

Потенциал действия, механизм его возникновения. Деполяризация и реполяризация мембраны клетки как результат изменения ионной проницаемости.
Потенциал действия (ПД) - это кратковременные высоко амплитуды и изменения МПС, возникающих при возбуждении. Основной причиной ПД является изменение проницаемости мембраны для ионов. Рассмотрим развитие ПД на примере нервного волокна. Регистрировать ПД можно при введении одного из электродов в волокно или располагая оба электрода на его поверхности. Проследим процесс формирования ПД при внутриклеточном методе. 1. В состоянии покоя мембрана поляризована и МПС равен 90 мВ. 2. Как только начинается возбуждение, величина этого потенциала уменьшается (это уменьшение называется деполяризацией). В ряде случаев потенциал сторон мембраны меняется на противоположный (так называемый овершут). Это первая стадия ПД - деполяризация. 3. Стадия реполяризации, при которой величина разности потенциалов падает почти до первоначального уровня. Эти две фазы в пиком ПД. 4. После пика наблюдаются следовые потенциалы - следовая деполяризация и следовая гиперполяризация (гиперполяризация - увеличение разности потенциалов между сторонами мембраны). Например, было 90 мВ, а становится 100 мВ. Развивается ПД очень быстро - за несколько миллисекунд. Параметры ПД: 1) переменный характер, поскольку меняется направление движения тока, 2) величина, которая благодаря овершута может превышать МПС; 3) время, в течение которого развиваются ПД и отдельные его стадии - деполяризация, реполяризация, следовая гиперполяризация. Как формируется ПД. В состоянии покоя «ворота» потенциалзависимых Na +-каналов закрыты. Закрытые также «ворота» потенциалзависимых К +-каналов. 1. Во время фазы деполяризации происходит активизация Na +-Kaналив. При этом изменяется конформационный состояние белков, входящих в состав «ворот». Эти «ворота» открываются, и проницаемость мембраны для Na + увеличивается в несколько тысяч раз. Na + лавообразные входит в волокно нерва. В настоящее время К +-каналы открываются очень медленно. Так, в волокно поступает значительно больше Na +, чем выводится из него К +. 2. Реполяризация характеризуется закрытием Na +-каналов. «Ворота» на внутренней поверхности мембраны закрываются - наблюдается инактивация каналов под воздействием электрических потенциалов. Инактивация происходит медленнее, чем активация. В настоящее время ускоряется активация К +-каналов и растет диффузия К + наружу. Таким образом, деполяризация связана преимущественно со входом Na + в волокно, а реполяризация - с выходом из него К +. Соотношение между входом Na + и выходом К + изменяется в процессе раз «витке ПД: в начале ПД входит Na + в несколько тысяч раз больше, чем получается К +, а затем выходит более К +, чем входит Na +. Причиной следовых потенциалов дальнейшие изменения соотношения между этими двумя процессами. Во время следовой гиперполяризации много К +-каналов еще остаются открытыми и К + продолжает выходить наружу. Восстановление ионных градиентов после ПД. Одиночные ПД изменяют разность концентраций ионов в нервном волокне и за его пределами очень мало. Но в тех случаях, когда проходит значительное количество импульсов, эта разница может быть весьма существенной. Восстановление ионных градиентов происходит тогда за счет усиления работы Na + / K +-HacociB - в большей степени нарушается этот градиент, тем интенсивнее работают насосы. При этом используется энергия АТФ. Часть ее выделяется в виде тепла, поэтому в этих случаях наблюдается кратковременное повышение температуры волокна. Условия необходимые для возникновения ПД. ПД возникает лишь при определенных условиях. Раздражители, действующие на волокну, могут быть разными. Чаще используется постоянный электрический ток. Он легко дозируется, мало травмирует ткань и ближайший тех раздражителей, которые существуют в живых организмах. При каких условиях постоянный ток может зумовитК появление ПД? Ток должен быть достаточно сильным, действовать определенное время, его нарастание должно быть быстрым. Наконец, имеет значение и направление тока (действие анода или катода). В зависимости от силы различают подпороговый (недостаточный для возникновения возбуждения), пороговый (достаточный) и надпороговый (чрезмерный) ток. Несмотря на то что подпороговый ток не вызывает возбуждение, он все же деполяризует мембрану, и эта деполяризация тем больше, чем выше его напряжение. Деполяризация, развивающийся при этом, называется локальным ответом и является видом местного возбуждения. Оно характеризуется тем, что не распространяется, величина его зависит от силы раздражения (закрном силовых отношений: чем больше сила раздражения, тем активнее ответ). При локальной ответа возбудимость ткани повышается. Возбудимость - это способность отвечать на раздражение и переходить в состояние возбуждения. Если сила раздражителя достаточное (пороговая), то деполяризация достигает определенной величины, называется критическим уровнем деполяризации (Ек). Для нервного волокна, покрытого миелином, Ек составляет около 65 мВ. Таким образом, разница между МПС (Е0), равный в данном случае 90 мВ, и Ек составляет 25 мВ. Эта величина (ДЕ = Е0-Ек) очень важна для характеристики возбудимости ткани. Когда Е0 увеличивается при деполяризации, возбудимость выше и, наоборот, снижение Е0 при гиперполяризации приводит к ее уменьшению. ГДЕ может зависеть не только от величины Е0, но и от критического уровня деполяризации (Ек). При пороговой силе раздражителя возникает ПД. Это уже не местное возбуждение, оно способно распространяться на дальние расстояния, подлежит закону «все или ничего» (при увеличении силы раздражителя амплитуда ПД не растет). Возбудимость при развитии ПД отсутствует или значительно снижена. ПД является одним из показателей возбуждения - активного физиологического процесса, которым живые клетки (нервные, мышечные, железистые) отвечают на раздражение. Во время возбуждения меняются метаболизм, температура клеток, нарушается ионная равновесие между цитоплазмой и внешней средой, происходит ряд других процессов. Кроме силы постоянного тока, возникновения ПД зависит также от длительности его действия. Существует обратная пропорциональная зависимость между силой тока и длительностью его действия. Подпороговый ток даже при очень длительном воздействии не приведет возбуждения. Надпороговый ток при слишком короткой действия также не приведет к возбуждению. Для возникновения возбуждения требуется также определенная скорость (крутизна) нарастание силы тока. Если повышать силу тока очень медленно, то при этом будет меняться Ек и Е0 может не достичь его уровня. Имеет значение и направление тока: ПД возникает при замыкании тока только тогда, когда катод помещается на внешней поверхности мембраны, а анод - в клетке или волокне. При прохождении тока изменяется МП. Если на поверхности лежит катод, то развивается деполяризация (возбудимость повышается), а если анод - гиперполяризация (возбудимость снижается). Знание механизмов действия электрического тока на живые объекты крайне необходимо для разработки и применения в клинике методов физиотерапии (диатермия, УВЧ, гипергидроз и др.).. Изменение возбудимости при ПД. При локальной ответа возбудимость повышается (ДЭ уменьшается). Изменения возбудимости во время самого ПД можно заметить, если раздражать повторно в разные стадии развития ПД. Оказывается, что во время пика даже очень сильное повторное раздражение остается без ответа (период абсолютной рефрактерности). Затем возбудимость постепенно нормализуется, но она все же ниже, чем начальная (период относительной рефрактерности). При выраженной следовой деполяризации возбудимость выше, чем первоначальная, а при положительной следового потенциала возбудимость снова снижается. Абсолютная рефрактерность объясняется инактивизациею Na + каналов и повышением проводимости К + - каналов. При относительной рефрактерности вновь активизируются Na + - каналы и снижается правиднисть К + - каналов. Двухфазный характер ПД. Обычно в условиях, когда микроэлектрод содержится внутри клетки или волокна, наблюдается однофазный ПД. Иная картина бывает в тех случаях, когда оба электрода лежат на внешней поверхности мембраны - биполярная регистрация. Возбуждение, которое представляет собой волну электроотрицательности, перемещаясь по мембране, доходит сначала до одного электрода, затем помещается между электродами, наконец достигает второго электрода, а затем распространяется дальше. В этих условиях ПД имеет двухфазный характер. Регистрация ПД широко используется в клинике для диагностики заболеваний сердца, мозга, опорно-двигательного аппарата, желудка и др. Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации, начальной быстрой реполяризации, переходящей в фазу медленной реполяризации (фаза плато) и фазы быстрой конечной реполяризации (рис. 1). Фаза быстрой деполяризации создается резким повышением проницаемости мембраны для ионов натрия, что приводит к возникновению быстрого входящего натриевого тока. Последний, однако, при достижении мембранного потенциала 30–40 мВ, инактивируется и в последующем, вплоть до инверсии потенциала (около +30 мВ) и в фазу «плато», ведущее значение имеют кальциевые ионные токи. Деполяризация мембраны вызывает активацию кальциевых каналов, в результате чего возникает дополнительный деполяризующий входящий кальциевый ток.
Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя. Длительность потенциала действия кардиомиоцитов составляет 300–400 мс, что соответствует длительности сокращения миокарда (рис. 2).
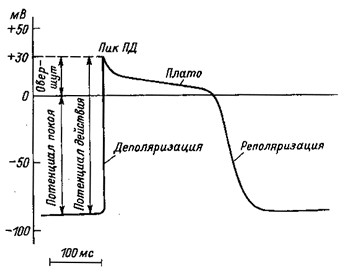
Рис. 1. Потенциал действия клетки рабочего миокарда. Быстрое развитие деполяризации и продолжительная реполяризация. Замедленная реполяризация (плато) переходит в быструю реполяризацию
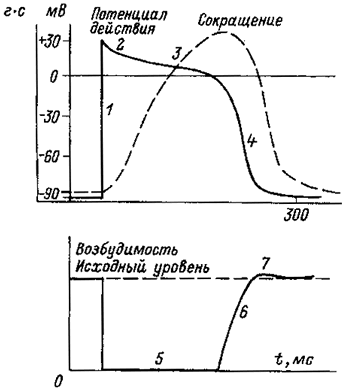
Рис. 2. Сопоставление потенциала действия и сокращение миокарда с фазами изменения возбудимости при возбуждении
1 – фаза деполяризации; 2 – фаза начальной быстрой реполяризации; 3 – фаза медленной реполяризации (фаза плато); 4 – фаза конечной быстрой реполяризации; 5 – фаза абсолютной рефрактерности; 6 – фаза относительной рефрактерности; 7 – фаза супернормальной возбудимости. Рефрактерность миокарда практически совпадает не только с возбуждением, но и с периодом сокращении
Пищеварение в полости рта. Состав и свойства слюны, реакция слюны на действие различных раздражителей. Регуляция слюноотделения
Пищеварение в полости рта
Пищеварение начинается в ротовой полости, где происходит механическая и химическая обработка пищи. Механическая обработка заключается в измельчении пищи, смачивании ее слюной и формировании пищевого комка. Химическая обработка происходит за счет ферментов, содержащихся в слюне.
В полость рта впадают протоки трех пар крупных слюнных желез: околоушных, подчелюстных, подъязычных и множества мелких желез, находящихся на поверхности языка и в слизистой оболочке нёба и щек.
Околоушные железы и железы, расположенные на боковых поверхностях языка, — серозные (белковые). Их секрет содержит много воды, белка и солей. Железы, расположенные на корне языка, твердом и мягком нёбе, относятся к слизистым слюнным железам, секрет которых содержит много муцина. Подчелюстные и подъязычные железы являются смешанными.
Состав и свойства слюны
Слюна, находящаяся в ротовой полости, является смешанной. Ее рН равна 6,8-7,4. У взрослого человека за сутки образуется 0,5-2 л слюны. Она состоит из 99% воды и 1% сухого остатка. Сухой остаток представлен органическими и неорганическими веществами. Среди неорганических веществ — анионы хлоридов, бикарбонатов, сульфатов, фосфатов; катионы натрия, калия, кальция магния, а также микроэлементы: железо, медь, никель и др. Органические вещества слюны представлены в основном белками. Белковое слизистое вещество муцин склеивает отдельные частицы пищи и формирует пищевой комок.
Основными ферментами слюны являются амилаза и мальтаза, которые действуют только в слабощелочной среде. Амилаза расщепляет полисахариды (крахмал, гликоген) до мальтозы (дисахарида). Мальтаза действует на мальтозу и расщепляет ее до глюкозы. В слюне в небольших количествах обнаружены также и другие ферменты: гидролазы, оксиредуктазы, трансферазы, протеазы, пептидазы, кислая и щелочная фосфатазы. В слюне содержится белковое вещество лизоцим (мурамидаза), обладающее бактерицидным действием. Пища находится в полости рта всего около 15 секунд, поэтому здесь не происходит полного расщепления крахмала. Но пищеварение в ротовой полости имеет очень большое значение, так как является пусковым механизмом для функционирования желудочно-кишечного тракта и дальнейшего расщепления пищи.
Функции слюны
Пищеварительная функция — о ней было сказано выше.
Экскреторная функция. В составе слюны могут выделяться некоторые продукты обмена, такие как мочевина, мочевая кислота, лекарственные вещества (хинин, стрихнин), а также вещества, поступившие в организм (соли ртути, свинца, алкоголь).
Защитная функция. Слюна обладает бактерицидным действием благодаря содержанию лизоцима. Муцин способен нейтрализовать кислоты и щелочи. В слюне находится большое количество иммуноглобулинов, что защищает организм от патогенной микрофлоры. В слюне обнаружены вещества, относящиеся к системе свертывания крови: факторы свертывания крови, обеспечивающие местный гемостаз; вещества, препятствующие свертыванию крови и обладающие фибринолитической активностью; вещество, стабилизирующее фибрин. Слюна защищает слизистую оболочку полости рта от пересыхания.
Трофическая функция. Слюна является источником кальция, фосфора, цинка для формирования эмали зуба.
Регуляция слюноотделения
При поступлении пищи в ротовую полость происходит раздражение механо-, термо- и хеморецепторов слизистой оболочки. Возбуждение от этих рецепторов по чувствительным волокнам язычного (ветвь тройничного нерва) и языкоглоточного нервов, барабанной струны (ветвь лицевого нерва) и верхнегортанного нерва (ветвь блуждающего нерва) поступает в центр слюноотделения в продолговатом мозге. От слюноотделительного центра по эфферентным волокнам возбуждение доходит до слюнных желез и железы начинают выделять слюну. Эфферентный путь представлен парасимпатическими и симпатическими волокнами. Парасимпатическая иннервация слюнных желез осуществляется волокнами языкоглоточного нерва и барабанной струны, симпатическая иннервация — волокнами, отходящими от верхнего шейного симпатического узла. Тела преганглионарных нейронов находятся в боковых рогах спинного мозга на уровне II-IV грудных сегментов. Ацетилхолин, выделяющийся при раздражении парасимпатических волокон, иннервирующих слюнные железы, приводит к отделению большого количества жидкой слюны, которая содержит много солей и мало органических веществ. Норадреналин, выделяющийся при раздражении симпатических волокон, вызывает отделение небольшого количества густой, вязкой слюны, которая содержит мало солей и много органических веществ. Такое же действие оказывает адреналин. Субстанция Р стимулирует секрецию слюны. СО2 усиливает слюнообразование. Болевые раздражения, отрицательные эмоции, умственное напряжение тормозят секрецию слюны.
Слюноотделение осуществляется не только с помощью безусловных, но и условных рефлексов. Вид и запах пищи, звуки, связанные с приготовлением пищи, а также другие раздражители, если они раньше совпадали с приемом пищи, разговор и воспоминание о пище вызывают условно-рефлекторное слюноотделение.