

ГЛАВА 4. Гуморальные механизмы адаптивного иммунитета
Происхождение и дифференцировка В-лимфоцитов. Субпопуляции специализированных В-лимфоцитов. Основные рецепторы и маркеры В-лимфоцитов. Антитела как главные эффекторные молекулы гуморального звена адаптивного иммунитета. Химическое строение иммуноглобулинов. Генетический контроль синтеза иммуноглобулинов. Изотипы, аллотипы и идиотипы иммуноглобулинов. Антиидиотипы. Антиидиотипическая сеть. Особенности строения и функции отдельных классов иммуноглобулинов. Моноклональные антитела.
Гуморальный адаптивный иммунитет – это специализированное звено системы адаптивного иммунитета, главной функцией которого является синтез антител против антигенов. Функциональная активность этой системы в большинстве случаев зависит от тесного взаимодействия с Т-системой адаптивного иммунитета и антигенпредставляющими клетками.
Главными клетками системы гуморального адаптивного иммунитета являются В-лимфоциты, осуществляющие синтез антител. Участие антител в реализации гуморального адаптивного иммунного ответа связано с образованием иммунных комплексов с растворимыми и корпускулярными антигенами. В составе иммунных комплексов антигены подвергаются изоляции и/или уничтожению тремя основными способами: нейтрализацией, активацией системы комплемента и фагоцитозом.
Таким образом, именно антитела являются основным эффекторным звеном системы гуморального адаптивного иммунитета. При этом главной функцией антител является специфическое связывание антигена. Специфическое взаимодействие антигенов и антител основано на пространственном соответствии, или комплементарности, существующем между ними.
Центральный орган системы гуморального адаптивного иммунитета впервые был обнаружен у птиц, им оказалась Фабрициева сумка (Bursa Fabricius), отсюда лимфоциты, происходящие из этого образования, получили название В-лимфоцитов, а вся система гуморального адаптивного иммунитета – В-системы.
У млекопитающих аналогом Bursa Fabricius в эмбриональном периоде являются кроветворные очаги в печени, а в постнатальном периоде – костный мозг (Воne marrow).
Периферическими органами В-системы адаптивного иммунитета являются тимуснезависимые зоны селезенки, лимфоузлов и других лимфоидных образований (миндалин, аденоидов, пейеровых бляшек кишечника, аппендикса). Это такие зоны, в которых преимущественно локализованы В-лимфоциты, в отличие от тимусзависимых зон, заселенных главным образом Т-лимфоцитами, мигрировавшими в них через кровоток из тимуса.
В периферической крови содержится в среднем 10-20% В-лимфоцитов,
их абсолютное содержание составляет 0,19-0,38х109/л. В основном, это незрелые клетки, прошедшие первые стадии дифференцировки в костном мозге и мигрирующие в периферические лимфоидные органы. Здесь в результате контактов с антигенами происходит их размножение и окончательная дифференцировка, образуются клоны плазматических клеток, основной функцией которых является синтез антител против антигена, вызвавшего их образование. Часть отвечающих на антиген В-лимфоцитов превращается в В-клетки памяти.
Каждый организм обладает громадным набором клонов В-лимфоцитов, несущих иммуноглобулиновые рецепторы самой разнообразной специфичности, что дает возможность синтезировать антитела практически к любым антигенам. Количество теоретически возможных конфигураций антигенных детерминант и соответственно клонов В-клеток оценивается величиной порядка 107-108. Вопрос о происхождении такого разнообразия антител – один из самых трудных в иммунологии. В настоящее время общепризнанным считается положение о генетической запрограммированности многообразия антител в каждом индивидуальном организме за счет набора большого количества генов, кодирующих вариабельные участки тяжелых и легких цепей иммуноглобулинов, и их комбинаций.
Роль в-лимфоцитов в презентации антигена
Кроме своей основной функции – синтеза антител – В-лимфоциты могут выполнять и функцию антигенпредставляющих клеток. Ее осуществют находящиеся в лимфоузлах и других периферических лимфоидных органах так называемые фолликулярные В-лимфоциты. Эти клетки могут поглощать антигены путем неспецифического фагоцитоза, однако этот процесс в 10000 раз менее эффективен, чем поглощение В-лимфоцитами антигенов, связанных с рецепторными мембранными иммуноглобулинами (ВКР). После поглощения (интернализации) комплекса «ВКР-антиген» начинается его процессинг в фаголизосомах клетки и образование иммуногенных комплексов антигенных пептидов с молекулами МНС II класса, которые презентируются на поверхности В-лимфоцита для распознавания Т-хелперам. Параллельно на поверхности В-лимфоцита начинается экспрессия костимулирующих молекул В7, обеспечивающих путем взаимодействия В7/CD28 полноценную активацию Т-хелперов, распознающих антиген на поверхности В-лимфоцитов, а также стимулирующих продукцию ими цитокинов. Процессинг и презентация антигенов В-клетками стимулируется IFN-γ.
В-лимфоциты могут выполнять роль основных антигенпредставляющих клеток при низкой концентрации антигена, а также при вторичном адаптивном иммунном ответе.
Происхождение и дифференцировка в-лимфоцитов
Весь путь развития В-лимфоцитов от полипотентной стволовой кроветворной клетки до образования синтезирующих антитела плазматических клеток и В-клеток памяти включает антигеннезависимую и антигензависимую стадии дифференцировки. Антигеннезависимая дифференцировка В-лимфоцитов происходит в костном мозге, в ее основе лежит перестройка генов иммуноглобулинов, приводящая к формированию клонов В-лимфоцитов, экспрессирующих на своей поверхности рецепторные иммуноглобулины различной специфичности. В процессе антигензависимой дифференцировки в периферических лимфоидных органах происходит активация В-лимфоцитов в результате распознавания соответствующих антигенов с помощью иммуноглобулиновых антигенраспознающих рецепторов (ВКР); их пролиферация и дифференцировка в плазматические клетки, синтезирующие антитела, и В-клетки памяти. На этой стадии происходит также реарранжировка генов иммуноглобулинов, приводящая к синтезу разных изотипов иммуноглобулинов.
Образование В-лимфоцитов начинается в эмбриогенезе и продолжается в течение всей жизни. В эмбриональном периоде В-лимфоциты образуются в желточном мешке, эмбриональной печени и в эмбриональном костном мозге. После рождения единственным источником В-лимфоцитов у млекопитающих и человека является костный мозг.
Основные этапы дифференцировки в-лимфоцитов
Схема, отражающая основные этапы дифференцировки В-лимфоцитов, представлена на рис.19.
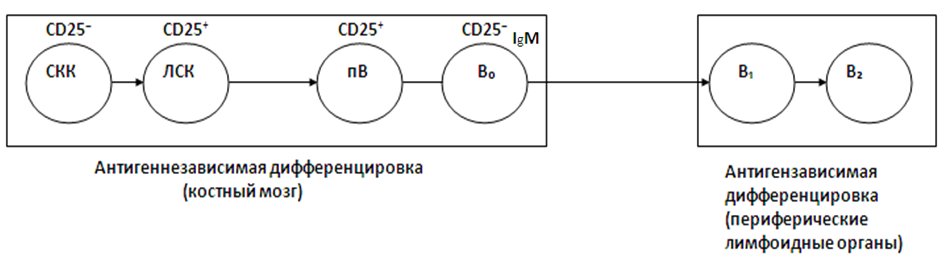
Рисунок 19. Основные этапы дифференцировки В-лимфоцитов
СКК – стволовая кроветворная клетка костного мозга;
ЛСК – лимфоидная стволовая клетка (общий предшественник лимфопоэза);
пВ – предшественник В-лимфоцитов;
В0–лимфоциты;
В1 – лимфоциты (наивные В-лимфоциты);
В2 – лимфоциты;
Родоначальником В-лимфоцитов, как и других клеток крови, является полипотентная стволовая кроветворная клетка костного мозга (СКК). Через стадию лимфоидной стволовой клетки (ЛСК) – общего предшественника всех (Т- и В-) лимфоцитов – происходит формирование предшественников В-лимфоцитов (пВ), которые затем превращаются в незрелые В0-лимфоциты. Эта дифференцировка происходит в костном мозге без взаимодействия с антигенами.
Предшественники В-лимфоцитов образуются из лимфоидных стволовых клеток под влиянием микроокружения костномозговых стромальных клеток. Стромальные клетки поддерживают процесс развития пВ-клеток путем прямых межклеточных взаимодействий с помощью различных молекул клеточной адгезии (VLA-4, VCAM-1 и др.), а также с помощью поверхностных молекул стромальных клеток, таких как SCF (фактор стволовых клеток). Стромальные клетки костного мозга секретируют также целый ряд цитокинов: ИЛ-3,4,6, а также ИЛ-7, который поддерживает процесс развития пВ-клеток на ранних этапах их развития.
В0-лимфоциты мигрируют затем через кровоток и заселяют тимуснезависимые зоны периферических лимфоидных органов. Там происходит дальнейшее созревание и дифференцировка В-лимфоцитов до стадии В1 и В2 и приобретение ими способности к синтезу всех классов иммуноглобулинов в ответ на поступающие в организм антигены.
В0-лимфоциты – это популяция иммунологически незрелых В-лимфоцитов. На их поверхности имеются только антигенраспознающие рецепторы, относящиеся к классу IgМ. Однако плотность этих рецепторов на единицу поверхности клеток очень низка, поэтому они не могут обеспечить эффективного взаимодействия с антигеном и не синтезируют IgМ в периферическую кровь. Стадия В0 соответствует моменту формирования клонов В-лимфоцитов. Клон В-лимфоцитов – это группа В-лимфоцитов, имеющих на своей поверхности иммуноглобулиновые рецепторы одной специфичности. На стадии В0 костномозговые В-лимфоциты проходят негативную селекцию, в результате которой аутореактивные клоны В-лимфоцитов погибают от апоптоза или подвергаются рецепторному редактированию, что лишает их иммуноглобулиновые рецепторы способности к взаимодействию с собственными антигенами. Эти процессы лежат в основе формирования центральной В-клеточной толерантности.
Незрелые В0-лимфоциты, которые покидают костный мозг и выходят на периферию, называются еще транзиторными В-лимфоцитами. При контакте с антигенами на периферии функционально незрелые транзиторные В-лимфоциты вступают в состояние анергии или погибают от апоптоза.
В1-лимфоциты – это наивные В-лимфоциты, которые еще не встречались с антигеном, они обладают довольно короткой продолжительностью жизни (от 5-6 дней до 3-4 месяцев). Если наивные В1-лимфоциты не встретятся с соответствующим им по специфичности антигеном, они погибают, так как не смогут стать резидентными, находящимися в лимфоидных фолликулах В2-лимфоцитами.
На поверхности В1-лимфоцитов экспрессируются одновременно IgМ и IgD одной и той же антигенной специфичности, но IgD-рецепторы на В1-лимфоцитах представлены в гораздо меньшей степени. Хотя IgD является характеристическим поверхностным маркером зрелых наивных В-лимфоцитов, его функция пока не выяснена, однако доказана необходимость наличия IgD для развития В-лимфоцитов и их ответа на большинство антигенов. Плотность рецепторного IgM на единицу поверхности В1-клеток значительно выше, чем на стадии В0.
При встрече с соответствующим им по специфичности антигеном наивные В1-лимфоциты распознают его с помощью ВКР, активируются и вступают в пролиферацию с последующей дифференцировкой в плазматические клетки, синтезирующие антитела. Однако В1-лимфоциты могут непосредственно реагировать преимущественно на тимуснезависимые антигены синтезом IgМ. Примированные антигеном В1-лимфоциты – это уже полностью зрелые В-клетки, но еще не перешедшие в стадию В2 и не ставшие резидентными фолликулярными В-лимфоцитами.
В2-лимфоциты (называемые еще фолликулярными В-лимфоцитами, или В-2-лимфоцитами) образуются из примированных В2-лимфоцитов и находятся главным образом в фолликулах лимфоузлов и других лимфоидных органов, где контактируют с антигеном, представленным на поверхности АПК. Затем они пролиферируют, образуя зародышевые центры фолликулов и дифференцируются в плазматические клетки, синтезирующие антитела. В2-лимфоциты – это большая по численности популяция, состоящая из зрелых иммунокомпетентных клеток. На поверхности В2-лимфоцитов в высокой концентрации экспрессируется иммуноглобулин D (дифференцировочный), а также содержатся антигенраспознающие рецепторы, относящиеся ко всем классам иммуноглобулинов. В2-лимфоциты способны реагировать на любые антигены (как тимуcнезависимые, так и тимусзависимые) и осуществлять синтез иммуноглобулинов всех классов, что составляет основу вторичного гуморального адаптивногоиммунного ответа. Они требуют для своей активации помощи со стороны Т-хелперов, взаимодействие с которыми осуществляется в лимфоидных фолликулах как при непосредственном контакте этих клеток, так и с помощью синтезируемых Т-хелперами цитокинов (ИЛ-2, IFN-γ, ИЛ-4, ИЛ-5 и др.).
Сигналы от Т-хелперов индуцируют в фолликулярных В2-лимфоцитах переключение изотипов иммуноглобулинов, что обеспечивает продукцию антител с оптимальными свойствами, необходимыми для борьбы с данным антигеном. В процессе развития иммунного ответа средняя аффинность продуцируемых антител возрастает (процесс, называемый аффинным созреванием). Это происходит потому, что в процессе пролиферации отвечающего на антиген клона В-лимфоцитов в этих клетках происходят соматические гипермутации, а затем селекция наиболее эффективно связывающих антиген вариантов (тех В-лимфоцитов, чьи ВКР обладают максимальной аффинностью к данному В-клеточному эпитопу).
В2-лимфоциты дифференцируются не только в плазматические клетки, синтезирующие иммуноглобулины, но из них образуются и В-клетки памяти, хранящие информацию об антигене. В-клетки памяти – это долгоживущие малые В-лимфоциты, образовавшиеся из зрелых В-клеток в результате стимуляции антигеном при участии Т-лимфоцитов. При повторном внедрении в организм этого антигена благодаря В-лимфоцитам памяти ускоряется распознавание антигена, пролиферация соответствующего клона В-лимфоцитов и осуществляется быстрый синтез большого количества специфических антител против него.
В-лимфоциты памяти в отсутствие антигена могут также превращаться в плазматические клетки и синтезировать антитела определенной специфичности, обеспечивая иммунитет. Так, на вирусы оспы, полиомиелита, кори такой процесс осуществляется практически всю жизнь, на столбнячный токсин – всего около 5 лет, на дизентерийную палочку – около 1 месяца. Проблема усиления иммуногенности и увеличения сроков иммунологической памяти является весьма важной для создания эффективных вакцин.
На стадии В2 в результате антигензависимой дифференцировки внутри каждого клона В-лимфоцитов, реагирующего на данный антиген, поочередно, в результате переключения генов, кодирующих синтез тяжелых цепей молекул иммуноглобулинов, образуются четыре основные группы В2-лимфоцитов: BBBB которые осуществляют соответственно синтез и секрецию IgM, IgG, IgE и IgA. Этот процесс схематически изображен на рис. 20.
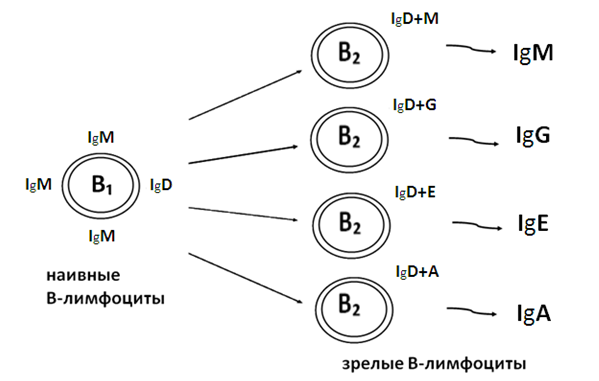
Рисунок 20. Дифференцировка В2-лимфоцитов
Первой в процессе дифференцировки образуется группа В2-лимфоцитов, на поверхности которых имеются IgD и IgM, эти клетки будут осуществлять синтез на экспорт IgM. Затем, в результате переключения генов в процессе дифференцировки, образуется группа В2-лимфоцитов, содержащих на поверхности IgD и IgG, которые синтезируют IgG против данного антигена; третьей образуется группа В2-лимфоцитов, имеющих на поверхности IgD и IgE и синтезирующих IgE; и, наконец, четвертой - В2-лимфоциты, содержащие на поверхности IgD и IgA, они дифференцируются в плазматические клетки, синтезирующие IgA. IgD, содержащийся на поверхности В2-лимфоцитов, имеет только мембраносвязанную форму и на экспорт в норме практически не синтезируется, поэтому в плазме крови он содержится в следовых количествах. Механизм действия этого иммуноглобулина пока еще мало изучен, однако экспериментально показано, что его удаление с поверхности В-лимфоцитов приводит к утрате ими способности к реакции на тимусзависимые антигены.
Таким образом, при поступлении в организм любого антигена (тимуснезависимого или тимусзависимого) первым против данного антигена в процессе гуморального иммунного ответа синтезируется IgM, а затем идет синтез всех остальных классов иммуноглобулинов - IgG, IgE и IgA, что обеспечивает эффективную защиту всех органов и тканей организма.