

1.Самореплицирующиеся молекулы. Естественный отбор самореплицирующихся молекул. Значение возникновения мембранных структур в эволюции клетки
2.Нуклеиновые кислоты. Строение азотистых оснований. Пуриновые и пиримидиновые основания. Строение нуклеозидов и нуклеотидов
3.Доказательства генетической роли нуклеиновых кислот. Явление трансформации у бактерий. Эксперименты Эйвери, Мак-Леонда и Мак-Карти.
4. Модель ДНК Уотсона и Крика
5. Вращение связей между атомами, формирующими сахарофосфатный остов ДНК и свободное вращение С-1/-N-гликозидной связи – основа структурных вариаций в ДНК. Cин- и анти-конформации.
6.Альтернативные двухспиральные структуры ДНК. Параметры B-, A- и Z-форм ДНК.
7. Положение атомов N7,O6,N6в пуринах – положение Хугстена. Хугстеновское спаривание – основа формирования триплексов ДНК(Н-ДНК).
. Явление суперспирализации ДНК. Образование супервитков в концевых молекулах ДНК – основа формирования третичной структуры.
9. Регуляция топологии ДНК in vivo. ДНК-топоизомеразы. Классификация ДНК-топоизомераз
10.Организация бактериального генома. Нуклеоид. Роль белков HU и H в компактизации ДНК E. coli и их краткая характеристика. Вспомогательная функция полиаминов, белков HLP1 и Р в конденсации ДНК бактерий.
11. Характеристика волокон хроматина диаметром 10 нм и 30 нм. Мономерная нуклеосома. Структурирующая роль гистонов в организации генома эукариот.
12. Классификация гистонов. Эволюционная стабильность гистонов Н3 и Н4. Роль оснóвных N-концевых «хвостов» гистонов в конденсации ДНК.
13. Первый уровень упоковки ДНК в хромосомею Минимальная нуклеосома( нуклеосомный кор). Нуклеосома. Гистон-ацетилтрансфераза НАТ.
14. Значение ацетилирования в регуляции процессов сборки нуклеосом de novo. Сайты модификации N-концевых последовательностей гистонов Н3 и Н4 с участием цитоплазматической гистон-ацетилтрансферазы HAT1
15. Доказательство полуконсервативного способа репликации ДНК. Эксперимент Месельсона и Сталя. Репликативная вилка. Одно- и двунаправленная репликация.
16. Типы репликации. Репликация кольцевых ДНК с образованием глазка. -структура. Репликация по типу катящегося кольца (репликация ДНК фагов М13, Х174, ). Репликация с образованием D-петель.
17. ДНК-полимеразы E. coli (I, II, III, IV и V). Сравнительная характеристика ДНК-полимераз. ДНК-полимераза III – основной фермент репликации. Понятие процессивности.
18.Локус oriC – точка начала репликации. Структура локуса oriC. Предзатравочный комплекс. Роль белков dnaА, HU, dnaС и dnaВ в формировании предзатравочного комплекса.
19. Регуляция инициации репликации ДНК E. coli. Dam-метилаза. Последовательности GATC. Терминация репликации.
20. Множественность точек начала репликации у эукариот. «Лицензирование» как механизм контроля согласованной активация точек начала репликации.
21.Точки начала репликации Saccharomyces cerevisiae. АRS-элементы. Консервативные последовательности в составе ARS-элементов.
22. ARS-кор, как основная (базовая) последовательность ARS-элементов. Белки ORC – основа формирования предрепликационных комплексов.
23. Сборка предрепликационного комплекса. Комплекс ДНК-полимераза /праймаза. Инициаторная ДНК. Процессивный синтез ДНК с участием ДНК-полимеразы .
24. Системы рестрикции и модификации. Уникальный тип метилирования ДНК прокариот. Представление о метилированных, полуметилированных и неметилированных сайтах. Явление рестрикции ДНК.
25. Рестриктазы типа 2(класс 1). Характеристика рестриктазы Eco R1. Природа сайтов узнавания и рестрикции. «Липкие» и «тупые» концы.
26. Молекулярная основа мутаций. Точечные мутации. Транзиции и трансверсии. Горячие точки. Молчащие мутации. Нейтральные мутации.
26. Молекулярная основа мутаций. Точечные мутации. Транзиции и трансверсии. Горячие точки. Молчащие мутации. Нейтральные мутации.
27. Причины мутаций. Таутомерные формы оснований. Дезаминирование оснований. Апуринизация ДНК. Алкилирующие агенты.
28. Действие физических факторов (влияние на равновесие амино – имино-форм). Ошибки ДНК-полимеразы, связанные с включением аналогов природных нуклеотидов.
29. Система коррекции несоответствий спаривания оснований. Роль последовательностей GATC в mismatch репарации.
30. Эксцизионная репарация оснований. АР-сайты. ДНК-гликозилазы. Удаление урацила и тимина. Субстратная специфичность ДНК-гликозилаз UNG, hGMUG1, TDG и MBD4 человека.
31. Эксцизионная репарация нуклеотидов. Удаление пиримидиновых димеров. Нуклеаза АВС (АВС-эксцинуклеаза). Компоненты АВС-эксцинуклеазы – белки UvrА, UvrB, UvrC. UvrD-геликаза.
32. Прямая репарация (обращение повреждений) ДНК. Механизмы обращения повреждений. О6-метилгуанин-ДНК-метилтрасфераза. Прямая репарация 1-метиладенина и 3-метилцитозина. Белок AlkB – a-кетоглутарат-Fe2+-зависимая диоксигеназа.
33. Репарация, включающая рекомбинацию. SOS-ответ. Понятие SOS-генов. Ферменты и белки SOS-репарации. Белки RecA и LexA.
34. Концепция информационной РНК. Концепция РНК-посредника Ф. Жакоба и Ж. Моно. иРНК – наиболее короткоживущая форма рибонуклеиновых кислот. Функциональное и химическое время полужизни информационных РНК
35.Строение информационных РНК. Строение информационных РНК прокариот. Лидерные, трейлерные и кодирующие участки. Полицистронность иРНК прокариот.
36. Строение иРНК эукариот. Структурные элементы иРНК эукариот. Полиаденилирование и кэпирование иРНК. Структура кэпов. Метилирование кэпов.
37.Специфические участки взаимодействия РНК-полимераз с ДНК. Структура промоторов. Стартовая точка, положения по ходу транскрипции и положения против хода транскрипции. Блок Прибнова и –35-блок.
38. Классификация РНК-полимераз эукариот
39. Промоторы эукариот. Блок Хогнесса. –70-блок.
40.Закрытый и открытый двойные комплексы. Тройной комплекс. Роль σ-фактора.
41. Терминация транскрипции. ρ-зависимые и ρ-независимые терминаторы.
42. Прерывистость генов Э. Процессинг и сплайсинг транскриптов РНК-полимеразы II.
43. Интроны, подчиняющиеся «правилу GU/AG». Донор сплайсинга и акцептор сплайсинга. Точка ветвления интрона. Малые ядерные РНК (мяРНК) и сплайсисома
44.Процессинг транскриптов, синтезируемых РНК-полимеразой III.
45. Механизм сплайсинга тРНК. тРНК-лигаза.
46. Процессинг транскриптов РНК-полимеразы I. Наружные (НТС) и внутренние (ВТС) транскрибируемые спейсеры. Специфические малые ядрышковые РНК C/D и Н/АСА в реакциях 2′-О-метилирования и псевдоуридилирования рРНК
47. Представления об интронах группы I как о рибозимах. Самосплайсинг рРНК Tetrahymena thermophila.
48. Интроны группы II – второй класс самосплайсирующихся интронов. Явление РНК-интерференции и редактирование РНК.
49. Экспериментальная расшифровка генетического кода. Синтетические олигонуклеотиды. Метод связывания рибосом.
50. Периодические сополимеры Кораны. Установление значения терминирующих кодонов.
51. Строение тРНК. Пространственная L-форма тРНК. Минорные компоненты тРНК.
52. Гипотеза «качаний».
53. Активация аминокислот. Активные центры в аминоацил-тРНК-синтетазах. Аминоацил-тРНК-синтетазы – самокорректирующие ферменты.
54.Стадии активации АК.
55. Инициация трансляции. Инициаторные тРНК прокариот. Факторы инициации прокариот.
56. Элонгация. Факторы элонгации. Образование пептидной связи.
57. Терминация синтеза полипептидных цепей. Терминирующие кодоны. RF-1 и RF-2 – белковые факторы терминации. Фактор терминации RF-3
58. Перепрограммирование трансляции. Включение Se-цистеина как один из механизмов перекодирования трансляции. Уникальность структуры тРНКSeс П. и Э.
59. Явление транс-трансляции. Бактериальная тм-РНК. Совмещение свойств транспортной и матричной РНК в тмРНК.
60. Методы промышленной микробиологии. Суперпродукция лизина.
61. Представления о принципах генетической рекомбинации с использованием подвижных генетических элементов.
62. Способы соединения фрагментов ДНК invitro. Метод линкеров. Коннекторный метод.
63.Понятие вектора. Векторы в генетической инженерии. Свойства «идеального» вектора.
64. Плазмиды, классификация плазмид. Структура сайтов рестрикции эндонуклеаз типа II. «Тупые» и «липкие» концы.
65. Трансформация, трансфекция, клонирование.
66. Способы получения ДНК для клонирования
1.Самореплицирующиеся молекулы. Естественный отбор самореплицирующихся молекул. Значение возникновения мембранных структур в эволюции клетки.
Пространственная трехмерная укладка полинуклеотида не может не влиять на его стабильность и на способность реплицироваться, поэтому не все молекулы в реплицирующейся смеси одинаково успешно смогут размножаться. Реплицирующиеся молекулы РНК подвержены своеобразному естественному отбору, в результате которого в зависимости от создаваемых условий начинает преобладать та или иная последовательность. Оба свойства молекулы РНК - информационное и функциональное - являются необходимыми предпосылками эволюционного процесса. Они могут катализировать следующие процессы:
-образование и разрушение ковалентных связей, в том числе и между нуклеотидами;
-некоторые специализированные молекулы РНК могут катализировать изменения в других молекулах РНК, разрезая нуклеотидную последовательность в определенной точке
-другие типы РНК могут вырезать часть своей собственной последовательности и сшивать разрезанные концы
Важнейшая роль в эволюции клеточных мембран принадлежит классу амфипатических соединений: одна часть молекулы гидрофобна, а другая – гидрофильна.
Генетические и каталитические свойства РНК позволяют предположить, что эти молекулы первыми включились в эволюцию. После возникновения достаточно эффективного механизма синтеза белка ДНК приняла на себя генетическую функцию, при этом белки стали катализаторами, а РНК сохранилась, главным образом, как промежуточное звено между ними. ДНК стала необходимой в качестве главной информационной молекулы только тогда, когда клетки значительно усложнились и для них потребовалось больше генетической информации, чем та, которую могли стабильно обеспечить молекулы РНК.
Дезоксирибонуклеиновая кислота используется для хранения генетической информации по ряду причин:
- молекула ДНК химически более стабильна, чем молекула РНК;
- в молекуле ДНК отсутствует одна гидроксильная группа, что делает ее более устойчивой к гидролизу, чем РНК;
- ДНК существует в виде двухцепочечной молекулы, состоящей из двух комплементарных полинуклеотидных цепей, что позволяет ДНК легко реплицироваться и репарировать возникающие в ней повреждения, при этом неповрежденная цепь используется в качестве матрицы для восстановления комплементарной дефектной цепи;
- используя принцип комплементарности, ДНК направляет синтез отдельных видов молекул РНК. Синтезированные таким способом одноцепочечные молекулы РНК выполняют две другие функции первобытных полинуклеотидов: одни направляют синтез белков, как кодирующие молекулы (информационные РНК) , а другие, как каталитические молекулы (рибосомные и другие неинформационные РНК).
2.Нуклеиновые кислоты. Строение азотистых оснований. Пуриновые и пиримидиновые основания. Строение нуклеозидов и нуклеотидов
Нуклеиновые кислоты обеспечивают хранение и передачу наследственной информации, принимают участие в механизмах ее реализации. Структурные компоненты нуклеиновых кислот выполняют функции кофакторов, аллостерических эффекторов, входят в состав коферментов, принимая непосредственное участие в обмене веществ, а также в накоплении, переносе и трасформации энергии.
Нуклеиновые кислоты (ДНК и РНК) – сложные высокомолекулярные соединения, которые обеспечивают хранение и передачу наследственной информации; полимеры, мономерами которых служат нуклеотиды. Каждый нуклеотид (рибонуклеотид или дезоксирибонуклеотид) состоит из трех частей:
1) остатка фосфорной кислоты,
2) пятиуглеродного моносахарида в циклической форме,
3) азотистого основания.
В молекуле ДНК углевод представлен дезоксирибозой, а в молекуле РНК – рибозой.
Существует 2 типа азотистых оснований – пуриновые и пиримидиновые. Основу их составляют пурин и пиримидин соответственно. Пуриновые основания: аденин, гуанин. Пиримидиновые основания: тимин, цитозин, урацил. В ДНК содержится тимин, а в РНК – урацил.
Пуриновое или пиримидиновое основание в нуклеотидах соединяется с углеводом N-гликозидной связью. В образовании N-гликозидной связи в пуриновых нуклеотидах принимают участие N-9 пурина и С-1' пентозы, а в пиримидиновых нуклеотидах – N-1 пиримидина и С-1' пентозы.
Нуклеозид состоит из пятиуглеродного моносахарида в циклической форме и азотистого основания (аденозин, гуанозин, уридин, цитидин, тимидин).
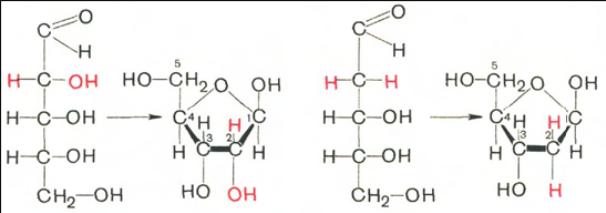
D-рибоза D-рибофураноза D-2-дезоксирибоза D-2-дезоксирибофураноза
D-рибоза D-рибофураноза D-2-дезоксирибоза D-2-дезоксирибофураноза
Основу структуры пуриновых и пиримидиновых оснований составляют два ароматических гетероциклических соединения – пиримидин и пурин:
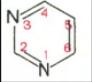
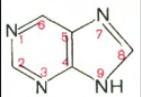
Пиримидин Пурин
Молекула пурина состоит из двух конденсированных колец: пиримидина и имидазола.
В составе нуклеиновых кислот встречаются три главных пиримидиновых основания: цитозин, урацил и тимин.
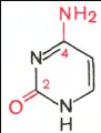
![]()
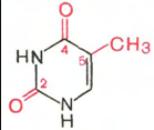
Цитозин Урацил Тимин
Два пуриновых основания, постоянно встречающихся в гидролизатах нуклеиновых кислот, имеют следующее строение:
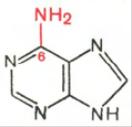
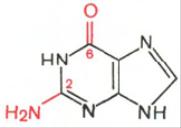
Аденин Гуанин