемых буквами от А до G. Каждая группа хромосом характеризуется следующими особенностями:
Группа А содержит 3 пары длинных хромосом (1-3), каждую из которых можно легко индивидуализировать. Хромосомы 1,3 являются метацентриками, аромосома 2 - субметацентрична;
Группа В содержит две пары хромосом (4-5). Они короче хромосом из группы А и являются субметацентриками;
Группа С содержит 6 пар аутосом (6-12), все хромосомы с субмедиальным расположением центромеры, средних размеров, их трудно индивидуализировать. К этой группе по размеру относится Х-хромосома, которая отличается тем, что заканчивает синтез ДНК позднее других;
Группа D содержит 3 пары хромосом (13-15). Хромосомы средних размеров имеют почти терминальное расположение центромеры - акроцентрики. Все они имеют спутники, морфологически похожи;
Группа Е состоит из 3 пар коротких хромосом (16-18). Хромосомы 16-й пары являются метацентриками. Хромосомы 17-й и 18-й пары, похожи между собой и являются субметацентриками;
Группа F имеет 2 пары коротких метацентрических хромосом (19-20), которые неотличимы друг от друга;
Группа G состоит из 2-х пар хромосом (21-22). Это очень короткие акроцентрические хромосомы со спутниками, трудно различимы, хотя несколько отличаются по величине и морфологии. К ним примыкают У-хромосома, которая несколько длиннее и имеет на длинном плече вторичную перетяжку (Дубинина, 1977).
В настоящее время для более тонкой дифференциации каждой из хромосом человека разработаны новые методы. Однако для исследования спонтанного хромосомного мутагенеза достаточно применения методики рутинной окраски хромосом, в результате которой все хромосомы перечисленных выше групп в исследуемой метафазной пластинке равномерно окрашиваются и хорошо идентифицируются.
20.Генная инженерия и биотехнологии
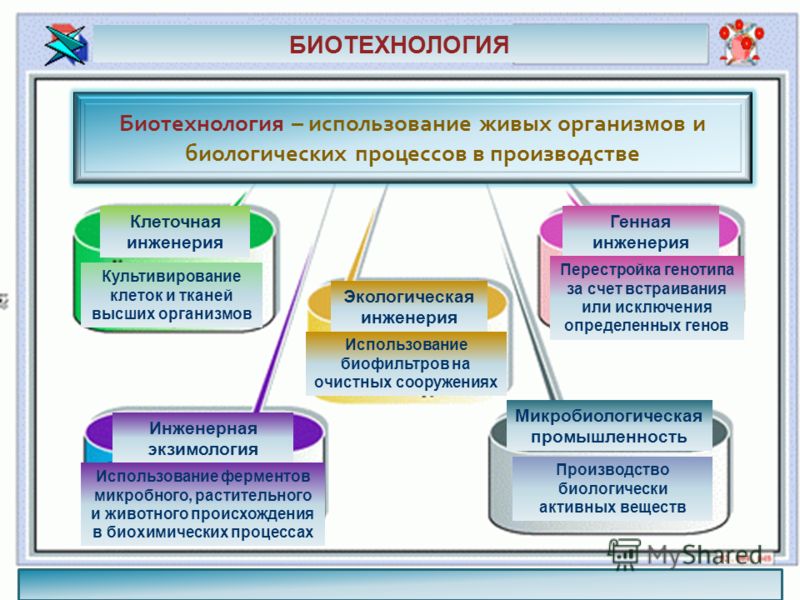
Биотехноло́гия — дисциплина, изучающая возможности использования живых организмов, их систем или продуктов их жизнедеятельности для решения технологических задач, а также возможности создания живых организмов с необходимыми свойствами методом генной инженерии.
Генетическая инжене́рия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами и введения их в другие организмы.
Рождение биотехнологии обусловлено потребностями общества в новых, более дешевых продуктах для народного хозяйства, в том числе для медицины и ветеринарии, а также принципиально новых технологиях. Целью биотехнологии являются получение продуктов из биологических объектов или с их применением, а также воспроизводство биоэффектов, не встречающихся в природе. В качестве биологических объектов чаще всего используются одноклеточные микроорганизмы, животные и растительные клетки, а также организм животных, человека или растений, так как вещества, синтезируемые этими организмами, недоступны пока человеку без использования биотехнологий.
Клетки чрезвычайно быстро воспроизводятся. Так, бактериальная клетка делится через каждые 20-60 мин, дрожжевая - через 1,5-2 ч, животная - через 24 ч, что позволяет за относительно короткое время искусственно нарастить на сравнительно дешевых и недефицитных питательных средах в промышленных масштабах огромные количества биомассы микробных, животных или растительных клеток.
Помимо микроорганизмов, животных и растительных клеток, биотехнология в качестве биологических объектов использует органы и ткани человека и животных, растения, организм животных и человека. Например, для получения инсулина используется поджелудочная железа крупного рогатого скота и свиней, гормона роста - гипофизы трупов человека, иммуноглобулинов - организм лошадей и других животных, препаратов крови - кровь доноров и т.д.
|
Помимо этого биотехнология играет большую роль в оздоровлении окружающей среды: с помощью биотехнологических процессов проводят очистку от загрязняющих веществ почвы, водоемов, воздушной среды путем их биоконверсии и биодеградации.
|
Из бактерий в биотехнологии чаще всего используют род Acetobacter - для превращение этанола в уксусную кислоту, углекислый газ и воду; родBacillus - для получения ферментов (B. subtilis), средств защиты растений (B. thuringiensis); род Clostridium - для сбраживания Сахаров в ацетон, этанол, бутанол; молочнокислые бактерии (Lactobacillus и др.); псевдомонады, например РР. denitrificans, - для получения витамина В12;Corynebacterium gentamicum - для получения аминокислот и др.
Из грибов в биотехнологии для получения разнообразных антибиотиков применяют род Streptomyces, Peniciliumchrysogenium, Cefalosporumacremonium, Streptomycesspp. и др.
Естественно, широкое применение в получении диагностикумов, вакцин, иммуноглобулинов, пробиотиков, фагов и других микробных препаратов находят патогенные и вакцинные штаммы болезнетворных микробов, а также условно-патогенные микроорганизмы.
Генетическая инженерия является сердцевиной биотехнологии. Она по существу сводится к генетической рекомбинации, т.е. обмену генами между двумя хромосомами, которая приводит к возникновению клеток или организмов с двумя и более наследственными детерминантами (генами), по которым родители различались между собой.
|
Экспрессируемый ген в виде рекомбинатной ДНК (плазмида, фаг, вирусная ДНК) встраивается в бактериальную или животную клетку, которая приобретает новое свойство - продуцировать несвойственное этой клетке вещество, кодируемое экспрессируемым геном.
В качестве реципиентов экспрессируемого гена чаще всего используют E. coli, B. subtilis, псевдомонады, нетифоидныесеровары сальмонелл, дрожжи, вирусы.
Методом генной инженерии созданы сотни препаратов медицинского и ветеринарного назначения, получены рекомбинантные штаммы-суперпродуценты, многие из которых нашли практическое применение. Уже используются в медицине полученные методом генной инженерии вакцины против гепатитаВ, интерлейкины-1, 2, 3, 6, инсулин, гормоны роста, интерфероны α, β, γ, фактор некроза опухолей, пептиды тимуса, миелопептиды, тканевый активатор плазминогена, эритропоэтин, антигены ВИЧ, фактор свертывания крови, моноклональные антитела и многие антигены для диагностических целей.

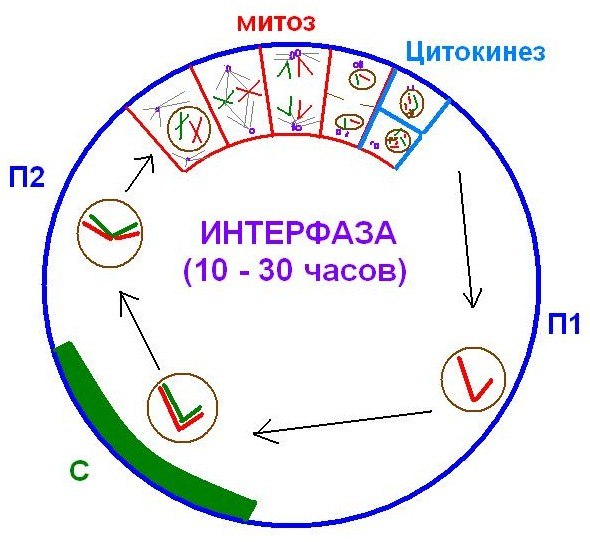
21. Временная организация клетки. Клточный цикл, его возможные направления и периодизация.
Время существования клетки от ее образования до следующего деления или смерти называют жизненным циклом клетки (ЖЦК). В ЖЦК эукариотических клеток многоклеточного организма можно выделить несколько периодов (фаз), каждый из которых характеризуется определенными морфологическими и функциональными особенностями:
- фаза размножения и роста
- фаза дифференцировки
- фаза нормальной активности
- фаза старения и смерти клетки.
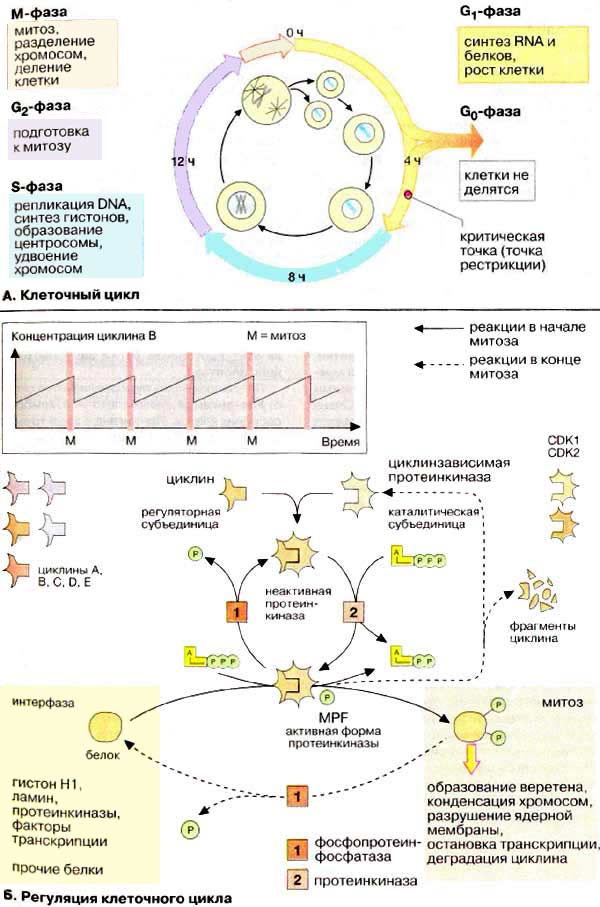
В жизненном цикле клетки можно также выделить митотический цикл, включающий подготовку клетки к делению и само деление. Основные принципы организации ЖЦК: 1. Продолжительность клеточного цикла различна в зависимости от типа клеток. 2. В нормальных клетках каждая стадия клеточного цикла зависит от правильного завершения предыдущей стадии( осуществляют ферментативные и белковые системы клеток).. 3. Усиление дифференцировки клеток сопровождается снижением их митотической активности.
Митоз – способ деления генетически и морфологически неизмененных клеток.. 1. Постмитотический (пресинтетический) период характеризуется ростом клетки, увеличением ее объема. В этой стадии следует выделить два взаимосвязанных явления:
- усиление процессов обмена веществ
- увеличение количества органоидов клетки
Варианты перехода в следующие стадии клеточного цикла: 1. Клетка вступает в митотический цикл, обязательным условием которого является репликация ДНК. Начинается синтетический период интерфазы. 2. Клетка прекращает рост и переходит в фазу дифференцировки и нормальной активности.
По степени специализации клетки можно разделить на недифференцированные и дифференцированные. Но только дифференцированные клетки могут полноценно выполнять свои функции. Поэтому любое нарушение дифференцировки приводит к нарушению или не выполнению.
Нормальное функционирование в составе органа – это основной период жизненного цикла клеток. Но неизбежным завершением ЖЦК является переход в фазу старения и гибели
Синтетический период (S-период)
Основной особенностью периода является репликация ДНК. Процесс репликации требует совместного действия многих белков: 1) ДНК-геликаза 2) дестабилизирующие белки (SSB – белки3)ДНК-полимераза 4)ДНК-лигазы 5) ДНК-топоизомеразы Основное значение репликации ДНК – удвоение наследственной информации, которая в последующем митозе будет равномерно распределена между дочерними клетками.
Механизмы контроля репликации стабильности ДНК.
1. Ферментативный контроль осуществляет ДНК-полимераза.
2. Репаративный контроль осуществляется особыми ферментативными системами на всех стадиях ЖЦК.
После полного завершения репликации и проверки правильности удвоения ДНК клетка переходит в следующий период – постсинтетический (премитотический, G2)
Особенности постсинтетического периода:
- накопление АТФ
- образование белков веретена деления
- синтез в клетке М-стимулирующего фактора.
Митоз – непрямое деление эукариотической клетки, в результате которого образуются клетки идентичные по кариотипу и генотипу. Кариотип – набор хромосом клетки. Генотип – совокупность генов диплоидной клетки.
Термин «Соматические мутации» - мутации связанные с нарушением наследственного аппарата диплоидных клеток. При этом уровень нарушения может быть различным: генный, хромосомный, геномный.
Регуляция митотического цикла.
Изучение механизмов регуляции митозов является одной из важнейших проблем современной биологии, поскольку митозы лежат в основе роста организмов, регенерации тканей, нарушение митотической активности – основной механизм возникновения опухолей.
В регуляции митотического цикла участвуют различные факторы, их сбалансированное взаимодействие приводит к нормальному протеканию клеточного цикла.
В регуляции периодов интерфазы принимают участие ранее упомянутые факторы: активаторы S-периода, М-стимулирующий и М-задерживающий факторы, циклины.
Старение и гибель клеток. После определенного периода нормального функционирования у клетки начинается период старения, который морфологически проявляется:
- уменьшением объема клетки
- увеличением содержания крупных лизосом
- накоплением пигментных и жировых включений
- появлением вакуолей в цитоплазме и ядре
Гибель клетки – завершающий этап клеточного цикла
При гибели клетки можно выделить два различных механизма ее развития: некроз и апоптоз.
22. Деление клетки. Понятие митотической активности. Нарушения митоза.
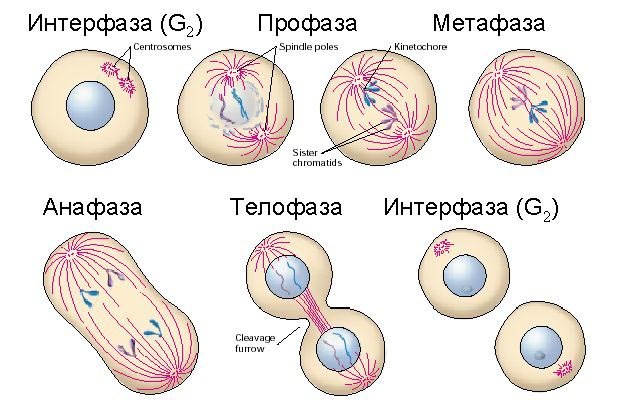
Деление клетки — процесс образования из родительской клетки двух и более дочерних клеток
Существует два способа деления ядра эукариотических клеток: митоз и мейоз.
Мейоз - это особый способ деления клеток, в результате которого происходит редукция (уменьшение) числа хромосом вдвое. С помощью мейоза образуются гаметы. В результате редукции споры и половые клетки хромосомного набора получают в каждую гаплоидную спору и гамету по одной хромосоме из каждой пары хромосом, имеющихся в данной диплоидной клетке. В ходе дальнейшего процесса оплодотворения (слияния гамет) организм нового поколения получит опять диплоидный набор хромосом, т. е. кариотип организмов данного вида в ряду поколений остается постоянным.
Митоз —(реже: кариокинез или непрямое деление) — деление ядра эукариотической клетки с сохранением числа хромосом. В отличие от мейоза, митотическое деление протекает без осложнений в клетках любой плоидности, поскольку не включает как необходимый этап, конъюгацию, хромосом в профазе. Митоз (от греч.Mitos- нить) непрямое деление, - основной способ деления эукариотических клеток. Митоз - это деление ядра, которое приводит к образованию двух дочерних ядер, в каждом из которых имеется точно такой же набор хромосом, что и в родительском ядре. Вслед за делением ядра обычно следует деление самой клетки, поэтому часто термином - «митоз» обозначают деление клетки целиком.
МИТОТИЧЕСКАЯ АКТИВНОСТЬ (наличие делящихся клеток) ткани подразделяются на пролиферирующие (где есть размножение клеток) и непролиферирующие, или дифференцированные (где клетки приобрели окончательную специализацию и утратили способность к делению). Высокую митотическую активность клеток в ткани связывают с накоплением в ней гликогена (Bullough, 1949). При понижении количества гликогена интенсивность деления клеток снижается. Л. В. Суворова (1955) нашла, что у незрело-рождающихся животных на ранних этапах постнатального развития этот эффект отсутствует. В этот период в тканях как раз преобладают процессы гликолиза и низка активность окислительных ферментов.
Нарушения митоза
Правильное течение митоза может быть нарушено различными внешними воздействиями: высокими дозами радиации, некоторыми химическими веществами. Например, под действием рентгеновых лучей ДНК хромосом может разорваться. Хромосомы в таком случае тоже разрываются. При этом могут возникнуть хромосомы без центромерного района. Такие хромосомы лишены способности двигаться в прометафазе и анафазе. В зависимости от того, в каком месте ядра бесцентромерная хромосома находилась накануне деления,будет складываться ее дальнейшая судьба. Если хромосома была смещена к одному из полюсов клетки, то при формировании дочерних клеток она может целиком включиться в одну из них, т. е. обе сестринские хроматиды окажутся в одном ядре. Одни хромосомы начинают двигаться быстрее, другие отстают. Отставшие хромосомы могут не включиться в формирующиеся дочерние ядра. Иногда в делящейся клетке образуется не два, а три или четыре полюса, что ведет к возникновению соответственно трех или четырех дочерних клеток. При таком делении нарушается весь слаженный механизм распределения хромосом. Метафазная хромосома, состоящая из двух сестринских хроматид, может взаимодействовать одновременно только с двумя полюсами. Если полюсов больше, то каждая хромосома вынуждена "выбирать", с какими двумя полюсами из трех или четырех ей взаимодействовать. Этот выбор совершается случайно. В результате каждая дочерняя клетка получает не весь набор хромосом, а только его часть. Клетки, получившие неполный набор хромосом, как правило, оказываются нежизнеспособными и погибают. Изучение нарушений митоза, вызванных различными факторами, с одной стороны, помогает лучше понять митотические процессы, с другой - позволяет устанавливать механизмы повреждающего действия этих факторов и, следовательно, создает условия для целенаправленного поиска методов устранения таких нарушений.
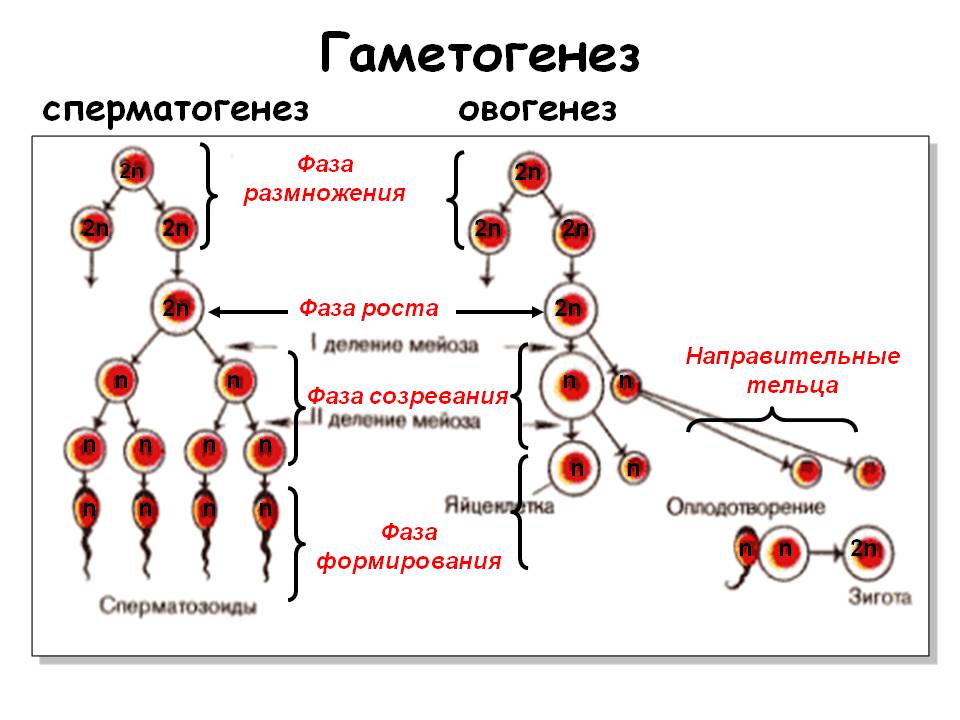
25. Гаметогенез. Мейоз.
Гаметогенез или предзародышевое развитие — процесс созревания половых клеток, или гамет. Поскольку в ходе гаметогенеза специализация яйцеклетоки сперматозоидов происходит в разных направлениях, обычно выделяют оогенез (устаревшее название — овогенез в современных изданиях по эмбриологии больше не употребляется) и сперматогенез.
Гаметогенез закономерно присутствует в жизненном цикле ряда простейших, водорослей, грибов, споровых и голосеменных растений, а также многоклеточных животных. В некоторых группах гаметы вторично редуцированы (сумчатые и базидиевые грибы, цветковые растения). Наиболее подробно процессы гаметогенеза изучены у многоклеточных животных.
Мейо́з или редукционное деление клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом — образованием специализированных половых клеток, или гамет, из недифференцированных стволовых.
С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности(переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса.
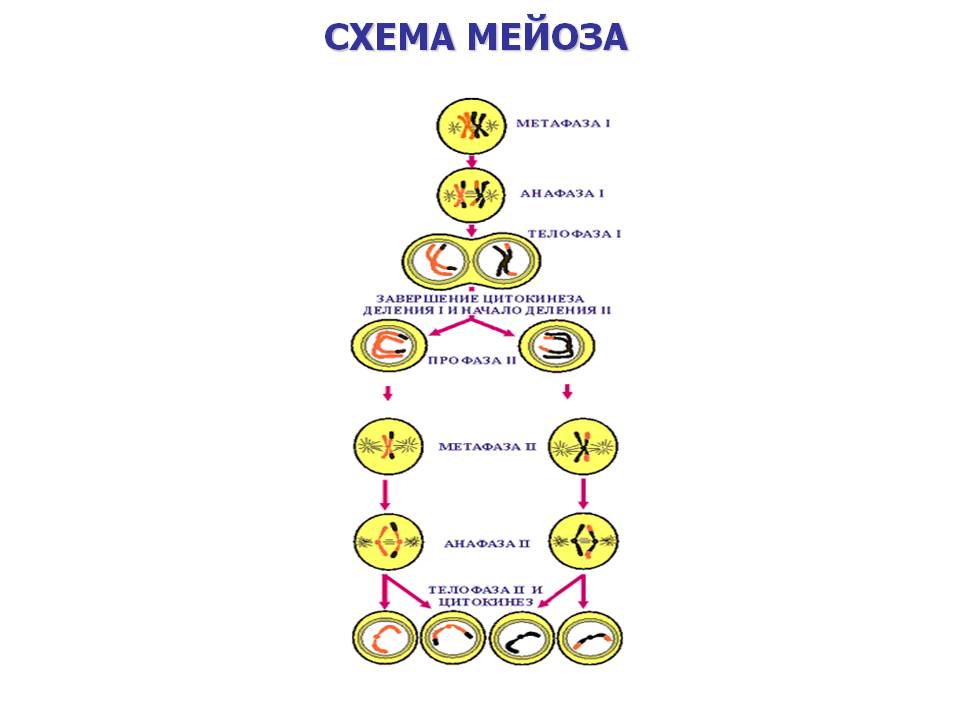
Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
Профаза I — профаза первого деления очень сложная и состоит из 5 стадий:
Лептотена или лептонема — упаковка хромосом, конденсация ДНК с образованием хромосом в виде тонких нитей (хромосомы укорачиваются).
Зиготена или зигонема — происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами и их дальнейшая компактизация.
Пахитена или пахинема — (самая длительная стадия) — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них происходит кроссинговер — обмен участками между гомологичными хромосомами.
Диплотена или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток.
Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой.
К концу Профазы I центриоли мигрируют к полюсам клетки, формируются нити веретена деления, разрушаются ядерная мембрана и ядрышки
Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки.
Анафаза I — микротрубочки сокращаются, биваленты делятся, и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе.
Телофаза I — хромосомы деспирализуются и появляется ядерная оболочка.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
Профаза II — происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления, перпендикулярное первому веретену.
Метафаза II — унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку.
Анафаза II — униваленты делятся и хроматиды расходятся к полюсам.
Телофаза II — хромосомы деспирализуются и появляется ядерная оболочка.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки.
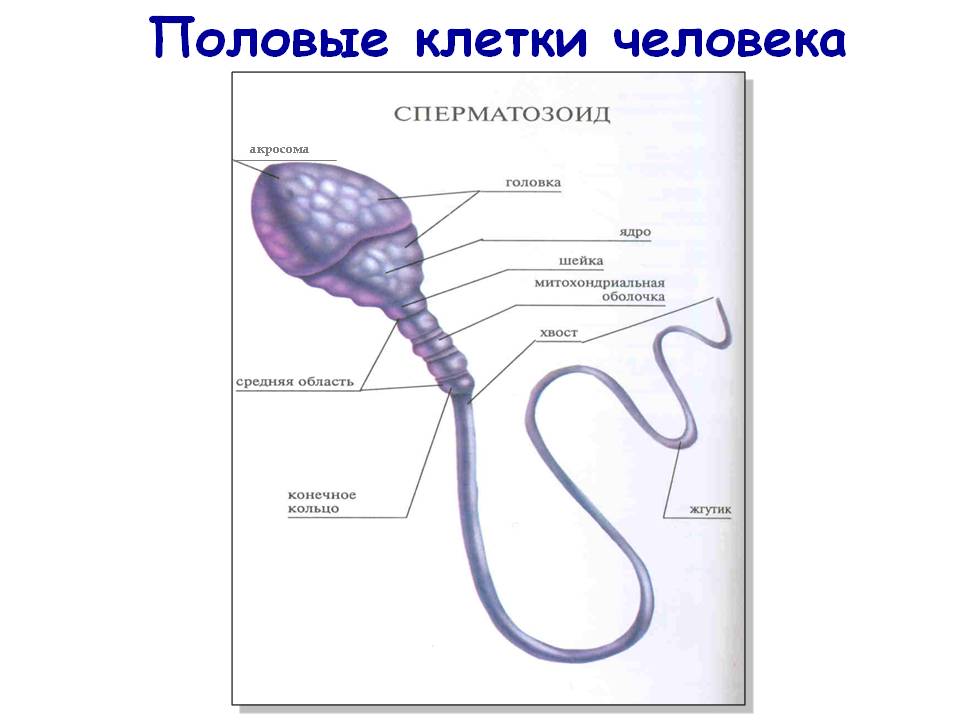
26. Половые клетки человека.
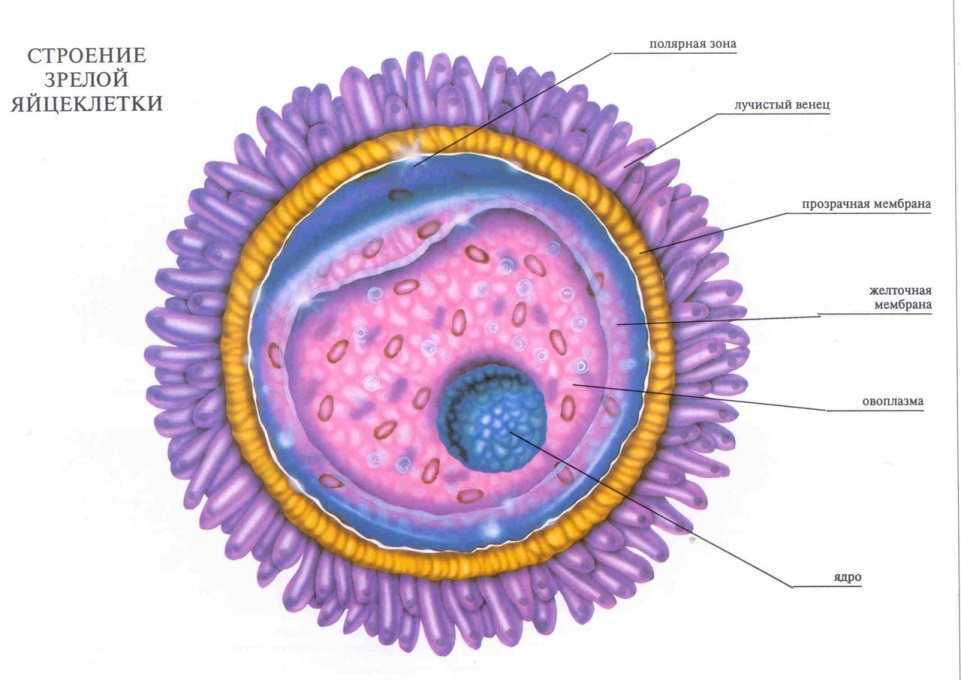
Гаметы - это половые клетки: яйцеклетки (женские гаметы) и сперматозоиды (мужские гаметы), которые обеспечивают передачу наследственной информации от родителей к потомкам. Гаметы представляют собой высокодифференцированные клетки. В процессе эволюции они приобрели свойства выполнение специфических функций. Ядра как мужских, так и женских гамет содержат одинаковую наследственную информацию, необходимую для развития организма. Однако другие функции яйцеклетки и сперматозоида разные, поэтому по строению они очень различаются.
Яйцеклетки неподвижны, шаровидной или слегка удлиненной формы. Они содержат все типичные клеточные органеллы, но по строению отличаются от других клеток, так приспособлены для реализации развития целого организма. Яйцеклетки гораздо больше, чем соматические клетки. Внутриклеточная структура цитоплазмы специфическая для каждого вида животных, чем обеспечиваются видовые (а часто и индивидуальные) особенности развития. В яйцеклетках содержатся вещества, необходимые для развития зародыша. К ним относится питательный материал (желток). В некоторых видов животных накапливается в яйцеклетках столько желтка, которые можно увидеть невооруженным глазом (икринки рыб и земноводных, яйца пресмыкающихся и птиц). Из современных животных самые яйцеклетки вселедочной акулы (29 см в диаметре). У птиц яйцом считается то, что в быту называется "желтком"; диаметр яйца страуса 10,5 см, курицы - около 3,5 см.
Например, у ракообразных они должны вырасти в виде лучей или отростков, в круглых червей - форму шарообразных или овальных телец т.д. Таким образом, половые клетки существенно отличаются от соматических клеток:
1) в половых клетках гаплоидный набор хромосом, в соматических - диплоидный;
2) в половых клетках ядерно-цитоплазматическое соотношение разное: в сперматозоидах оно высокое, в яйцеклетке - низкое,
3) форма и размеры половых клеток иные, чем в соматических;
4) половые клетки отличаются низким уровнем обменных процессов;
5) для яйцеклеток характерна цитоплазматическая сегрегация закономерный перераспределение цитоплазмы после оплодотворения). Относительно большим ядром и значительным количеством цитоплазмы.
27.Особенности репродукции человека. Оплодотворение Репродукция(размножение) человека – это физиологическая функция, необходимая для сохранения человека как биологического вида. Процесс размножения у человека начинается с зачатия (оплодотворения), т.е. с момента проникновения мужской половой клетки (сперматозоида) в женскую половую клетку (яйцеклетку).
Способность к репродукции становится возможной после полового созревания. Признаком наступления полового созревания у человека являются первые поллюции у мальчиков и первые менструации у девочек. Половая зрелость наступает у лиц женского пола в возрасте 16-18 лет, мужского – 18-20 лет. Сохраняется способность к репродукции у женщин до 40-45 лет, а у мужчин до старости. Продукция гамет у представителей обоих полов различна: зрелый семенник непрерывно вырабатывает огромное количество сперматозоидов; половозрелый яичник периодически (один раз в лунный месяц) выделяет зрелую яйцеклетку, созревающую из числа овоцитов, которые закладываются на ранних этапах онтогенеза и запасы которых убывают в течение жизни женщины. Значение того, что овоциты закладываются еще до рождения состоит в том, что потомство, появляющееся к концу репродуктивного периода, развивается из овоцитов, в которых за жизнь женщины могли возникнуть генетические дефекты.
Следствием этого является то, что у пожилых матерей относительно чаще рождаются дети с врожденными дефектами. Необходимо подчеркнуть, что основную опасность представляет не сам возраст матери, а мутагенные факторы и факторы, влияющие на развитие плода. У человека, как и у других организмов, имеющих внутреннее оплодотворение, мужские половые клетки при половом акте вводятся в половые органы женщины. Во время извержения семенной жидкости (эякуляция) у человека выделяется около 100-200 млн сперматозоидов, но только один из них оплодотворяет яйцеклетку.
Оплодотворение Встреча женских и мужских гамет происходит в верхних отделах маточных труб. Верхних отделов маточной трубы достигает около 100 сперматозоидов. Перемещение их осуществляется благодаря собственной подвижности, а также в результате мышечных сокращений стенок полового тракта и направленного движения ресничек слизистой оболочки маточных труб. Сперматозоиды сохраняют способность к оплодотворению в течение 1-2 суток, яйцеклетки – на протяжении суток после овуляции. Оплодотворение осуществляется обычно в течение первых 12 часов после овуляции. В процессе проникновения сперматозоида через барьер фоликуллярных клеток, окружающих яйцеклетку и её оболочку, большую роль играет акросомная реакция. Вслед за проникновением сперматозоида в яйцеклетку образуется оболочка оплодотворения, препятствующая проникновению других сперматозоидов. Зигота опускается по маточным трубам и на восьмые- десятые сутки зародыш внедряется в стенку матки. Если оплодотворение не наступило, яйцеклетка удаляется из организма.

29.Этапы эмбрионального развития
Период эмбрионального развития наиболее сложен у высших животных и состоит из нескольких этапов.
Период начинается с этапа дробления зиготы, т. е. серии последовательных митотических делений оплодотворенной яйцеклетки. Образующиеся в результате деления две клетки (и все последующие их поколения) на этом этапе называются бластомерами. Одно деление следует за другим, причем не происходит роста образующихся бластомеров и с каждым делением клетки становятся все более мелкими. Такая особенность клеточных делений и определила появление образного термина «дробление зиготы»
В результате дробления (когда количество бластомеров достигнет значительного числа) образуется бластула. Часто она представляет собой полый шар (например, у ланцетника), стенка которого образована одним слоем клеток — бластодермой. Полость бластулы — бластоцель, или первичная полость, заполнена жидкостью.
На следующем этапе осуществляется процесс гаструляции — формирование гаструлы. У многих животных она образуется путем впячивания бластодермы внутрь на одном из полюсов бластулы при интенсивном размножении клеток в этой зоне. В результате и возникает гаструла.
Наружный слой клеток получил название эктодермы, а внутренний — энтодермы. Внутренняя полость, ограниченная энтодермой, полость первичной кишки сообщается с внешней средой первичным ртом, или бластопором. Существуют и другие типы гаструляции, но у всех животных (кроме губок и кишечнополостных) этот процесс завершается образованием еще одного клеточного пласта — мезодермы. Она закладывается между энто- и эктодермой.
По завершении этапа гаструляции появляются три клеточных пласта (экто-, эндо- и мезодерма), или три зародышевых листка.
30.Критические периоды эмбриогенеза человека. Тератогенные факторы.
Критические периоды эмбриогенеза это временные промежутки, характеризующиеся повышенной чувствительностью плода к влиянию повреждающих факторов.
Критические, потому что вероятность возникновения какой-либо патологии в эти периоды очень высока.
В эмбриональном развитии человека имеются следующие3 основных критических периода.
1. Имплантация (6 – 7-е сутки после зачатия) – внедрение зиготы в стенку матки.
2. Плацентация (конец 2-й недели беременности) – образование у эмбриона плаценты.
3. Перинатальный период (роды) – переход плода из водной в воздушную среду через 9 месяцев после зачатия.
С критическими периодами в организме новорожденного связаны резкое изменение условий существования и перестройка деятельности всех систем организма (изменяется характер кровообращения, газообмена, питания). Наиболее уязвимым эмбрион становится в первом и втором критических периодах онтогенеза.
Существует масса неблагоприятных факторов, вызывающих вышеперечисленные нарушения у плода. Это, например, интоксикация материнского организма, диабетическая и алкогольная фетопатии, заболевания эндокринной системы, воздействие физических (температура, ионизирующее излучение, механическое воздействие), химических (медикаменты, алкоголь, курение, наркотики, пары ртути, толуол, хлордифенил) и биологических (инфекционные болезни, вирусы) факторов. Такие факторы называются тератогенными - факторы, которые могут привести к нарушению развития плода и его органов.
Основные группы тератогенных факторов.
1. Лекарственные средства и химические вещества.
2. Ионизирующее излучение.
3. Инфекции.
4. Метаболические нарушения и вредные привычки у беременной.


31. Старение как этап онтогенеза. Теории старения.
Старение - это универсальный и закономерный процесс, которому присуща последовательность, прогрессирование, которое приводит к снижению адаптационных возможностей и жизнеспособности индивида. Старение касается всех уровней организации: от молекулярных структур до целостного организма.
К наиболее характерным:
1. Уменьшение роста (на 0,5-1 см за пятилетие после 60)
2. Изменение формы тела (сглаживание контуров, усиление кифоза, перераспределение жирового компонента)
3. Снижение массы тела, упругости и эластичности мышц
4. Снижение амплитуды движений грудной клетки
5. Снижение плавности движений
6. Уменьшение размеров лица впоследствии потери зубов и редукции альвеолярных отростков челюстей
7. Увеличение объёма мозговой части черепа, ширины носа и рта
8. Изменения в коже (уменьшение количества сальных желез, толщины эпидермиса)
Для процесса старения характерны также изменения в функционировании важных систем организма:
1. В ЦНС наблюдаются структурные (уменьшение массы мозга, величины и плотности нейронов) и функциональные (снижение работоспособности нейронов)
2. Снижение остроты зрения, функции слухового аппарата, вкуса, части кожной чувствительности)
3. Уменьшение массы желез, снижение их гормонообразовательной функции (щитовидная, половых желез)
4. Снижается секреторная активность пищеварительных органов, жизненная ёмкость лёгких, основных функций почек, сокращающая целостность миокарда, ритмическая деятельность сердца уменьшается
5. Резко снижается иммунный гомеостаз, количество и функциональная активность Т-лимфоцитов (снижение активности иммунной системы приводит к развитию аутоиммунных процессов, растёт возможность образования опухолей)
Старение - длительный процесс, последним этапом которого является старость.
Французский биолог Ж.Бюффон рассчитывал, что длительность жизни человека должна превышать длину его роста в 6-7 раз и составляет около 90-100 лет. Все последующие расчёты по разным критериям были близки к этой цифре(максимальная длительность жизни человека достигает 130 лет).
У человека различают:
- физиологическую старость, связанную с календарным возрастом
- преждевременноесатрение, обусловленное социальными факторами и болезнями.
60-75 лет - пожилой
74 лет и более - старость
Теории старения:
1) одна из наиболее известных первых теорий: теория немецкого врача Х.Гуфеланде, который придавал в долголетии значение трудовой деятельности. Он говорил, что ни один лентяй не дожил до преклонного возраста.
2) эндокринная теория Бертольда, который показал, что пересадка семенников от одних животных к другим сопровождается развитием вторичных половых признаков. В начале 20в. уже сложилось убеждение о том, что наступление старости связано с угасанием деятельности желез внутренней секреции, в частности половых желез. На основе этого убеждения в разных странах было проделано много операций по омолаживанию пожилых или старых людей. Все эти операции приводили к некоторым эффектам, но лишь временным. После таких воздействий процессы старения продолжались, причём ещё интенсивнее.
3) микробиологическая теория старения И.И.Мечникова, различавшего физиологическую старость и патологическую. Он считал, сто старость человека является патологической, т.е. преждевременной. Основу представлений И.И.Мечникова составляло учение об ортобиозе, в соответствии с которой основной причиной старения является повреждение нервных клеток продуктами интоксикации, образующимися в результате гниения в толстьм кишечнике. Развивая учение о нормальном образе жизни (соблюдение правил гигиены, регулярный труд, воздержание от вредных привычек), И.И.Мечников предлагал также способ подавления гнилостных бактерий кишечника путём употребления кисломолочных продуктов.
4) теория о роли ЦНС в старении И.П.Павлова, который утановил интегрирующую роль ЦНС в нормальном функционировании организмов. Последователи И.П.Павлова в экспериментах на животных показали, что преждевременное старение вызывается нервными потрясениями и продолжитьным нервным потрясением.
5) теория возрастных изменений соединительной ткани, сформулированная А.А.Богомольцем.он считал, что физиологическую активность организма обеспечивает соединительная ткань (костная, хрящи, сухожилия, связки и волокнистая соединительная ткань) и что изменения строения клеток определяют возрастные изменения.
Наиболее распространенные современные представления о механизме старения сводятся к тому, что в процессе жизнедеятельности в клетках организма накапливаются соматические мутации, в результате которых происходит синтез дефектных белков или нерепарируемые сшивки ДНК с белком. Поскольку дефектные белки играют деезинтегрирующую роль в клеточном метаболизме, то это ведёт к старению. Однако исчерпывающей теории старения все же не создано, поскольку ясно, что ни одна из этих теорий самостоятельно объяснить механизмы старения не может.

33.Клиническая и биологическая смерть.
Клиническая смерть – это период, когда все угасшие жизненно важные процессы организма еще обратимы. Клиническая смерть может быть вызвана обильным кровотечением, электротравмой, утоплением, рефлекторной остановкой сердца, острым отравлением и т.д. Признаки клинической смерти:
отсутствие пульса на сонной или бедренной артерии(основной признак остановки кровообращения),
отсутствие дыхания,
потеря сознания,
широкие зрачки и отсутствие их реакции на свет.
Биологи́ческая смерть (или истинная смерть) - необратимое прекращение физиологических процессов в клетках и тканях. Связана с прекращением самообновления, химические процессы становятся неупорядоченными, в клетках происходит аутолиз (самопереваривание), разложение. Скорость процессов определяется степенью чувствительности. Наиболее чувствительные - нервные клетки (изменения происходят уже через 5-6 минут).
К ранним признакам биологической смерти относятся:
Отсутствие реакции глаза на раздражение (надавливание)
Помутнение роговицы
Появление симптома «кошачьего глаза»: при боковом сдавлении глазного яблока зрачок трансформируется в вертикальную веретенообразную щель, похожую на кошачий зрачок.
Отсутствие пульса.
В дальнейшем обнаруживаются трупные пятна с локализацией в отлогих местах тела, затем возникает трупное окоченение, разложение. Трупное окоченение обычно начинаются с мышц лица, верхних конечностей.


34.Регенерация и её виды.
Регенерация – процесс восстановления утраченных или поврежденных тканей или органов.
Различают физиологическую, репаративную и патологическую регенерацию.
Физиологическая регенерация свойственна всем организмам. Процесс жизнедеятельности обязательно включает два момента – утрату и восстановление морфологических структур на клеточном, тканевом, органном уровнях. У млекопитающих и человека непрерывно слущиваются наружные слои кожного эпителия, продолжительность жизни кишечного эпителия составляет несколько дней. Сравнительно быстро происходит смена эритроцитов.
Репаративная регенерация возникает, когда в организме происходит повреждение и гибель клеток и тканей. Она широко распространена, но способность к ней выражена неодинаково у различных животных. Репаративная регенерация может быть типичной(гомоморфоз) и атипичной(гетероморфоз). При гомоморфозе восстанавливается такой же орган, как и утраченный, при гетероморфозе восстановленные органы отличаются от типичных.
Восстановление осуществляется путем эпиморфоза, морфаллаксиса и эндоморфоза.
Эпиморфоз – рубцевание, безвосстановление органа. Морфаллаксис – разрастание оставшейся части, завершается образованием организма или органа. Эндоморфоз (регенерационная гипертрофия) – восстановление внутри органа.
Патологическая регенерация – разрастание тканей, не идентичных здоровым тканям в данном органе. Например, на месте глубоких ожогов может быть массивное разрастание плотной соединительной рубцовой ткани, а нормальная структура кожи не восстанавливается.

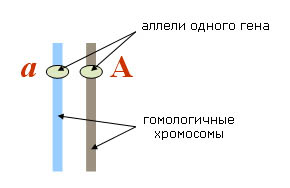
40. Взаимодействие аллельных и неаллельных генов
Взаимодействиеаллельныхгенов:
1) Полное доминирование - взаимодействие аллельных генов, при котором доминантный ген полностью подавляет фенотипическое проявление рецессивного гена. Например, жёлтый цвет гороха полностью подавляет зелёный.
2) Неполное доминирование - когда оба аллельных гена и доминантный, и рецессивный проявляются фенотипически в результате чего гибридное потомство имеет промежуточный фенотип. Например, у цветов АА - красный, аа - белый, а Аа - розовый.
3) Кодоминантность - вид взаимодействия, когда оба гена проявляют каждый себя самостоятельно в равной степени.
4) Сверхъдоминирование - заключается в том, что у доминантного аллеля в гетерозиготном состоянии иногда отмечается более сильное проявление, чем в гомозиготном состоянии.
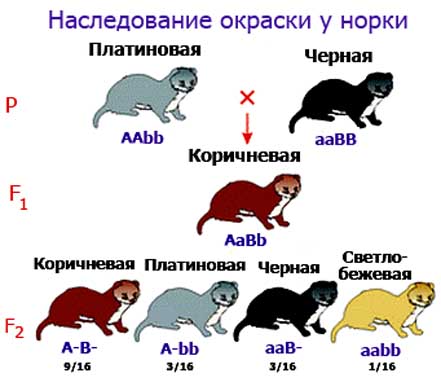
Взаимодействие неаллельных генов:
1) Комплементарное действие - когда для формирования признака необходимо наличие двух или более неаллельных (обычно доминантных) генов. Например, у человека нормальный слух обусловлен двумя доминантными генами Д и Е, из которых один определяет развитие улитки, а другой - слухового нерва. Доминантные гомозиготы и гетерозиготы по обоим генам имеют нормальный слух, рецессивные по одному из этих генов - глухие.
2) Эпистаз - подавление неаллельным геном действия другого гена, названного гипостатическим.
3) Полимерия - заключается в том, что различные неаллельные гены могут оказывать действие на один и тот же признак, усиливая его проявление. Такие гены получили название полимерных, а признаки ими определяемые - пол генных. Так цвет кожи у человека определяется 5ю парами аллельных генов.
4) Плейотропия - зависимость нескольких признаком от одного гена, т.е. проявление множественных эффектов одного гена, явление обратное полимерии. Например, болезнь Марфана, арахнодактилия - наследственноезаболение человека. Гены, определяющий это заболевание, вызывают нарушение развития соединительной ткани и оказывает влияние одновременно на развитие нескольких признаков: нарушение в строении хрусталика глаза, аномалии в ССС.
\
41. Сцепленное наследование
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
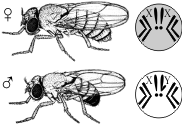
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
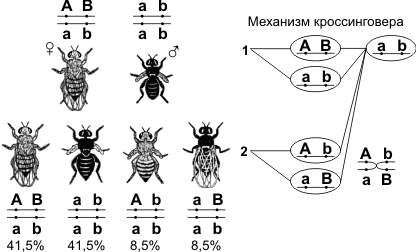
1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
|
Образуются гаметы: |
|
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правилокроссоверные гаметы составляют небольшую часть от всего количества гамет.
|
Образуются гаметы: |
|
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
43.Генетика пола
В хромосомном наборе у женщин имеется 22 пары аутосом и две одинаковые половые хромосомы XX; у мужчин 22 пары аутосом и половые хромосомы X и V (неодинаковые). В процессе мейоза каждая из пары гомологичных хромосом уходит в разные гаметы. Так как у женщин 23 пары гомологичных хромосом, то во все гаметы попадает 22 аутосомы и одна Х-хромосома (гаметы одинаковы), поэтому женский пол гомогаметный. У мужчин образуется два типа гамет: 22+Х и 22+Y, поэтому мужской пол гетерогаметный. Вероятность рождения девочек так же, как и мальчиков, составляет 50 %.
P - ♀22×2 + XX × 22×2 + XY♂
G – 22+X 22+X 22+Y
F - 22×2 + XX 22×2 + XY
50% девочки 50% мальчики
Пол будущего ребенка определяется сочетанием половых хромосом в момент оплодотворения. Если яйцеклетку оплодотворяет сперматозоид с Х- хромосомой, то рождается девочка, а если яйцеклетку оплодотворяет сперматозоид с Y-хромосомой, то рождается мальчик. Признаки организма, связанные с полом, подразделяют на три категории.
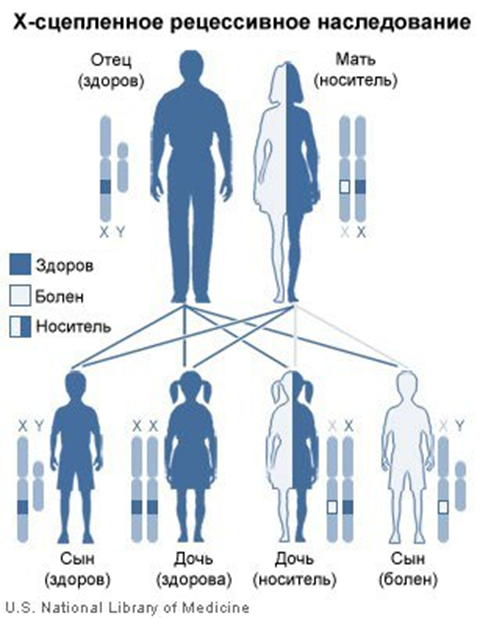
1)признаки, сцепленные с половыми хромосомами. Это признаки, развитие которых обусловлено генами, расположенными в одной из половых хромосом. Х - хромосома по размерам значительно больше, чем Y - хромосома.
В Х-хромосоме есть большой участок, которого нет в Y-хромосоме, в нем расположены рецессивные гены (гемофилии, дальтонизма, миопатии и доминантные (рахита, не поддающиеся лечению витамином D, темной эмали зубов). Гены этого участка полностью сцеплены с полом, передаются исключительно через Х-хромосому. Доминантные гены из этого участка одинаково проявляются у обоих полов; рецессивные — у женщин только в гомозиготном состоянии, а у мужчин - в гемизиготном состоянии.
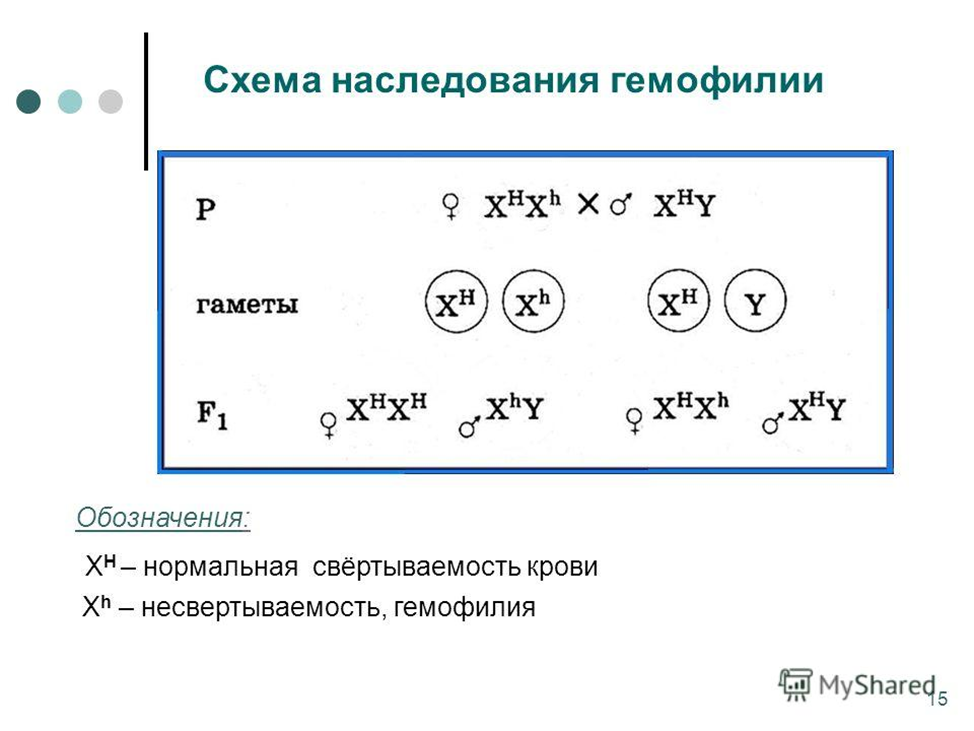
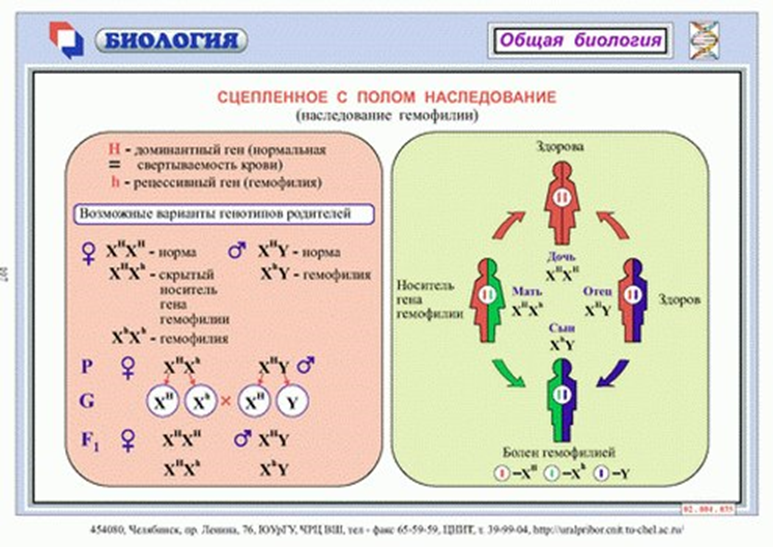
Рассмотрим наследование, сцепленное с полом, на примере гемофилии. У мужчин ген гемофилии находится в гемизиготном состоянии (один из пары), и поэтому гемофилия у мужчин всегда проявляется. У женщин, если и есть, этот ген в гетерозиготном состоянии, он не проявляется, так как доминантный ген нормальной свертываемости его подавляет. В брак вступают здоровая женщина-носительница и здоровый мужчина:
H-ген нормальной свертываемости крови
h-ген гемофилии
P-♀ XHXh × XHY♂
G- XHXhXHY
F - XHXH ,XHXh , XHY, XhY
XHXH -девочка здоровая
XHXh - девочка носитель
XHY - мальчик здоровый
XhY - мальчик больной
В непарном участке У-хромосомы, кроме генов, детерминирующих пол, расположено небольшое число генов, которые могут встречаться только у лиц мужского пола и передаются от отца всем сыновьям (голандрические гены): волосатость ушей, перепонки между пальцами ног, ихтиоз (кожа в виде рыбьей чешуи).
В парном сегменте (гомологичном для Х- и У-хромосом) локализованы гены, детерминирующие пигментную ксеродерму, болезнь Огучи, спасти¬ческую параплегию, эпидермолиз буллезный, общую цветовую слепоту и другие. Их "называют неполно или частично сцепленными с полом. Они могут передаваться как с Х-, так и с У-хромосомой;
2) признаки, контролируемые полом. Развитие этих признаков обусловлено генами, расположенными в аутосомах обоих полов, но степень проявления их (экспрессивность) разная у мужчин и женщин (особенно у гетерозигот, так как происходит сдвиг доминантности). Изменение доминирования гена обусловлено половыми гормонами. Например: а) у мужчин ген облысения доминантный, а у женщин — рецессивный и проявляется только в гомози¬готном состоянии, поэтому лысых мужчин значительно больше, чем женщин; б) пенетрантность гена подагры у мужчин 80 %, а у женщин 12 %, значит, подагрой чаще болеют мужчины;
3) признаки, ограниченные полом. Развитие этих признаков обусловлено генами, расположенными в аутосомах обоих полов, но проявляются они только у одного пола (гены удойности и жирности молока имеются у коров и быков, но проявляются только у коров).
44.Изменчивость и ее формы
Изменчивость – способность организмов приобретать новые или утрачивать прежние признаки или свойства.
Степень фенотипического проявления данного гена называется экспрессивностью, а частота его проявления - пенетрантностью. С изменчивостью связаны явления фенокопий и генокопий. Фенокопии — изменения фенотипа под влиянием неблагоприятных факторов среды, по проявлению похожие на мутации. В медицине фенокопии — ненаследственные болезни, сходные с наследственными.Генокопии—это одинаковые изменения фенотипа, обусловленные аллелями разных генов.
Изменчивость
1. Ненаследственная, модификационная или фенотипическая;
2. Наследственная, мутационная или генотипическая;
3. Наследственная, комбинативная или генотипическая.