

Лекція № 4 Тема. Філогенія тваринного світу
Філогенія найпростіших
Первинні живі істоти безумовно були набагато примітивнішими за своєю організацією ніж сучасні найпростіші. Частина зоологів вважають найпримітивнішими і найдавнішими з найпростіших саркодових, а саме голих амеб (нестала форма тіла, псевдоподії як органоїди руху).
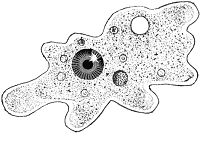
Інша частина зоологів за вихідну форму вважають джгутикових, яким властивий, крім гетеротрофного, ще й гетеротрофний спосіб живлення. Вони виходять з того, що вважають автотрофний спосіб живлення давнішим за голозойний (живлення твердою органічною їжею, яка перетравлюється в травному тракті. Один з видів гетеротрофного живлення).
Вважаючи саркодових найпримітивнішими, все ж наявність джгутиків у гамет саркодових, дає підстави припустити, що первинними одноклітинними були гетеротрофні примітивні джгутикові, від яких походять і саркодові, і більш високорганізовані гетеротрофні і автотрофні джгутиконосці.
Форамініфери, сонцевики й радіолярії походять від амебоподібних предків, еволюція яких пішла в напрямку спеціалізації.
Інфузорії – найскладніше організовані найпростіші. Їх походження не зовсім ясне через диференціацію ядерного апарату й особливості статевого процесу (кон’югація). Вважають, що інфузорії це група, яка давно відокремилась від давніх джгутикових. Сисні інфузорії походять від війчастих, але втратили війки черз пристосування до паразитизму.

Споровики – одноклітинні організми, які ведуть виключно паразитичний спосіб життя. Існують ці найпростіші в ораганах травлення, виділення, розмноження і в крові тварин та людини. Це збірна група, що об’єднує тварин різного походження.

Походження багатоклітинних
Перехідними до багатоклітинних форм були колоніальні джгутикові. Спочатку колонія складалась з одноманітних клітин, а потім виникла диференціація клітинних елементів. Це найстаріша й дуже поширена гіпотеза. У 70-х роках ХІХ століття Е. Геккель запропонував теорію гастреї (інвагінаія).

Ернст Геккель (1834 - 1919)
І. Мечников запропонував теорію паренхімули (імміграція). Паренхімула – стадія ембріонального розвитку, коли двошаровий зародок утворюється шляхом імміграції клітин бластули у бластоцель. Диференціація клітин на локомоторні і фагоцити. Ця гіпотетична предкова форма названа фагоцителою.

Ілля Ілліч Мечников (1845-1916)
Теорія фагоцители - гіпотеза походження багатоклітинних тварин, запропонована І. І. Мечніковим у 1886 р.
Згідно теорії фагоцители вихідною формою багатоклітинних є гіпотетична тварина - фагоцитела (інша назва - паренхімела), яка побудована, як і личинка сучасних нижчих хребетних - паренхімула, з шару поверхневих клітин - ектодерми (кінобласт) і внутрішньої клітинної маси - паренхіми (фагоцитобласт). Кінобласт виконує функцію відмежування, зовнішнього обміну і руху; фагоцитобласт - внутрішнього обміну і внутрішньоклітинного травлення. З кінобласта і фагоцитобласта, згідно теорії фагоцители, в ході еволюції виникла вся різноманітність форм тканин багатоклітинних тварин.
Філогенія губок

Губки поєднують низький рівень організації і високий ступінь спеціалізації:
Відсутня диференціація тканин, є лише диференціювання клітин на хоаноцити (спеціалізовані джгутикові клітини в губок, які здатні в ході трансдиференціації перетворюватись у статеві клітини) й покривні клітини (археоцити) та клітини мезоглеї (наповнена водою сполучна тканина).
Відсутні нервові й мязові клітини.
Відсутні зародкові листки.
Губки – це багатоклітинні тварини. Близькі до колоніальних комірцевих джгутикових. Клітини, подібні до хоаноцитів, зустрічаються майже виключно у комірцевих джгутикових. Це дає підстави вважати, що губки походять від гіпотетичних давніх комірцевих джгутикових, і являють собою особливу гілку багатоклітинних тварин, які виникли від одноклітинних незалежно від інших гілок.
Губки утворюють колонії, де втрачається індивідуальність особин, що складають її. Розвиток складної системи каналів з джгутиковими камерами й особливо розвиток складного скелету зумовили збереження цієї давньої групи тварин, незважаючи на низький рівень організації.
Губки сліпа гілка у розвитку тваринного світу. Вони найпримітивніші й найдавніші з сучасних багатоклітинних. Сидячий спосіб існування не сприяє ускладненню їх будови. Простота їх будови, відсутність кишечної порожнини, завершення ембріонального розвитку стадією паренхімули підтверджують думку більшості зоологів, що ці тварини походять від паренхімел. Більшість видів колоніальні форми, менше поодиноких, характерних для предків. Відомо 5000 видів, більшість морських.
Філогенія кишковопорожнинних

Тип поділяється на два підтипи: Жалкі (Cnidaria) та Нежалкі (Acnidaria). Найдавнішим є перший, а серед нього найпримітивніший гідрозом (відсутність ектодермальної глотки, проста будова гастральної порожнини, мезоглея переважно тонка безструктурна, дифузна, нервова система).
З гідрозоями генетично пов’язані і сифонофори, хоча вони мають складнішу будову. Це обумовлено диференціюванням колонії. Колонії сифонофор – це модифіковані нащадки поліплоїдної колонії.
Кишковопорожнинні походять від колоніальних джгутикових (не ті, від яких походять губки). Коралові поліпи і прісноводні гідри походять від спільного гіпотетичного предка поліплоїдної будови.
Предки кишковопорожнинних, на відміну від губок, тривалий час еволюціонували як вільно плаваючі форми й звичайно пізніше пристосувалися до сидячого способу життя. Таким чином, поліпне покоління є первинним. Давні поліплоїдні дали початок медузам, які стали виконувати функцію статевого розмноження. Серед сучасних кишковопорожнинних найпримітивнішими є гідроїдні, давні форми яких були вихідними в еволюції сцифомедуз і коралових поліпів, і реброплавів (нежалких). (МАРКЕВИЧ)
Філогенія червів
У Чорному та Азовському морях зустрічаються тварини, які зовні схожі на медуз. Це реброплави. Раніше їх відносили до типу Нежалкі кишковопорожнинні, а тепер виділяють в окремий тип ектодерми у напряму кишечної ентодерми на стороні, протилежної бластопору. Первинний рот у них (бластопор) або заростає, або з нього утворюється анальний отвір. Первинно- і вториннороті походять від різних предкових груп.
Філогенія сисунів і стьожкових червів
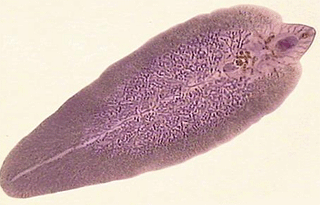 сисуни
сисуни
 сисуни
сисуни
Вважається, що і моногенетичні й дигенетичні сисуни походять від турбелярій, незалежно один від одного. Стьожкові черви – група плоских червів, дуже змінена в напрямку пристосування до паразитичного способу життя. Найближче вони стоять до моно генетичних сисунів.
Філогенія
круглих червів

Первиннопорожнинні черви походять від турбелярій, які перейшли до активнішого способу існування, у зв’язку з чим ускладнилась їхня будова.
Філогенія круглих червів вивчена вкрай недостатньо. Питання про походження та про філогенетичні зв’язки між різними класами цього типу до цього часу ще не вирішено. Най вірогіднішим є припущення про турбелярне походження різних груп. Але ще нема єдності думок про те походять ці групи від турбелярій беруть початок два чи навіть три, незалежних стобура, що привели до різних класів круглих червів. Отже. Якщо питання про спільне походження всіх класів круглих червів з турбеляріями більш-менш з’ясовано, то питання про ступінь їх спорідненості між собою вимагає додаткових ретельних досліджень.
Перехід до паразитизму
Життя в грунті серед гниючих решток, що його ведуть грунтові круглі черви, та життя в іншому організмі, що його ведуть паразити, мають багато спільного. В обох випадках жити доводиться у досить щільному середовищі, де мало кисню і де можуть міститись шкідливі речовини. Перехід від життя в грунті до життя в організмі хазяїна у круглих червів стався досить просто.
Предки паразитичних куглих червів потрапляли до організму хазяїна випадково, наприклад з їжею. Завдяки шкільній кутикулі вони могли залишатися живими у кишечнику великої тварини. Там їх оточувала цілком доступна пожива. В кишечнику хазяїна міститься суміш бактерій та поживних речовин, що розкладаються, тобто те ж саме, чим живляться грунтові круглі черви. Хижі черви могли розривати стінку кишечника своїми гострими зубцями й живитися тканинами хазяїна.
З часом зв'язок паразита з хазяїном зміцнювався. Паразити обирали певних хазяїв, удосконалювався їхній життєвий цикл. Одні паразити почали виводити яйця у зовнішнє середовище, інші пристосувалися до розселення за допомогою комах-кровососів.
У тілі людини може паразитувати близько 50 видів круглих червів, більшість їх поширені у тропіках. Найпоширеніші в нашій країні: людська аскарида, гострик, трихінела.
Філогенія
кільчастих червів

Існує кільках гіпотез стосовно походження кільчастих червів.
Гіпотеза Майєра і Ланга – походять від турбелярій.
Гіпотеза Лиманова – походить від немертино подібних предків.
Гіпотеза В. Беклемішева – «трохофорна теорія»: кільчики походять від гіпотетичного предка «трохозоона», що має трохофоро подібну будову й походить від гребневиків.
У плоских червів простір між покривами (що розвиваються з ектодерми) й кишечник (з ендодерми) заповнені паренхімою (з мезодерми). У круглих червів між органами екто-, ендо-, і мезодермального походження міститься заповнена рідиною первинна порожнина тіла. Порожина тіла кільчастих червів також заповнена рідиною, але розвивається всередині мезодермального походження. Така порожнина називається вторинною. У кільчастих червів вона поділена на сегменти попреерчними перетинками. Цікаві міркування про вдосконалення способу пересування у круглих червів, що може вказувати на ймовірний хід еволюції.
Поширення круглих червів пов’язано з використанням ними заповненої рідиною під підвищеним тиском і тому виконує опорну й захисну функції. Вона разом із смужками поздовжніх м’язів дозволила круглим червам освоїти новий спосіб пересування – заривання у грунт. Спосіб пересування нематод своєрідний. Вони пересуваються, лежачи на боці. Під час скорочення м’язів тіло згинається у спинно-черевному напрямі. Під час згинання тіла щільна кутикула майже не розтягується, і тиск у порожині тіла підвищується. Як тільки м’язи розслаблюються, червяк розгинається. Чергуючи згинання і розгинання, черв’як звивається і набуває здатності пересуватись навіть у щільному середовищі.
Однак часто є вигідним згинати-розгинати не все тіло, а лише деякі його ділянки. Щоб використати при цьому як опору порожнину з рідиною, її треба поділити на відсіки – сегменти. Підвищивши тиск в одному сегменті, можна розігнути тіло тільки в цьому місці. Саме так побудовано порожнину тіла кільчастих червів. Під час їх руху різні ділянки тіла водночас здійснюють різні рухи.
Дя тварини, яка рухається згинаючись, корисні вирости, що опираються на навколишні предмети. Це спрямовані вбік вирости кожного сегмента тіла – параподії. Згодом розвивається мускулатура, що забезпечує рух окремих параподій – «первинних ніг».
Оскільки порожнина тіла кільчастих червів сегментована, у них розвивається сегментованість інших систем (видільної, нервової, статевої, рухової).
Кільчасті черви є високоорганізованими первинноротими. Вони походять від давніх первинноротих, які в свою чергу походять від форм, близьких до гребників.
Від давніх полі хет походять не лише сучасні багатощетинкові черви, а й інші групи анелід. Особливо важливо, що полі хети є вузловою групою і в еволюції вищих первинноротих. Від них походять молюски й членистоногі.
Філогенія молюсків

Молюски походять від вториннопорожнинних червів, тіло яких складалось з небагатьох сегментів. Молюски вели активний спосіб життя, що привело до ускладнення їх будови. Схожість деяких основних етапів ембріонального розвитку молюсків і кільчиків (спіральне дробіння, личинка, трохофорного типу, телобластний спосіб утворення целому) говрить про спорідненість обох типів. Але відсутність сегментації у дорослих, гангліозна нервова система, особливості кровоносної системи вказують на значні відмінності. Однак більшість вчених вважають, що обидва типи безхребетних тварин походять від гіпотетичних вищих червів, у яких був целом і поділене на сегменти тіло. Ці ознаки мають давні примітивні молюски. Сучасні ж молюски, у зв’язку з відомим способом існування, значно відхилились у своїй будові від вихідних форм.
Примітивні молюски, мабуть, виникли тоді, коли кільчиків і членистоногих ще не існувало й не було конкурентів та захисної раковини. Пізніше, коли зявились кільчики, а ще пізніше членистоногі, у молюсків з’явились раковини. Усе це привело до зникнення метамерії деяких органів, характерної для нижчих груп м’якотілих, рух яких здійснюється спецвальною частиною тіла – ногою, а не всім тілом.
Вважають, що молюски пішли від предків, подібних до поских червів. Як відомо, частина потомків плоских червів набула здатності зариватись у грунт, рятуючись від хижаків. Молюски пішли іншим шляхом: на спинній поверхні їнього тіла сформувався захисний панцир – черепашка. Перевагу мали ті молюски, тіло яких могло повністю сховатися під цим панциром, отже він повинен був рости разом із твариною. Для забезпечення росту черепашки у молюсків утворилася особлива шкіряна складка – мантія. Вона прилягала до черепашки: між нею і тілом молюска утоврилася мантійна порожина.
У зв’язку з придбанням черепашки змінилася будова молюсків. У захованих під черепашку молюсків для пересування виник широкий сполющений виріст – нога. Перші молюски рухались на зразок війчастих червів, за допомогою хвилеподібних м’язових скорочень, що пробігали по нозі.
Захищені зверху черепашкою тварини могли живитися лише тією їжею, що знаходили на поверхні, якою рухались. Тому вже перші молюски мали спеціальний орган – тертку, що дозволяв їм зшкрябувати водорості з каміння. Рот із теркою та органи чуття знаходились на голові, рухомо пов’язаній з ногою.
Отже, тіло молюсків поділялось на три відділи: тулуб, поміщений в черепашку, ногу й голову. Хоча молюски пішли від предків із двобічною симетрєю, більшість з них стали асиметричними.
Черепашка утруднювала дихання через покриви. Щоб забезпечити потребу організму в газообміні, у молюсків виникли зябра – вирости, розтшовані у мантійній порожнині. Тип Молюски налічує понад 100 тисяч видів. Це процвітаюча група.
Філогенія членистоногих
Генетичний зв'язок членистоногих із кільчиками очевидний: розчленування тіла, схожість ЦНС, членисті ноги виникли з параподій, серце – з спинної судини, складні мета нефридії – з простих і т.п. Деякі органи членистоногих виники заново (трахеї, мальпігієві судини тощо).
З кільчаків лише полі хети могли бути предками членистоногих: у них є параподії, добре розвинений головний відділ тіла й т.ін. Ускладнення організації, які привели до виникнення членистоногих, могли здійснитися лише у полі хет, що вели активний спосіб життя. У сидячих форм організація спрощується.
Найпримітивнішими серед викопних членистоногих були трилобіти з нижнього палеозою. Тіло їх поділялось на голову й велику кількість схожих сегментів. Усі кінцівки були двогіллястими.
У будові сучасних первиннотрахейних з субтропіків і тропіків Австралії, Південно-Східної Азії, Південної Африки, Південної і Центральної Америки та Нової Зеландії поєднуються ознаки кільчастих і членистоногих. Ознаки кільчастих: червоподібна форма тіла, ноги короткі, майже не розчленовані, м’язи гладенькі, органи виділення – численні мета нефридії, дуже довга середня кишка, примітивна нервова система.
Ознаки членистоногих: придатки ніг розчленовані, є ротові кінцівки, незамкнута кровоносна система, спинна судина перетворена на серце, міксоцель, органи дихання – трахейні трубочки. Наявність первиннотрехейних підтверджує спорідненість кільчастих червів з членистоногими і можливість еволюційного преходу від першого до другого.
Предками членистоногих були тварини, схожі на трилобітів.
Клас Ракоподібні

Ракоподібні походять від трилобітоподібних членистоногих, які перейшли до швидкого пересування по дну водойм і в товщі води. Це привело до ускладнення їх організації: підвищилась диференціація тіла на відділи, відбулось злиття багатьох сегментів, ускладнилась нервова система; будова кінцівок урізноманітнилась; інтенсифікувалась робота усіх органів і систем. Деякі живуть на суші в вологих місцях.
З усіх типів тварин перехід до наземного способу життя найуспішніше здійснили членистоногі. Цей процес був дуже складним і тривалим. Основна небезпека при виході на сушу – втрата води, що приводить до порушень обміну речовин і до загибелі. Зовнішній скелет, потужні м’язи, зяброве дихання – водні.
Клас Павукоподібні

Павукоподібні походять від водних членистоногих з класу ракоскорпіонів, які поступово перейшли до існування на суші. Але павукоподібні, як правило, віддають перевагу зволоженим наземним біотопам, деякі ведуть нічний спосіб життя. Пристосування павукоподібних до існування на суші ще не дуже досконалі. Багато які види (переважно з роду кліщів) є вторинноротими тваринами.
Павукоподібні мають спільні ознаки: чотири пари ніг, ектодермальні мальпігієві судини, дві пари ротових кінцівок, що свідчить про спільність предків. З сучасних груп водних членистоногих найбільшу схожість з павукоподібними (із скорпіонами) мають мечехвости (до 80 см, морські). Тіло їх вкрите панциром і поділяється на голово груди й черевце. На голово грудях шість пар кінцівок: перші дві відповідають хеліцерам і педипальпам, чотири – ногам павукоподібних. Антени відсутні. Є схожість і в будові внутрішніх органів. На черевці у мечехвостів є ще шість пар кінцівок із зябрами. Органи виділення – коксальні залози – органи давніші за мальпігієві судини. Мечехвости – нащадки численних палеозойських ракоскорпіонів, які були споріднені з трилобітами. До того ж личинка мечохвостів дуже нагадує трилобітів. У процесі пристосування ракоскорпіонів до наземного способу життя черевні ноги з зябрами, мабуть, зрослись зі шкірою і виникли легеневі мішки, де зяброві вирости перетворились на перегородки. Дійсно, у нижчих павукоподібних (скорпіонів) легені містяться на тих же сегментах, де у мечехвостів зябра. Черевні ноги редукувались, з’явились мальпігієві судини. Отже, давні павукоподібні походять від водних членистоногих.
Клас Багатоніжки

Багатоніжки походять від давніх примітивних вищих ракоподібних, які перейшли до наземного способу життя. Пристосування до наземного існування не досконалі, тому вони ведуть прихований спосіб життя в вологих місцях, де вони захищені від висихання. У будові багато примітивних рис: голова й гомоном на сегментація, епіку тикала, одна пара вусиків, очі переважно прості, м’язи одноманітні, дихають трахеями, камери серця – по числу сегментів, ендодермальні мальпігієві судини, три пари ротових кінцівок, живляться органічними рештками і є хижаками.
Клас Комахи

Комахи походять від однієї з груп багатоніжок, які перейшли до активного життя у відкритих місцях на суші. Будова їх поступово ускладнювалась і набула досконалих адаптацій до наземного способу існування. На основі ускладненої організації виникла найхарактерніша риса більшості комах – їх здатність до польоту.
Походження комах від багатоніжок підтверджуються низкою ознак, властивих обом класам: голова добре відокремлена від інших ссегментів; одна пара вусиків; видовжене серце з крило видними мязами, які розширюють камери серця і забезпечують надходження в них в крові; органи дихання – трахеї; органи виділення – мальпігієві судини ектодермального походження; три пари ротових кінцівок (пара верхніх і дві пари нижніх); відсутня печінка. Організація багатоніжок безумовно простіша, ніж у комах, але на її основі в процесі прогресивної еволюції могла виикнути концентрація тіла комах (утворення грудей і черевця), ускладнення всіх систем органів (особливо нервової і м’язової), поява крил; перехід до внутрішнього запліднення, ускладнення онтогенезу (перехід від розвитку з неповним перетворенням до розвитку з повним метаморфозом).
Походження комах
У девонський період виникли справжні наземні рослини, що почали розселятися на суходолі. Багнисті ділянки суходолу заселялись різноманітними безхребетними, які пристосувались до наземного способу життя. Серед них були багатоніжки чи подібні їм членистоногі, кожний сегмент яких мав пару ніг.
Зявилося нове місце існування – прямостоячі рослини, що піднімалися над грунтом. Для пересування по них давнім членистоногим довелося перерозподіляти навантаження на кінцівки. Коли вони залазили на рослини, основне навантаження брали на себе ноги передньої частини тіла, що піднімали тіло вгору. Три передні пари ніг та їхня мускулатура посилювались. Це привело до відособлення грудного відділу тіла, що складаються з трьох сегментів. Наступні сегменти перестали брати участь в пересуванні, втратили кінцівки та утворили черевце. У результаті з’явився клас Комахи.
Більшість сучасних комах мають крила. Розвиток крил є найважливішим еволюційним надбанням комах. Спочатку це були невеликі бічні вирости вирости грудей, складки їх покривів. Можливо комахи використовували їх щоб швидше розігріватися на сонці, а потім пристосувались до ширяння за їх допомогою. Наступним етапом еволюції міг стати махальний політ, що властиво сучасним комахам.
Ще одне важливе надбання комах – складний ротовий апарат, що розвинувся із видозмінених кінцівок головних сегментів, що привело до урізноманітнення їжі, у першу чергу – рослинами. Комахи стали одним з найважливіших компонентів усіх наземних екосистем. Вони справили суттєвий вплив на еволюцію інших груп, особливо квіткових рослин.
Філогенія голкошкірих

Походження голкошкірих невідоме. Але встановлено, що найдавніші примітивні предк голкошкірих, вели прикріплений спосіб життя. При цьому найдавніша група мала сліди білатеральної симетрії. Тепер вважають доведеним походження голкошкірих від білатеральних предків. Про це свідчить рання личинка сучасних ехінодермат – диплеврула. Перехід до променевої симетрії був обумовлений переходом від повзання або плавання до сидячого способу існування.
Надалі відбувся вторинний перехід від прикріпленого до вільного способу існування, причому перехід не стільки до плавання, скільки до повзання. Тому морські лілії, які перейшли до плавання, не втратили променеву симетрію, а з другого боку, найбільше порушення променевої симетрії є у голу торій, які повзають і у «неправильних» морських їжаків. Філогенетичні стосунки між сучасними класами не з’ясовані.
Голкошкірі – вториннороті. Але філогенетичні зв’язки голкошкірих з іншими вторинноротими, з хордовими, відсутні. Голкошкірі – група тварин, яка дуже давно відокремилась від загальної гілки розвитку вторинноротих.
Голкошкірі відомі з кембрійського періоду. Вони пішли від нерухомо сидячих тварин, у яких виробилась п’ятипроменева симетрія. Проте більшість сучасних представників цього типу здатні до переміщення, хоча й незначного. У предків голкошкірих, що за способом життя були схожі на сучасних морських лілій, виникли водно судинні ніжки, які допомагали збирати поживу. Певне, їхні потомки повернулися ротом до низу й почали використовувати водно судинні ніжки для переміщення.
Деякі з голкошкірих втрачають променеву симетрію і вдруге набувають двобічної. У голотурій, які лежать на боці, це пов’язано з повзанням у певному напрямку, а у деяких морських їжаків – із пристосуванням до риття у грунті. Неправильні морські їжаки втрачають складний ротовий апарат і повертаються до живлення за допомогою воднодносудинних ніжок.
Отже, в історії голкошкірих багаторазово відбувалися відмови від якихось еволюційних надбань і повернення до ознак предків. Існує припущення, що личинка якихось давніх голкошкірих відмовилися переходити до малорухливого життя дорослих форм, і з цим пов’язане виникнення хордових.
Походження типу Хордові
Майже всі типи тварин виникли ще до початку палеозойської ери. Більшість із них зявилась у вендський період на його початку у тваринзявились органи й тканини, а наприкінці Землю вже населяли найрізноманітніші істоти. У вендський період було навіть більше типів тварин, ніж тепер, адже багато які з них вимерли. Від тих часів збереглася незначна кількість викопних решток, і тому родовід безхребетних відновлюють, ґрунтуючись на будові їхніх пізніших представників.
Поряд із іншими у палеозойську еру набуває розвитку ще один тип тварин. Його основні особливості сформувалися в умовах конкуренції з іншими вже сталими типами. До нього належать найкрупніші та найбільш високорганізовані тварини. У більшості екосистем саме представники цього типу виявляються найважливішими компонентами. Це Хордові.
Основні гіпотези походження хордових
Одна із перших концепцій, що пов’язує план будови хордових з планом будови інших безхребетних, була запропонована французьким науковцем Жоффруа Сент-Іллером ще в першій половник ХІХ століття. Він вважав, що членисті тварини (кільчасті черви й членистоногі) можуть розглядатись як переверуті хребетні. Саме ця ідея буда в основі публічної суперечки між Жоффруа Сент-Ілером та Жоржем Кювьє, що відбулась у Паржі в 1830 р. Перемогу отримав Жорж Кювьє. Однак до поглядів Сент-Іера науковці повертались неодноразово впродовж усього ХІХ століття. Цю точку зору підтримував видатний зоолог Антон Дорн. На початку ХХ століття ідея походження хордових від перевернутих на спину членистоногих відстоювали Гаскелл, Таттен та ін.
На початку нинішнього століття були розроблені гіпотези походження хордових від кишководихаючих – морських червоподібних організмів, які мешкали в тощі грунту. У цих тварин як і у хордових наявні зяброві щілини, а в передній частині тіла – опорний вирст кишечника, який порівнювали з хордою. Однак кишководихаючі відрізнялись від хордових зворотнім потоком крові, розташуванням серця, будовою нервової системи тощо. Здійснювались спроби встановлення походження хордових від голкошкірих, немертин, молюсків та інших груп безхребетних, але жодна з них не мала успіху.
Реакцією на невдалі спроби зоологів вирішити цю проблему стала розробка так званих неотенічних гіпотез, що стверджували походження хордових від личинок асцидій. Дорослі асцидії – прикріплені морські тварини, фільтруючі воду через місткий зябровий мішок і вловлюючи повислі органічні частинки з води. вони не мають ні хорди, ні нервової трубки і не схожін а хордових тварин, але, як відомо, личинки асцидій мають і хорду, і нервову трубку, які втрачаються під час метаморфозу. На основі цього науковці (Берилл, Уитер, Боун та ін.) вважають, що хордові походять від ставших статевозрілими личинок асцидій. Проте, виникає питання звідки взялися самі асцидії. Сумнівним є і той факт, що хорда й нервова трубка виникає саме у личинок, котрі живуть один-два дні й не харчуються. Кожна з гіпотез має як свої недоліки, так і корисні дані для реконструкції шляхів походження хордових.
Усі білатерально-симетричні орагінзми діляться на дві групи – первинно- і вториннороті. Назви «первиннорот» (Protostomia) і «вториннороті» (Deuterostomia) походять від способу розвитку ротового отвору в ембріогенезі. Так, у первинноротих бластопор (виникаючий в ембріональному розвитку отвір первинного кишечника) частково й повністю переходить в ротовий отвір. У вторинноротих (наприклад, голкошкірих) бластопор стає нанальним отвором, а рот проривається заново на передньому кінці личинки. Окрім цього є й інші відмінності. У первинноротих характер дроблення заплідненого яйця спіральний, а у вторинноротих – радіальний (рис ).
Первиннороті
(спіральне дроблення)
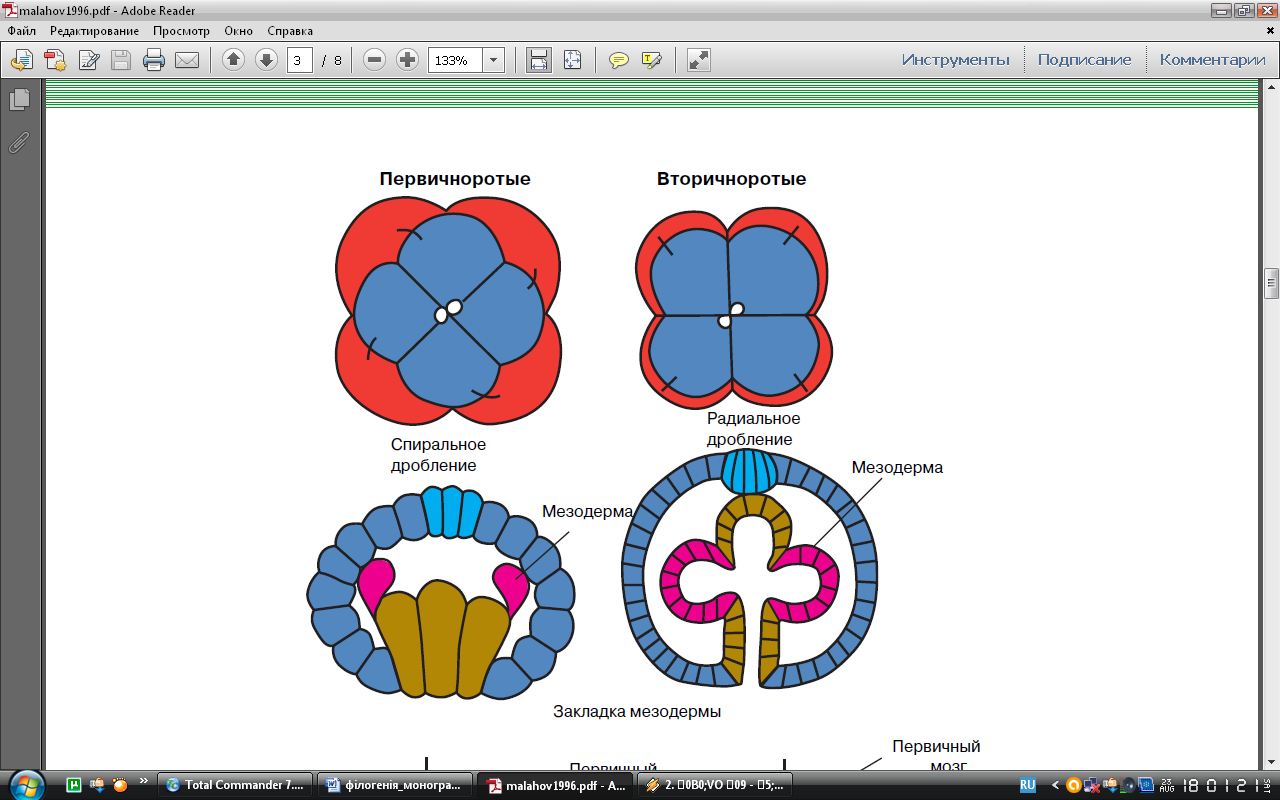
Вториннороті (радіальне дроблення)
У первинноротих стінки вторинної порожнини тіла походять від двох клітин, а вторинноротих у результаті вип’ячування карманів ембріонального кишечника (рис )

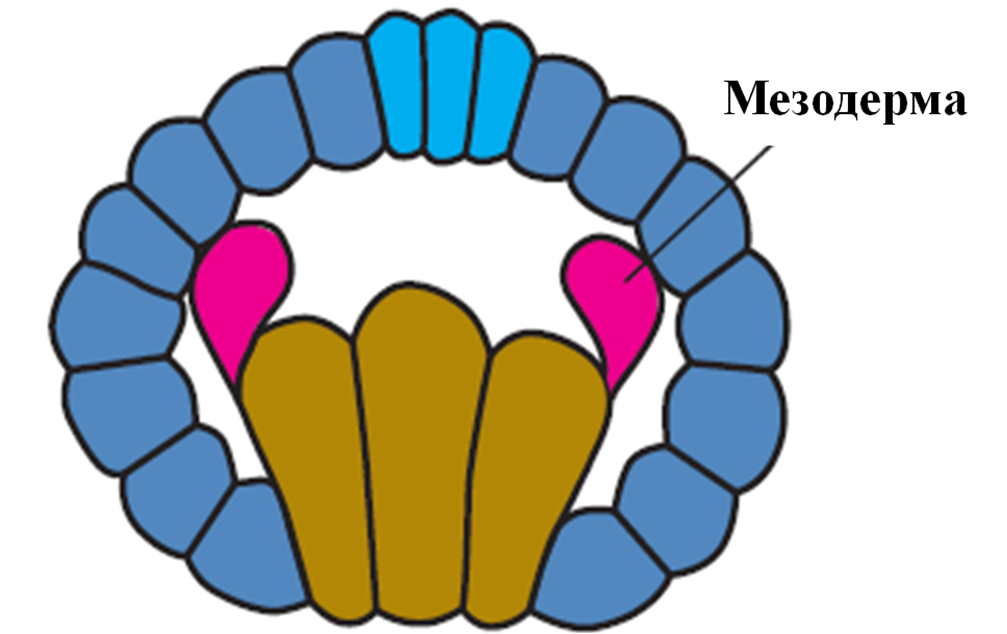
Первиннороті Вториннороті
Рис. Закладання мезодерми
У первинноротих зачаток первинного мозку дає початок мозку дорослих форм, а у вторинноротих – редукується, а новий нервовий центр виникає в іншому місці заново.
Хордові мають типове радіальне дробіння яйця, вторинна порожнина тіла – целом – формується за рахунок вип’ячування карманів первинного кишечника, а в нервовій системі немає слідів первинного мозку. Отже, логічно відносити хордових до вторинноротих тварин. Окрім хордових до них належать погонофори, напівхордові та голкошкірі. До складу первинноротих входять плоскі й круглі черви, кільчасті, молюски, членистоногі.
Шляхи еволюції вторинноротих тварин
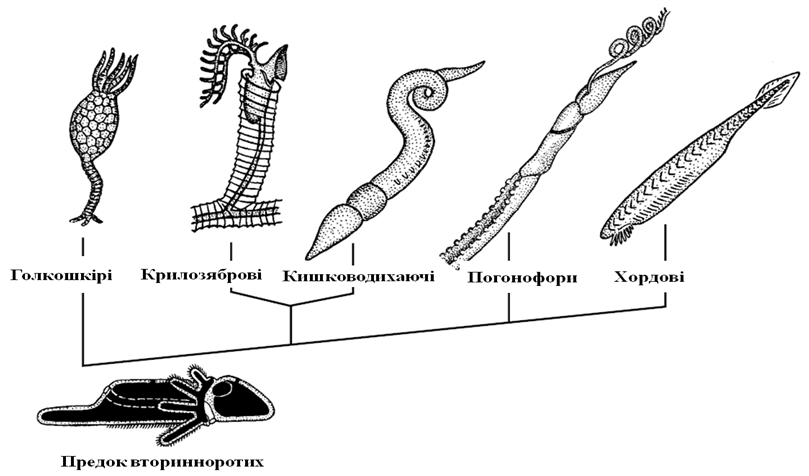 Уважається,
що спільний предок вторинноротих вів
придонний спосіб життя, повзаючи на
черевній стороні за допомогою війчастої
смужки (рис.
).
Уважається,
що спільний предок вторинноротих вів
придонний спосіб життя, повзаючи на
черевній стороні за допомогою війчастої
смужки (рис.
).
Рис. Філогенетичні зв’язки між вторинноротими тваринми
Він мав три пари целомічних мішків і щупальцевий апарат, за допомогою якого вловлював харчові частинки. Подалі могло відбутися відокремлення заднього відділу тіла, за допомогою якого організми тимчасово заривались у верхні шари грунту. Предки голкошкірих перейшли до сидячого способу життя, за допомогою щупалець відловлювали завислі у воді часточки їжі. Радіальна симетрія голкошкірих є насідком сидячого способу життя, який серед сучасних форм зберігається у морських лілій, у той час як морські зірки, морські їжаки, офіри і голоуторіі здатні повільно переміщуватись з місця на місце.
Напівхордові в якості основного органу руху стали використовувати передротовий відділ тіла: хобот кишководихаючих або головний щит крилозябрових. Досить часто напівхордових вважають близькими предками хордових. Насправді ця група не є настільки близькою, оскільки, наприклад, має два нервові тяжі, тільки один із яких може бути оцінений гомологом нервової трубки хордових. У них наявний сліпий виріст кишечника – стомо хорд, який утворився як опорна структура хобота і не має прямого відношення до хорди хордових тварин.
Сучасні погонофори представлені сидячими організмами, котрі мешкають всередині виділених ними ж трубках. У дорослому стані фони позбавлені кишечника і харчуються за рахунок симбіотичних хемосинтезуючих бактерій, які окислюють сірководень або метан. Імовірно, їхні предки харчувались цими бактеріями, збираючи їх із придонного шару води за допомогою щупалець. У результаті вони почали культивувати бактерії в клітинах кишечника. Личинки однієї із груп сучасних погонофор – вестиментифер – на ранніх стадіях розвитку збирають бактерії з поверхні грунту, а потім починають їх культивувати всередині видозміненого кишечника, після чого рот і анус у них редукується. Особливості ембріонального розвитку і будови нервової системи дозволяють розглядати погонофор як найближчу до хордових групу вторинноротих. Предки хордових таким же чином спочатку збирали харчові частинки за допомогою навколоротових щупалець, які й нині збереглись у ланцетника. Однією з особливостей еволюції хордових стало використання заднього, четвертого відділу тіла як органу активного руху. Він наявний у всіх вторинноротих. Голкошкірі й крилозяброві використовують його як прикрілююче стебло. У кишководихаючих цей відділ функціонує лишке в молодих особин, за допомогою нього вони закопуються в грунт. У погонофор цей відділ активно функціонує все життя: він оснащений щетинками, розташований на задньому кінці трубки й завядки його рухам погонофори заглиблюють задній кінець трубкив товщу грунту по мірі свого росту. Хордові – єдина група вторинноротих, котра використовує цей відділ як орган активного плавання. У зв’язку з активною руховою функцією цього органа в ньому відбувається повна сегментація мускулатури й розвивається хорда.
Ще один важливий факт еволюції хордових пов'язаний з інверсією сторін тіла. У філогенетичному розвитку хордових відбулося перевертання з морфологічно черевної на морфологічно спинну сторону. Це перевертання є порівняльно-анатомічним фактом, що витікає з співставлення плана будови хордових з іншими групами вторинноротих. Таке перевертання могло відбутися під час переходу від вловлювання завислих часточок їжі у воді за допомогою щупалець, що нині використовують сучасні крилозяброві, до збору їх з верхнього шару грунту, як це спостерігається з сучасними ланцетниками.
На сьогодні дискусія про походження хордових не вичерпана.
Перші представники типу Хордові з’явилися, напевне, у кембрійський період. Вважають, що найближчими родичами хордових є голкошкірі, незважаючи на усю зовнішню несхожість цих двох типів.
Формування основних ознак типу Хордових пов’язане з тим, що їхні предки вели життя вільно плаваючих фільтраторів. Багато які тварини живляться дрібними частками, що плавають у воді. Для цього вони повинні мати якийсь цідильний пристрій. Струмінь води забезпечують джгутики або війки, іноді – скорочення відповідних м’язів. Давні хордові використовували інший спосіб – вони переміщувалися у товщі води самі та фільтрували її набігаючий струмінь. Їхнім цідильним пристоємбула глотка (початкова частина травної системи), пронизана зябровими щілинами. Ці щілини не лише затримували поживу. Оскільки їхні стінки добре омивались водою, то в них відбувався і газообмін. Тому у хордових дихальна й травна система пов’язані одна з одною.
Із можливих способів плавання (робота війок, плавців, кінцівок тощо) давні хордові обрали дуже простий: вони хвилеподібно вигинали тіло. Для забезпечення цих рухів з боків тіла розміщувалися м’язи. Щоб вигинати тіло в потрібній ділянці, а не по всій довжині, м’язи були сегментовані. З часом виник осьовий стрижень, який допомагав витрачати менше зусиль під час плавання. Під час згинання тіла стрижень «накопичував» у собі зусилля і віддавав їх, коли тіло вирівнювалось. Так виникла хорда.
Плаванням та живленням таких тварин керувала центральна нервова система. Це була трубка з нервових клітин, що тягнулася уздовж усього тіла; від неї відходили нерви до кожного з сегментів м’язів. Відокремився головний мозок, розвинулись органи чуття.
Нащадками цих гіпотетичних тварин є сучасні й личинковохордові. Вони тепер ведуть не такий спосіб життя, завдяки якому виникли хордові. Як відомо, це було плавання за рахунок вигинів тіла, яке дозволяло фільтрувати воду. Безчерепні стали переважно природними тваринами, а покривними –сидячими, або плаваючими реактивним способом. Головний напрямок еволюції хордових був пов'язаний із тваринами, які продовжували плавати за рахунок зусиль м’язів. Цей спосіб життя приводив до вдосконалення внутрішнього скелету, мускулатури, подальшого розвитку органів чуття і нервової системи.
Вигини тіла забезпечувала сегментована мускулатура, і для її зміцнення було необхідно укріпляти місця прикріплення м’язів. З іншого боку, було необхідно захистити від можливих ушкоджень центральну нервову систему. Тому вздовж спинного мозку й хорди у цих тварин виник ряд хребців, що утворюють хребет. Хорда – це однорідний пружний стрижень; на відміну від неї хребетний стовп складається з твердих частин і тому сегментований. Крім того, активно плаваючим тваринам важливо захистити головний мозок. Тому у них формується череп – головна частина внутрішнього скелету.
Хордові, які відносяться до еволюційного напрямку, у якому виникли хребет і череп, належать до підтипу Хребетні.
Перші представники підтипу Хребетні належали до класу Щиткові. Вони населяли Землю від кембрійського до девонського періоду. У щиткових був добре розвинутий зовнішній скелет (панцир), який складався з окремих щитків. Панцир найкраще розвивався на передній частині тіла.
Більшість щиткових були придонними тваринами й живилися безхребетними. У разі небезпеки вони відпливали, енергійно працюючи хвостом. Розставлені в боки краї панцира щиткових працювали при цьому як крила літака: піднімали їхнє тіло догори у товщу води.
Згодом у воді з’являлися досконаліші хижаки. Щиткові вимерли, але й до сьогодні збереглися їхні родичі – представники класу Круглороті.
За деякими ознаками круглороті нагадують риб, раніше їх навіть відносили до класу Риби. Проте у круглоротих є суттєва відмінність – відсутність щелеп. Тому класи Щиткові і Круглороті об’єднують в надклас Безщелепні. Решту хребетних об’єднують в надклас Риби та надклас Чотириногі, які утворюють групу Щелепнороті.
Виникнення риб пов’язане з появою щелеп. Щелепи розвивалися із зябрового скелету, що складається з окремих частин. Зябровий скелет міног, навпаки, цілісний. Це пов’язане з тим, що коли мінога присмоктується до жертви, вода для дихання не може надходити через рот, і тому заходить крізь зяброві отвори в зяброві мішки. Видих мінога робить стискуючи м’язами глотку з зябровою решіткою, а вдих – розслаблюючи м’язи. Пружна зяброва решітка розширюється та засмоктує воду у зяброві мішки. Будова рота круглоротих зумовлена здатністю присмоктуватися до здобичі.
Походження риб
Інші хребетні добували корм, активно плаваючи. Поступово фільтрування замінилося хижацтвом. Деякі жертви намагалися утекти від хижака або вирватися з його рота. У відповідь хижаки стали рухливішими й набули пристосувань для утримання здобичі. Як відомо, вода для дихання потрапляла в глотку через рот, а виходила крізь зяброві щілини. Отже, здобич могли затримувати зяброві дуги. У зв’язку з пристосуванням до утримування здобичі одна із зябрових дуг перетворилась на щелепи, а наступна стала використовуватись для прикріплення щелеп до черепа.
Виникнення щелеп відкрило перед давніми рибами нові можливості. Щелепи дозволяли вбивати крупну здобич, відкривати від неї шматки. У давніх риб зберігався успадкований від щиткових зовнішній скелет із окремих лусок (плакоїдних). Добре розвинені луски вкривали й щелепи, виконуючи функцію зубів. Озброєні зубами щелепи, «працювали» набагато краще.
У хижаків і в їх жертв удосконалювались центральна нервова система, органи чуття та опорно-рухова система. Потрібно було не тільки швидко плавати, але й легко робити повороти. Для цього давніми рибами використовувались бічні шкіряні складки, що тягнулися вздовж тіла.
Оскільки основне навантаження виконували передні та задні ділянки складки, в цих місцях утворились парні плавці: грудні та черевні. Опору парним плавцям забезпечують пояси плавців грудний і тазовий.
Разом із розвитком парних плавців удосконалювалися непарні, які рухають тіло вперед. З них найпотужнішим став хвостовий плавець – основний рушій риб.
Перші риби з’явилися наприкінці силурійського періоду. Їхніми предками були щиткові. Три групи риб еволюціонували незалежно одна від одної.
Девонський період був часом найбільшої різноманітності представників класу Панцирні риби; у кам’яновугільний період вони вимерли, не залишивши потомків. До цього класу належали риби різних розмірів, які вели в основному придонний спосіб життя.
Дві інші групи риб, клас Хрящові риби та клас Кісткові риби процвітають і дотепер.
Риби виникли в прісних текучих водоймах і лише потім, хоча й досить швидко, заселили Світовий океан.
Походження чотириногих
Хребетні тварини мешкають у воді й на суходолі. У воді переважають риби, а на суходолі – чотириногі, що походять від риб.
Найяскравішою ознакою чотириногих є наявність у них двох пар кінцівок, що розвинулися із парних плавців риб.
У девонський період у прісних водоймах мешкали кистепері риби. Це були хижаки до 1,5 м довжини, з міцними плавцями. Вони мали легені, завдяки яким могли дихати у бідній на кисень воді. Подібно до деяких сучасних риб (муловий стрибун) девонські кистепері риби могли іноді виходити на суходіл. Ті кистепері, які пристосувалися до переповзання суходолом, мали перевагу над іншими. На суходолі було багато корму (безхребетні заселили його вже давно) й ще не було хижаків. Під час пересихання однієї водойми можна було переповзти в сусідню.
Повзаючи суходолом кистепері риби здійснювали рухи, що нагадували плавання. У міру перетворення плавців на кінцівки рибам (чи вже чотириногим) пересуватися суходолом ставало все легше. Спочатку їм було важко вдихнути повітря в легені, оскільки їхнє тіло було притиснуте до землі своєю вагою. Тому риби дихали на суходолі крізь шкіру. Проте з розвитком кінцівок перші чотириногі змогли піднімати тіло над поверхнею землі й дихати легенями.
Наприкінці девонського періоду з’явились перші чотириногі. З них найбільш відомі іхтіостеги. Вони поєднували в собі ознаки риб (тіло сплюснуте з боків, укрите лускою, плавальна лопать на хвості) з ознаками чотириногих (кінцівки). Більшість часу іхтіостеги проводили у воді, але їхні кінцівки були придатними і для пересування на суходолі. Девонські чотириногі з далекими предками сучасних. Перші чотириногі належали до класу Земноводні.
Рухаючись (повзаючи) суходолом, кистепері риби всією вагою тіла спиралися на плавці. Тонкі плавцеві промені не могли витримати такого навантаження. І з часом їхня кількість зменшувалася, а міцність збільшувалася. Базалії утворили скелет плеча і передпліччя (стегно і гомілка), а із променів плавців (радіалій) утворилися кисті й ступні. Із грудних плавців утворилися передні кінцівки, а із черевних – задні. Утворились членисті кінцівки, окремі відділи яких з’єднані суглобами. Опору кінцівкам дають пояси: переднім – плечовий, а заднім – тазовий. Надалі кінцівки чотириногих змінювалися у відповідності до способу життя їхніх власників.
На кінцях пальців часто є кігті, що складаються з рогової (білкової) речовини. Іноді вони перетворюються на копита та нігті або зовсім зникають.
Із виходом хребетних на суходіл способи їхнього пересування ускладнились.
Пересуваючи кінцівки, чотириногі можуть повзати, а відірвавши черево від землі, - ходити та бігати. Якщо під час повзання тіло притискується між якимись предметами, то ноги можуть стати непотрібними та зникнути.
Ходити й бігати можна на чотирьох кінцівках або на двох, випрямивши тіло більш-менш вертикально. Крім цього можна перейти до стрибків.
Освоюючи дерева, чотириногі навчилися лазити по них та стрибати з гілки на гілку. Збільшення довжини стрибка зумовило перехід деяких чотириногих до ширяння на розтягнутих шкірних складках, а деяких – до польоту за допопмогою крил.
Деякі чотириногі перейшли до пересування у грунті, розгрібачи собі шлях міцними лапами або продавлюють вузькі ходи та притискуються ними.
Нарешті, деякі чотириногі повернулися у воду. Одні плавають, вигинаючись усім тілом чи використовуючи хвіст, а інші загрібають воду кінцівками. На кінцівках можуть утворюватись плавальні перетинки, а в деяких ноги перетворюються на ласти.
У чотириногих хребет міцніший ніж у риб. Тіло риб підтримується з усіх боків з водою, а наземним чотириногим доводиться витрачати додаткову силу. В амфібій збільшується число відділв хребта: виникають шийний і крижовий відділи.
На суходолі зручніше повертатися не всім тулубом, а лише повернути голову (щоб щось роздивитись, або схопити щелепами). Тому відокремлюється шийний відділ хребта. На задні кінцівки припадає основна вага тіла. Тому тазовий пояс з’єднаний з крижовим відділом хребта. У амфібій і шийний, і крижовий відділи хребта мають по одному хребцю, інших чотириногих кількість хребців у цих відділах збільшується.
Стегоцефали
Перші чотириногі – стегоцефали, або панцероголові земноводні. Вони успадкували від риб покривні кістки черепа. Стегоцефали виникли в девонський період (іхтіостеги), а в кам’яновугільний були головною групою великих наземних тварин. Найкрупніші з них сягали 5 м завдовжки.
Однак еволюція не зупиниася. Виникли плазуни – тварини, краще пристосовані до умов суходолу. Предками плазунів були тварини, які займали проміжне місце між амфібіями й плазунами. Їх або вважають групою стегоцефалів, або вирізняють в окремий таксон. Деякі вчені вважають, що такі тварини (і плазуни) пішли від риб незалежно від амфібій.
У пермський період стегоцефалів почали витісняти плазуни, а у тріасовий останні великі земноводні зберігалися у воді. У юрський період усі стегоцефали зникли.
Пердки сучасних земноводних
В епоху панування плазунів збереглися лише невеликі навколо водні земноводні, які мешкали у високогір’ях або в холодильних районах, адже земноводні легше переносять холод, ніж плазуни (коливання температури у воді менші). Ці тварини і стали предками сучасних земноводних.
Так, безхвості пішли від великих стегоцефалів, які мешкали навколо високогірних річок та стрибали у воду, рятуючись від хижаків. Тіло таких тварин поступово вкорочувалось. Личинки безхвостих – пуголовки – набули обтічної форми тіла, що допомагало протистояти силі бурхливих гірських потоків. Оскільки в гірських потоках мало придатних до їжі безхребетних (їх зносить течія), пуголовки пристосувались до живлення рослинності.
Виникнення плазунів
Земноводні не змогли остаточно позбутися залежності від водного середовища. Їхнє розмноження і розвиток, як правило, пов’язані з водою, їхня волога шкіра висихає на сухому повітрі. У кам’яновугільний період амфібії поширились у вологих місцях, але посушливі та віддалені від води території були для них важкодоступними.
Ці місця існування зайняла інша група хребетних, у якої пристосування до життя на суходолі було тривалішим, ніж в амфібій, і зміни суттєвішими. Наприкінці кам’яновугільного періоду виникли перші плазуни. Їхні головні відмінності від амфібій – це здатність відкладати яйця, що розвиваються на суходолі та суха шкіра. У рептилій зародок міститься у яйці. Від висихання і пошкодження його захищають яйцеві оболонки. Але кожний зародок має розвиватись у рідкому середовищі. Тому тіло зародка рептилій оточене рідиною, яка міститься всередині зародкової оболонки – амніон. Інші зародкові оболонки (серозна й алантоїс) забезпечують дихання і екскрецію зародка. Зародкові оболонки лежать у середині яйцевих і є частиною самого зародка. Можна сказати, що рептилії взяли із собою в яйце частину водного середовища, в якому і відбувається розвиток.
Температура тіла плазунів
Як і амфібії, переважна більшість сучасних рептилій – холоднокровні тварини. Температура їхнього тіла залежать від температури навколишнього середовища. Так, прудкі ящірки вранці мають таку ж температуру тіла, як і остигла за ніч земля. Мляво виповзаючи на відкриті місця і гріються на сонці. Температура їх тіла підвищується і вони починають шукати поживу.
Температура тіла багатьох вимерлих рептилій була вищою від температури середовища. Деякі з них були настільки великими, що їхнє розігріте за день тіло просто не встигало охолонути за ніч.
Виникнувши в кам’яновугільний період, рептилії були з пермського до крейдяного періоду пануючою групою наземних тварин. У тріасовий період від них пішли ссавці і птахи. Наприкінці мезозойської ери великі плазуни вимерли. Із безлічі груп плазунів, які населяли Землю в минулому, тепер збереглися лише 4 ряди й 700 видів. Це ряд черепахи, дзьобоголові, лускаті, крокодили.
Різноманітність амфібій стримувалась двома факторами: вони не могли жити у посушливих місцях далеко від води і не могли вживати рослинний корм. Рослинний корм важче перетравлювати ніж тваринний, його основну частину складає клітковина. Рептилії розв’язали обидва ці завдання і перемогли амфібій у конкурентній боротьбі.
У пермський період значного поширення набула одна з ранніх груп рептилій – звіроподібні. Деякі з них мали великі гребені вздовж спини. Це дозволяло таким рептиліям нагріватися, підставляючи гребінь сонцю, та охолоджуватися, повертаючи його так, щоб сонячні промені ковзали вздовж нього.
Деякі зі звіроподібних стали рослиноїдними. Це були крупні тварини з великим об’ємом кишечника. Вони збільшувалися в розмірі, щоб одночасно перетравлювати більше корму; хижаки збільшувалися, щоб легше справлятися з рослиноїдними. Велике тіло таких тварин краще утримувало тепло, що дозволяло мати вищу температуру тіла, ніж навколишнє середовище. Завдяки цьому обмін речовин відбувається швидше.
Наприкінці перського періоду поширилася одна з груп звіроподібних – звірозубі рептилії. Деякі з них були великими хижаками. Інші живились комахами. Розміри тіла таких звірозубих зменшувалися, і це повинно було привести до зниження температури тіла. Але ці тварини вже пристосувалися до високої температури тіла й високої швидкості життєвих процесів. Щоб зберегти на цьому ж рівні температуру тіла, вони набули здатності підтримувати її за рахунок енергії, отриманої від корму – стали теплокровними. Для зменшення втрат тепла в них з’явився волосяний покрив. Від таких тварин пішли ссавці.
У тріасовий період звіроподібних потіснили інші групи рептилій. Від тріасових рептилій залишився лище один вид – гатерія (з ряду Дзьобоголові). Пізніше виникли лускаті, до яких належать більшість сучасних рептилій.
Ще наприкінці пермського періоду виникли текодонти – група, від якої пішли крокодили, динозаври, літаючі ящери та птахи. Динозаври виникли наприкінці тріасового періоду, згодом з’явилися перші літаючі ящери. Море населяли іхтіозаври та плезіозаври, які панували в юрський та крейдяний періоди.
До динозаврів належали найкрупніші наземні тварини за всю історію планети. Багато які з них були рослиноїдними, деякі з них сягали 30 і навіть 45 м завдовжки. Багато які динозаври пересувалися на двох ногах. Величезні динозаври розігрівалися за рахунок своєї маси (до 70 – 130 т), а дрібні рухливі могли бути теплокровними.
Динозаври відзначалися складною поведінкою. Їхні скам’янілі сліди свідчать, що вони часто жили стадами, деякі з них турбувалися про потомство.
Деякі текодонти переходили до життя на деревах і перестрибували з гілки на гілку. У них розвивалися шкірясті крила, які дозволяли спочатку пларенувати, а згодом і літати махальним польотом. Так виникли літаючі ящери. Крило являло собою велику складку шкіри, яка натягувалася на один з пальців передньої кінцівки. Перші літаючі ящери були розміром з ворону. Наприкінці крейдяного періоду розмах крил деяких з них сягав 12 м.
У продовж мезозойської ери відбувалося вимирання тих чи інших груп тварин: склад видів, які населяи Землю, змінювався багато разів. На зміну вимерлим видам з’являлися нові, і панування рептилій посилилося, а нові види перестали виникати. Вимирання бегеточисельних і різноманітних рептилій йшло повільно, різноманітність наземних рептилій падала поступово.
Великі рептилії досконало пристосувались до певних умов життя і втратили еволюційну «гнучкість». Зміна умов призвела до вимирання багатьох видів мезозойських плазунів. Вони вимерли не через конкуренцію з іншими групами тварин. Після зникнення великих рептилій минув тривалий час, доки їхнє місце в екосистемах зайняли ссавці та птахи, які виникли ще за часів панування рептилій.
Вплив розмірів на температуру тіла
Чим крупніша тварина, тим легше їй підтримувати високу температуру тіла. Виділення тепла відбувається у ході життєвих процесів масою тіла. Тепло втрачається через поверхню тіла. Тому, чим менше відношення площі поверхні до маси тіла. Тим простіше тварині зберігати тепло.
Із збільшенням розміру тіла відношення площі поверхні до маси зменшується.
Маленьким тваринам складніше розігріти тіло, а великим охолодити. Температура тіла величезних динозаврів напевне була високою і мало змінювалася під впливом умов зовнішнього середовища.
Походження птахів
Будова птахів досить одноманітна, хоча їх 8600 видів.
У тріасовий період дві групи плазунів почали освоювати повітряне середовище. Для польоту потрібні крила. Одна група плазунів (літаючі ящери) з цією метою використовувала шкірясті перетинки, друга кінцівки, вкриті сплощеними лусками (пір’ям). Від другої групи й пішли птахи.
Рештки найдавнішого птаха протоавіса («праптаха») знайдені нещодавно в Північній Америці. Його вік 225 млн років. Краще досліджений пізніший птах – археоптерикс. Його рештки (вік 150 млн років) знайдено в Європі.
Археоптерикс поєднував у собі ознаки плазунів та птахів. Ознаки плазунів: довгий хвіст з багатьох хребців, зуби на щелепах, на крилах пальці з кігтями, відсутність дзьоба. Ознаки пташині: передні кінцівки перетворені на крила, тіло вкрите пір’ям.
З достовірністю важко сказати, як «злетіли» предки птахів. Можливо, у них спочатку з’явилося пір’я як термоізолюючий шар. Предками птахів могли бути деревні тварини (протиставлення заднього пальця), що в пошуках здобичі стрибали з гілки на гілку. Поступово стрибки продовжувались завдяки ковзанню в повітрі на оперених крилах. З інши поглядів, предки птахів бігали по землі й підстрибували за своєю здобиччю – літаючими комахами. Помахи опереними передніми кінцівками допомагали їм підстрибувати, стернувати, а згодом і спурхувати на гілки дерев, рятуючись від хижаків.
Походження ссавців
У пермський період виникли звірозубі рептилії. До цієї групи належали і великі хижаки, і невеликі всеїдні тварини.
У більшості рептилій усі зуби мають схожу конічну форму й забезпечують утримання здобичі. Іноді рептилії мнуть здобич щелепами або відривають від неї шматки. У шлунок під час цього потрапляють великі шматки їжі. Звірозубі рептилії пристосувалися пережовувати (розмелювати) їжу зубами, змішуючи її зі слиною. Це зробило травлення ефективнішим.
Звірозубі жували зубами, розташованими в кутках рота (сила їх стискання сильніша). Ці зуби набували широких плоских поверхонь. Такі зуби називають корінними (кутніми): ними зручно жувати, але не зручно відкушувати. Для відкушування використовувались передні зуби з гострим ріжучим краєм – різці. Між різцями й кутніми містилися крупні зуби – ікла. Ними звірозубі утримували жертву, вбивали її та розривали на шматки. Отже, відбулася диференціація зубів, розподіл функцій між їх групами. Таку будову зубної системи успадкували й потомки звірозубих – ссавці.
У тріасовий період виникли нові групи крупних рептилій. Конкуренції з ними уникали ті звірозубі, які в ході еволюції ставали дрібнішими. Багато які з них вже пристосувалися до життя за високої температури тіла завдяки своїм великим розмірам. Зменшення розміру таких тварин сприяло розвиткові в них теплокровності. Для збереження тепла виник волосяний покрив. Малята обігрівалися теплом матері.
Теплокровні тварини захищались не лише від переохолодження, а й від перегрівання. Цьому сприяли потові залози, що виділяли на поверхню рідину. Її випаровування охолоджувало тіло. Якщо малята відчували спрагу, вони могли злизувати піт матері, особливо на череві.
Малята росли швидше, якщо у виділеннях потових залоз були поживні речовини. Коли частина розміщених на череві потових залоз перетворилася на молочні, звірозубі рептилії «стали» ссавцями. Ці еволюційні перетворення відбувалися, звичайно, впродовж тривалого часу.
Перші представники класу Ссавці виникли у тріасовий період. Тепер на Землі живе близько 4000 видів звірів. У кайнозойську еру вони стали панівною групою хребетних.
Упродовж юрського та крейдяного періодів (двох третин своєї історії) ссавці були дрібними й непомітними істотами. Оскільки вони були теплокровними, їм доводилося витрачати багато енергії для підтримання температури тіла. Вони мали вживати їжу, за яку вони не конкурували з рептиліями. Це привело до вдосконалення зубів і щелеп. Життя перших звірів було повним небезпеки, особливо в дитячому віці, і тому турбота про потомство набула особливого значення. Це сприяло розвитку головного мозку, умовнорефлекторної діяльності, чим обумовлена складна поведінка цих тварин.
Від ссавців юрського періоду незалежно виникли дві сучасні групи – підкласи Яйцекладні та Живородні.