

Физиология мышечного сокращения Морфо-функциональная характеристика мышечной ткани
Мышечная ткань специализируется на функции сокращения и является своеобразным хемомеханическим преобразователем энергии. Структурно-функциональной ее единицей является мышечное волокно, имеющее, как правило, веретеновидную форму. В зависимости от особенностей строения мышечных волокон в животном организме выделяют два типа мышечных тканей:
поперечно-полосатая (или исчерченная);
гладкая (неисчерченная).
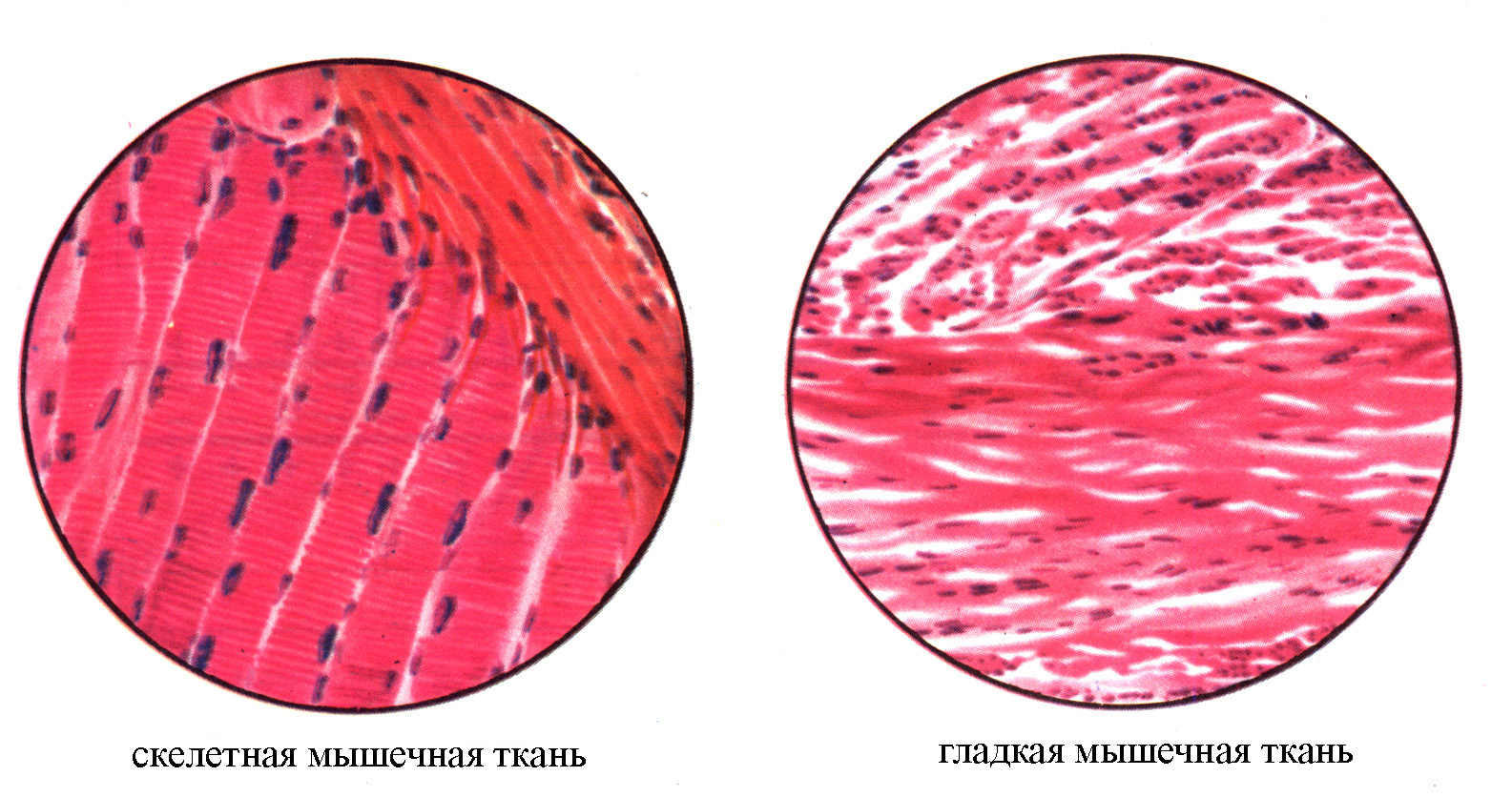
Отличительной особенностью поперечно-полосатой мышечной ткани служит наличие поперечной исчерченности ее мышечных волокон, обусловленное чередованием темных (анизотропных) и светлых (изотропных) участков (дисков) в сократительном аппарате волокна, представленном миофибриллами. Сократительный аппарат гладкомышечных волокон представлен миофиламентами (составными компонентами миофибрилл поперечно-полосатой мышечной ткани), не группирующимися в миофибриллы, в связи с чем при световой микроскопии этих волокон поперечная исчерченность отсутствует.
Поперечно-полосатая мышечная ткань, в свою очередь, представлена двумя разновидностями, характеризующимися определенными морфологическими и электрофизиологическими особенностями:
скелетной мышечной тканью;
с
 ердечной
мышечной тканью.
ердечной
мышечной тканью.
Рис. 16. Микрофотографии гладкой и скелетной мышечной ткани. Ув.: 400 (по Коробкову А.В., Чесноковой С.А., 1987).
Скелетная мышечная ткань входит в состав опорно-двигательного аппарата, составляя активную его часть (скелетные мышцы), образует язык, мышечную оболочку глотки, верхней трети пищевода и анальный сфинктер прямой кишки. Скелетная (соматическая) мускулатура опорно-двигательной системы, в свою очередь, обеспечивает передвижение организма в пространстве, перемещение частей тела друг относительно друга и поддержание позы. Кроме того, скелетные мышцы вносят существенный вклад в общий уровень теплопродукции организма, что определяет их роль в процессах терморегуляции. Наконец, сокращение некоторых скелетных мышц (наружные и внутренние межреберные, диафрагма, мышцы брюшного пресса и ряд других) сопровождается изменением объема грудной клетки, в связи с чем их называют дыхательной мускулатурой, во многом обеспечивающей внешнее дыхание организма. Скелетная мышечная ткань языка обеспечивает его движения, как при глотании, так и в процессе речи, а скелетная мышечная ткань глотки и начальной трети пищевода – произвольный характер проглатывания пищи.
Сердечная мышечная ткань формирует миокард (мышечную оболочку) сердца, периодическое ритмическое сокращение которого обеспечивает нагнетание крови в сосудистое русло и ее постоянную циркуляцию.
Гладкая (неисчерченная) мышечная ткань входит в состав большинства внутренних органов, как полых (образует мышечный слой их слизистых оболочек, а также собственно мышечную оболочку), так и неполых (например, в железах внешней секреции встречается в стенках крупных протоков), сосудов (за исключением капилляров), кожи. Наличие гладкомышечной ткани в составе большинства внутренних органов во многом обуславливает их функцию. Так, гладкомышечная ткань кишечника делает возможным различные его движения, необходимые для продвижения химуса (пищевого комка, подвергающегося действию пищеварительных соков) вдоль кишечника, всасывания продуктов расщепления (гидролиза) белков, жиров, углеводов и т.д. Гладкомышечная ткань стенок сосудов обуславливает их определенный тонус (степень сужения сосуда), а, следовательно, определенную величину периферического сосудистого сопротивления и артериального давления. Гладкомышечная ткань сфинктеров мочевого и желчного пузыря препятствует произвольному выходу их содержимого наружу и способствует его накоплению в этих органах.
Структурно-функциональной единицей мышечной ткани является мышечное волокно, которое для каждой ее разновидности характеризуется определенными морфологическими и электрофизиологическими особенностями. Так, скелетное мышечное волокно – многоядерное симпластическое образование (симпласт – результат слияния большого количества клеток предшественников), формирующееся в эмбриогенезе путем слияния клеток-предшественниц миобластов. Оно содержит до 100 ядер, диаметр скелетных мышечных волокон находится в пределах от 10 до 100 мкм, а длина от нескольких миллиметров (некоторые мышцы среднего уха) до нескольких сантиметров (иногда до 30 см). Волокно покрыто плазматической мембраной, которая вместе с окружающей его базальной мембраной составляет сарколемму (от греч. саркос – мясо). Непосредственно под сарколеммой (по периферии волокна) расположены многочисленные ядра скелетного мышечного волокна. Сократительный аппарат представлен миофибриллами, занимающими центральное положение в волокне и имеющими поперечную исчерченность. Причем миофибриллы ориентированы параллельно друг другу и параллельно продольной оси волокна.
Скелетная мышца образована совокупностью скелетных мышечных волокон и окружена собственной соединительнотканной оболочкой, называемой эпимизием. Внутри скелетной мышцы проходят прослойки плотной волокнистой соединительной ткани, отделяющие пучки скелетных мышечных волокон друг от друга и носящие название перимизия. Каждое в отдельности скелетное мышечное волокно окружено собственной прослойкой рыхлой волокнистой соединительной ткани, содержащей большое количество кровеносных капилляров, называемой эндомизием и обеспечивающей питание скелетного мышечного волокна. Кроме скелетной мышечной и волокнистой соединительной тканей в составе каждой мышцы имеются элементы нервной ткани, представленные чувствительными и двигательными нервными волокнами, проходящими в составе соединительнотканных прослоек, и чувствительными и двигательными нервными окончаниями. Чувствительные нервные волокна, проникающие в мышцу, оплетают интрафузальные мышечные волокна, образуя рецепторы растяжения мышцы, тогда как терминали двигательных нервных волокон вступают в синаптический контакт с мышечными волокнами, как экстрафузальными, так и интрафузальными. Интрафузальные мышечные волокна входят в состав т.н. мышечных веретен – рецепторов растяжения мышцы. Каждое мышечное веретено включает от 2 до 12 интрафузальных мышечных волокон и отграничено от остальных волокон мышцы собственной соединительнотканной оболочкой. Интрафузальные мышечные волокна получают и чувствительную и эфферентную иннервацию, тогда как остальные волокна мышцы, не входящие в состав веретен и в связи с этим называемые экстрафузальными, получают только эфферентную иннервацию. Собственно рабочими волокнами мышцы, во многом обеспечивающими развитие ею силы при сокращении, являются экстрафузальные волокна, составляющие в мышце большинство. Интрафузальные же волокна, несмотря на то, что способны к сокращению, не вносят весомого вклада в развитие мышечной силы при сокращении, однако, принимают участие в образовании рецепторного аппарата мышцы и восприятии ее длины и скорости сокращения.
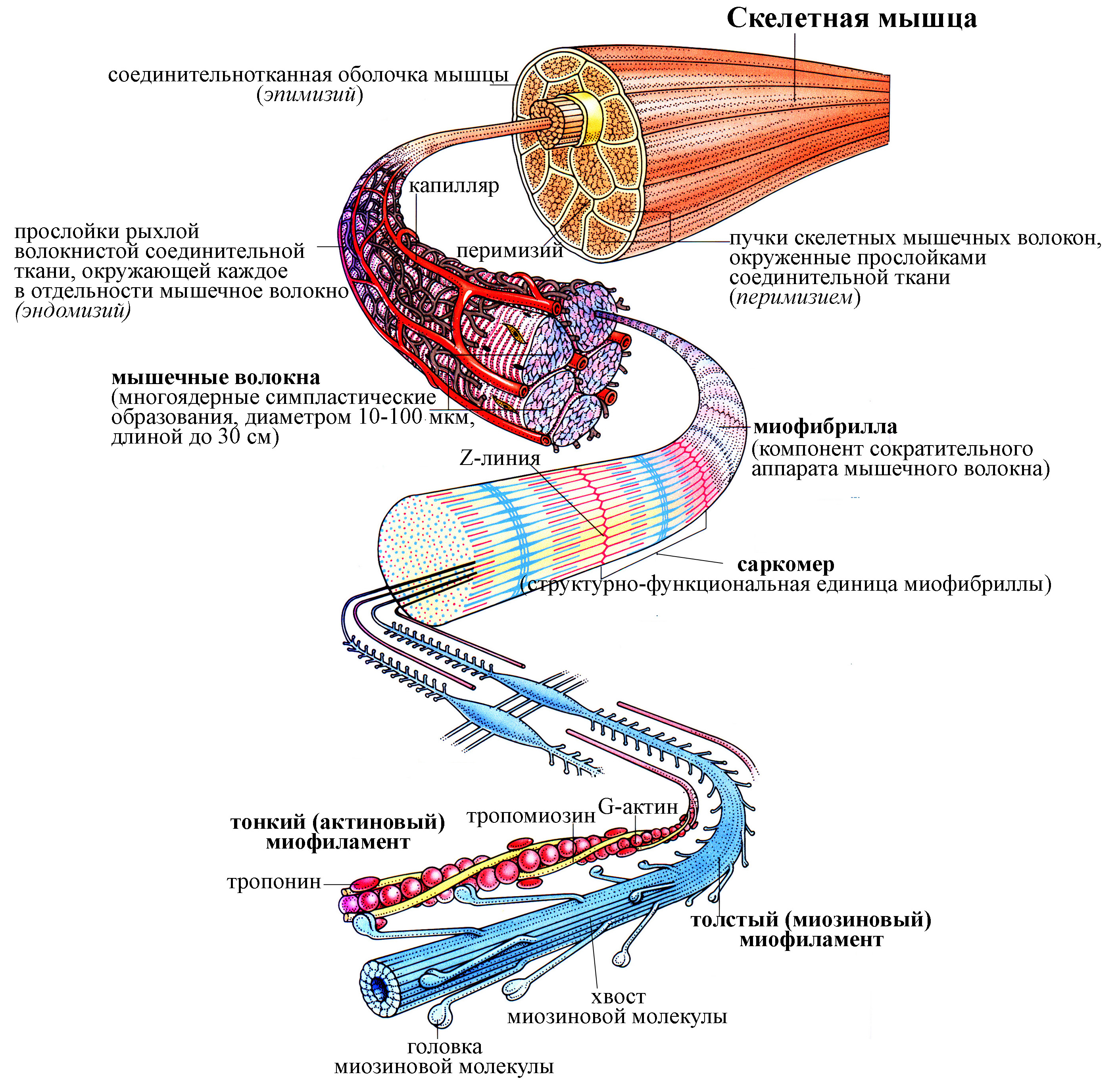
Рис. 17. Схема структурной организации скелетной мышцы, скелетных мышечных волокон и их сократительного аппарата – миофибрилл (по T. Smit, 2002).
Одной из отличительных черт скелетных мышечных волокон является хорошо развитая гладкая эндоплазматическая сеть, которая применительно к мышечной ткани называется саркоплазматической сетью (или ретикулумом, СР) и служит основным депо ионов кальция. Она представляет собой систему анастамозирующих мембранных трубочек (продольные цистерны СР), которые заканчиваются расширенными терминальными (или боковыми) цистернами. Причем такая сеть мембранных цистерн СР окружает каждую миофибриллу, а также залегает непосредственно под плазмолеммой волокна. Плазмолемма скелетного мышечного волокна образует регулярные выпячивания, направленные внутрь волокна и называемые Т-трубочками (Т-системой), с которыми контактируют с каждой стороны боковые цистерны СР. Т-трубочка вместе с окружающими ее двумя боковыми цистернами СР составляет триаду (ее роль будет рассмотрена ниже).
 А Б
А Б
В
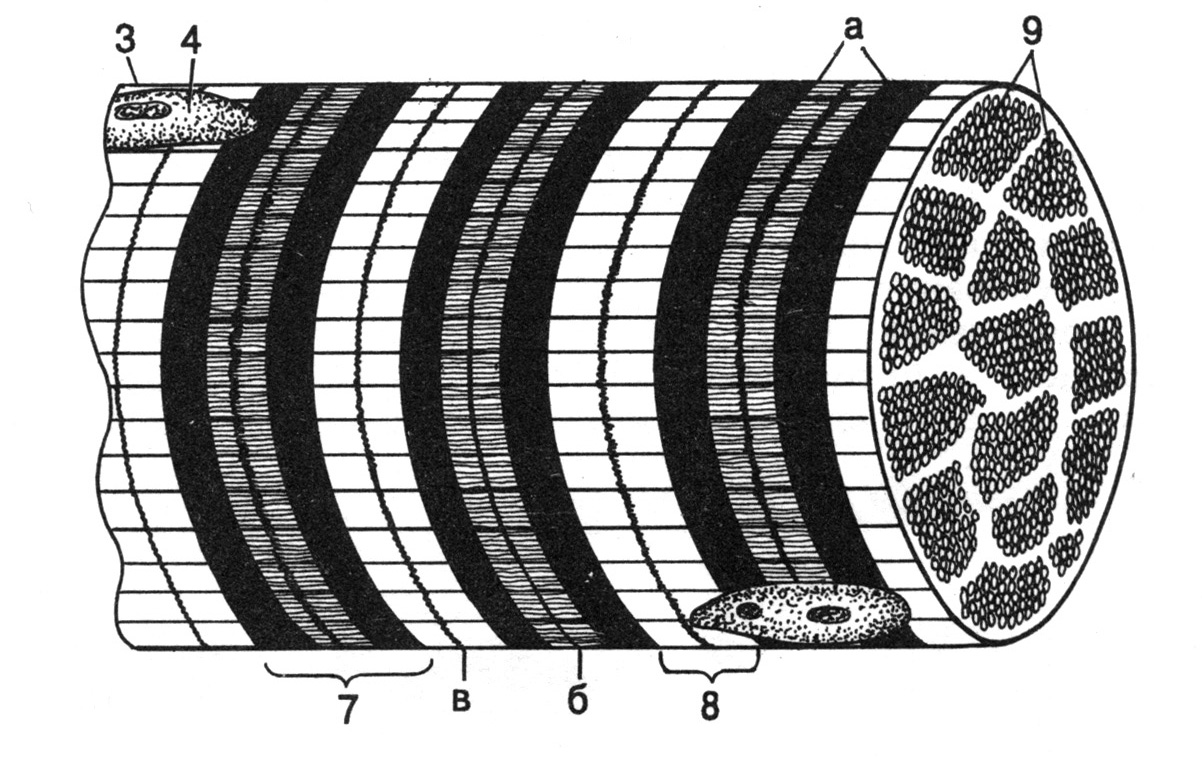
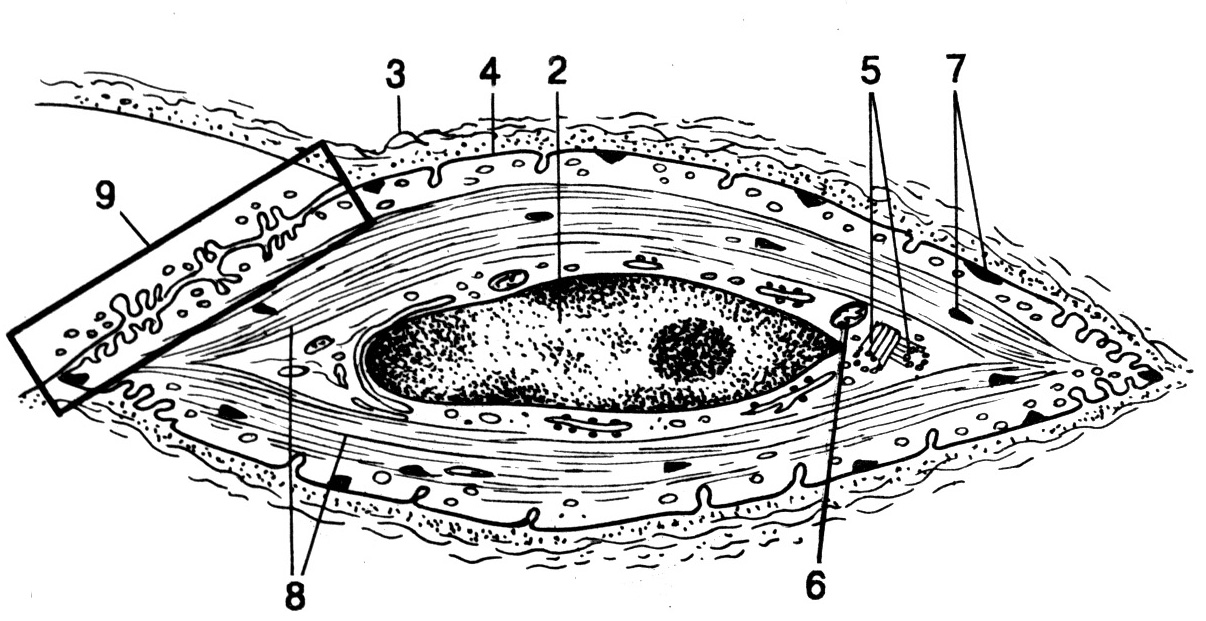
Рис. 18. Гистоструктура скелетной мышечной ткани. А и Б – схемы, В – ультраструктура (ТЭМ) (по Сидельниковой Л.П., 1994).
1 – эндомизий (прослойка соединительной ткани, окружающая каждое в отдельности волокно), 2 – ядра фиброцитов (соединительнотканных клеток), 3 – плазмолемма волокна, 4 – ядра скелетного мышечного волокна, 5 – поперечная исчерченность цитоплазмы (обусловлена чередованием темных и светлых дисков в миофибриллах), 6 – продольная исчерченность цитоплазмы (обусловлена наличием в волокне большого количества миофибрилл, расположенных параллельно друг другу), 7 – темный диск миофибриллы (диск А): а – светлая (Н-зона), б – М-линия (мезофрагма), 8 – светлый диск миофибриллы (диск I): в – Z-мембрана, 9 – поперечный срез миофибрилл.
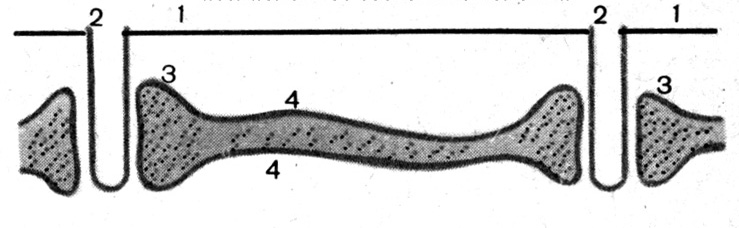
Рис. 19. Схема строения Т-трубочек и их связи с цистернами саркоплазматического ретикулума (СР) (по Косицкому Г.И., 1985).
1 – плазматическая мембрана скелетного мышечного волокна (сарколемма); 2 – Т-трубочка (регулярные выпячивания плазмолеммы волокна, направленные внутрь волокна, диаметром 0,05 мкм); 3 – боковые цистерны СР; 4 – продольные цистерны СР.
С ердечное
мышечное волокно, подобно скелетному,
является исчерченным, но, в
отличие от скелетного мышечного волокна,
представляет собой многоклеточное
образование, состоящее из совокупности
последовательно уложенных и соединенных
с помощью вставочных дисков сердечных
мышечных клеток – кардиомиоцитов.
ердечное
мышечное волокно, подобно скелетному,
является исчерченным, но, в
отличие от скелетного мышечного волокна,
представляет собой многоклеточное
образование, состоящее из совокупности
последовательно уложенных и соединенных
с помощью вставочных дисков сердечных
мышечных клеток – кардиомиоцитов.
А
Б
Рис. 20. Сердечная мышечная ткань. А – микрофотография сердечной мышечной ткани. Ув.: 400 (по Елисееву В.Г., Афанасьеву Ю.И., Котовскому Е.Ф., 1996), Б – схема строения сердечного мышечного волокна (по Волковой О.В., Елецкому Ю.К., 1996).
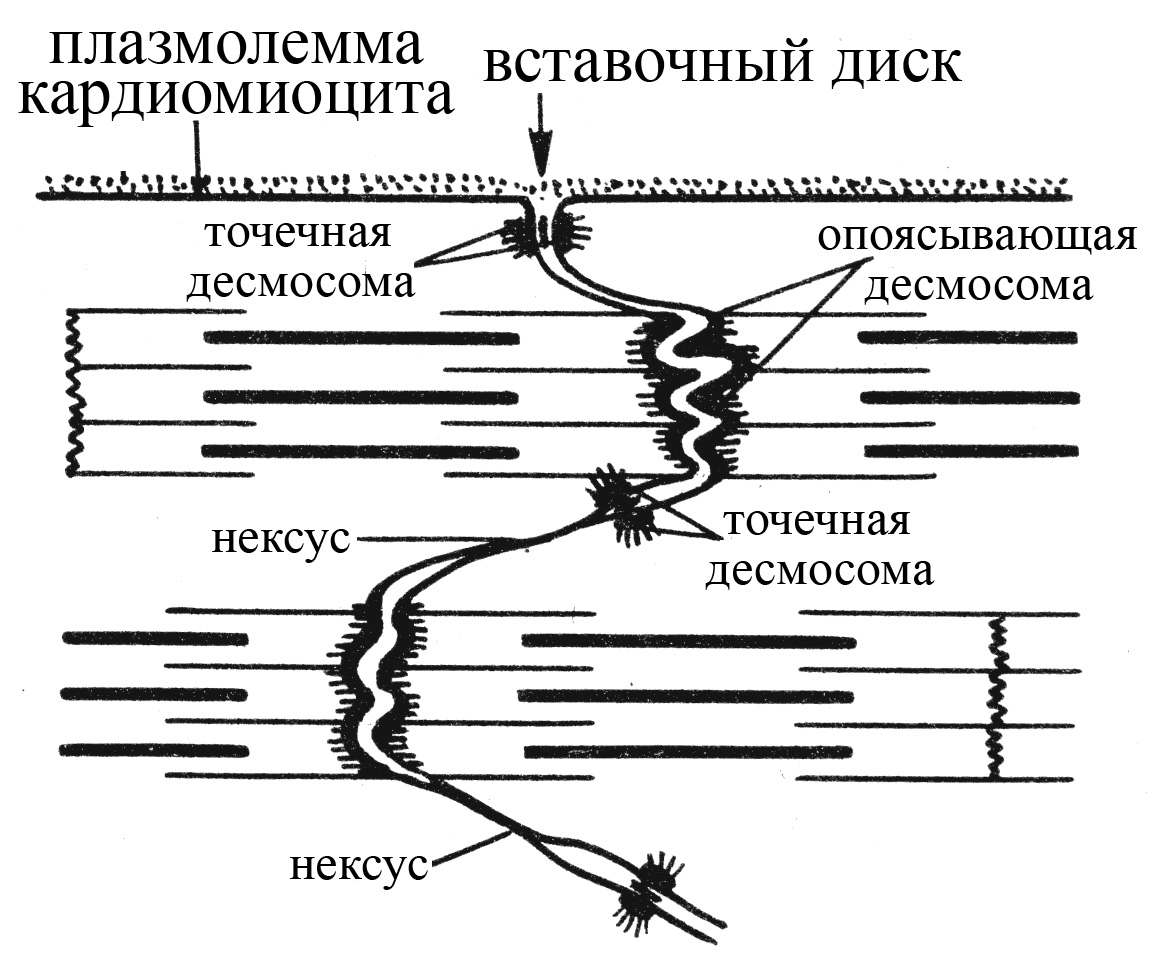
Вставочные диски представляют собой места контакта кардиомиоцитов, в области которых имеются различные типы межклеточных соединений. В частности, механическую связь между кардиомиоцитами обеспечивают точечные и опоясывающие десмосомы, а возможность обмена низкомолекулярными органическими веществами, а также проведение ионных токов – щелевые контакты или нексусы. В области точечных десмосом мембраны соседних кардиомиоцитов сближены и укреплены тонофиламентами, а в области опоясывающих десмосом – миофибриллами, концы которых зафиксированы изнутри на плазматической мембране кардиомиоцитов. Щелевые контакты или нексусы устроены по типу электрических синапсов и позволяют проводить местные ионные токи от одного кардиомиоцита на соседние. В месте образования нексуса мембраны соседних кардиомиоцитов сближаются на расстояние 2-4 нм, и между ними возникает единый канал, образованный молекулами интегрального белка коннексина. Коннексин состоит из 6 субъединиц, пронизывающих билипидный слой мембраны и окружающих собой пору диаметром 1,5 нм. Субъединицы коннексина мембраны одного кардиомиоцита располагаются строго против таковых другого, в результате чего между двумя кардиомиоцитами возникает единая, общая для них, пора (канал), через которую могут диффундировать ионы и низкомолекулярные органические вещества (АТФ, глюкоза, аминокислоты). Благодаря наличию щелевых контактов в сердечной мышечной ткани становится возможным распространение ионных токов между кардиомиоцитами без значительного их шунтирования в межклеточных щелях. Таким образом, щелевые контакты делают возможным непосредственную передачу возбуждения от возбужденного кардиомиоцита на соседние контактирующие с ним покоящиеся клетки, что не возможно в скелетной мышечной ткани между ее волокнами. Сердечная мышца, в отличие от скелетной, благодаря наличию нексусов между миокардиальными клетками, представляет собой функциональный синцитий (возбуждение даже небольшой группы кардиомиоцитов передается на соседние клетки и, в конце концов, может охватить последовательно все участки миокарда).

А Б
Рис. 21. Структурная организация вставочных дисков сердечной мышечной ткани (по Волковой О.В., Елецкому Ю.К., 1996).
А – схема строения вставочного диска, расположенного между соседними кардиомиоцитами
Б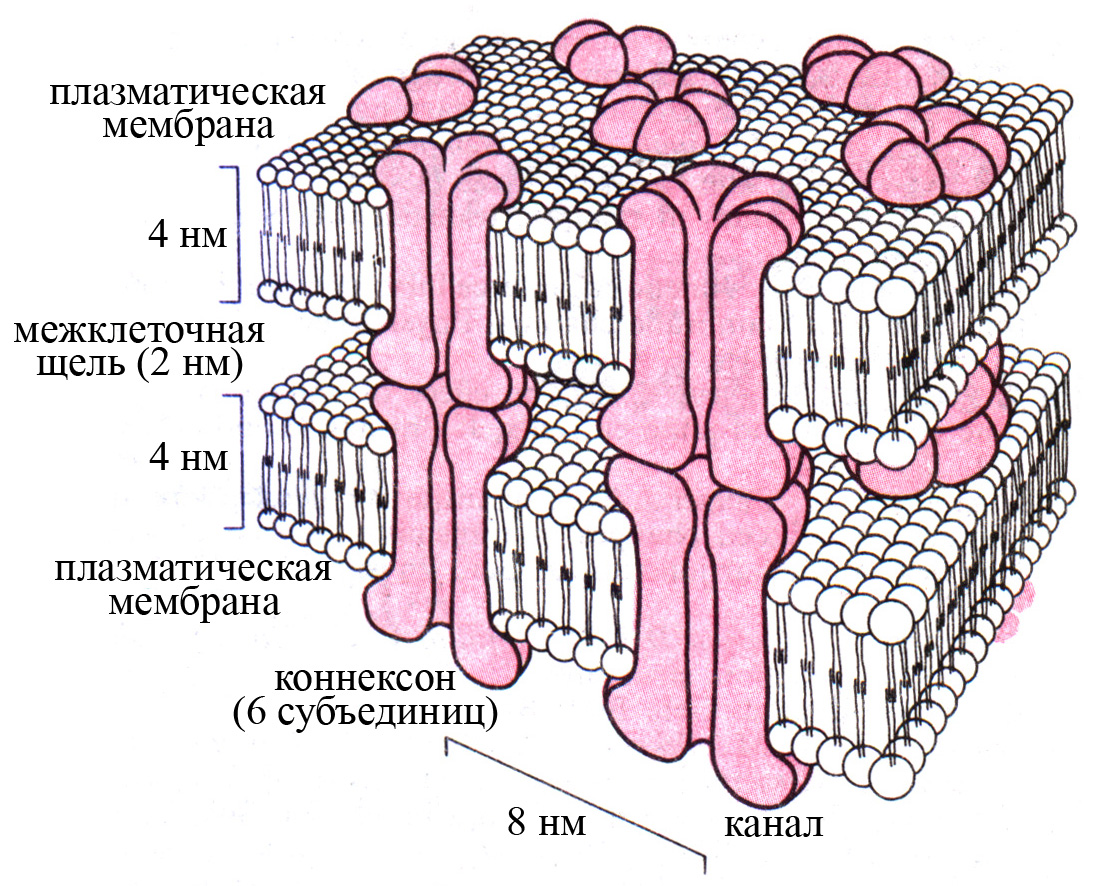 – ультраструктурная организация
вставочного диска, расположенного между
соседними кардиомиоцитами (ТЭМ). 1 –
нексус, 2 – опоясывающая десмосома, 3 –
миофибриллы, 4 – митохондрия.
– ультраструктурная организация
вставочного диска, расположенного между
соседними кардиомиоцитами (ТЭМ). 1 –
нексус, 2 – опоясывающая десмосома, 3 –
миофибриллы, 4 – митохондрия.
Рис. 21. Схема строения щелевых контактов (по Makowski L., Caspar D.L.D., Phillips W.C., 1977). В мембранах соседних кардиомиоцитов регулярно распределены каналы (коннексоны), находящиеся точно друг против друга. Внутри коннексона имеется просвет, так что каждая пара расположенных по одной линии коннексонов образует единый канал между двумя кардиомиоцитами, через посредство которого они сообщаются между собой.
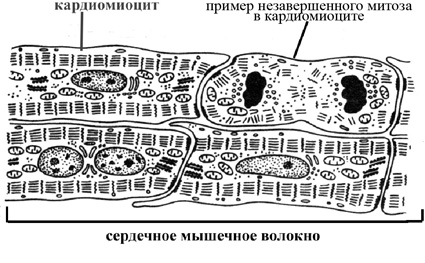
В связи с тем, что сердечное мышечное волокно представляет собой многоклеточное образование, его называют функциональным волокном. Сократительный аппарат кардиомиоцитов представлен миофибриллами, которые ориентированы параллельно продольной их оси, но, в отличие от скелетных мышечных волокон, занимают в кардиомиоците периферическое положение и зафиксированы своими концами на плазматической мембране, укрепляя десмосомоподобные контакты между сердечными мышечными клетками. Кардиомиоциты представляют собой высокодифференцированные не способные к делению в постэмбриональном периоде одно- или двуядерные клетки, характеризующиеся центральным расположением ядер. Появление двуядерных кардиомиоцитов является следствием незавершенных митозов и увеличивает способность сердечных клеток к синтезу белков и возможной их гипертрофии. Подобно скелетным мышечным волокнам, кардиомиоциты характеризуется наличием хорошо развитого саркоплазматического ретикулума и системы Т-трубочек.
Гладкое мышечное волокно – одноядерное одноклеточное образование, характеризующееся гораздо меньшими размерами по сравнению со скелетным и сердечным волокнами (диаметр – от 2 до 10 мкм, а длина от 50 до 400 мкм). Его сократительный аппарат представлен миофиламентами (двух типов: актиновыми и миозиновыми), не образующими миофибрилл и невидимыми при световой микроскопии. Саркоплазматический ретикулум в гладком мышечном волокне развит слабо, также как слабо развита и Т-система, представляющая собой в данном случае нерегулярные выпячивания плазмолеммы внутрь волокна, имеющие вид небольших пузырьков и называемых кавеолами. Между гладкомышечными клетками, подобно сердечной мышечной ткани, имеются щелевые контакты (нексусы), делающие возможным передачу возбуждения с одной гладкомышечной клетки на соседние, в связи с чем гладкая мышечная ткань представляет собой функциональный синцитий.
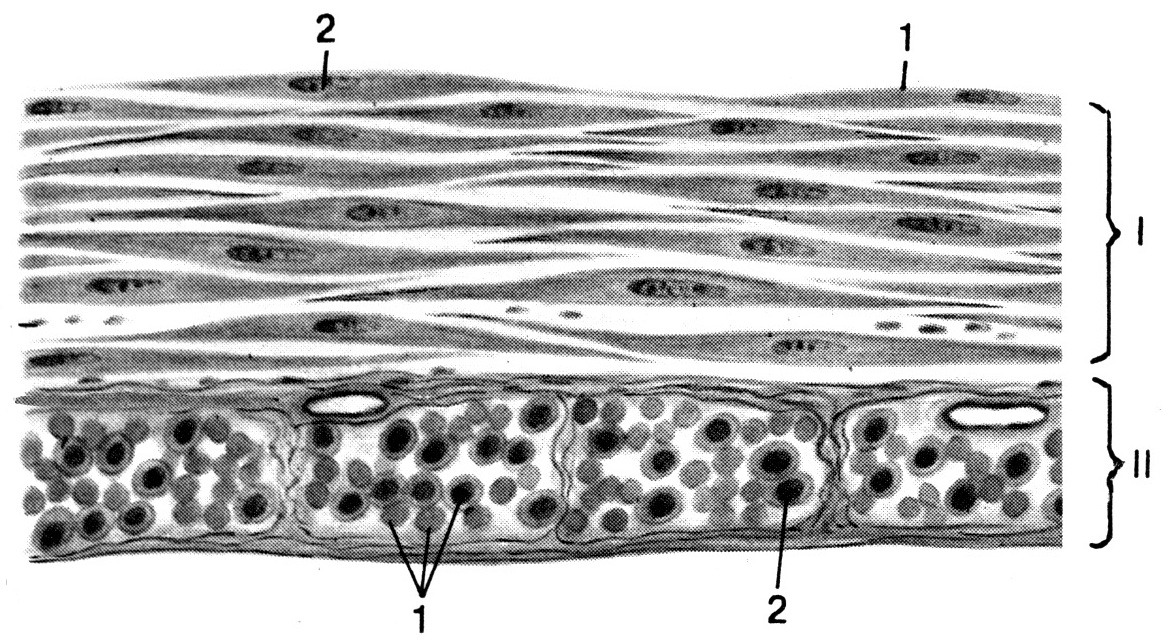
А Б
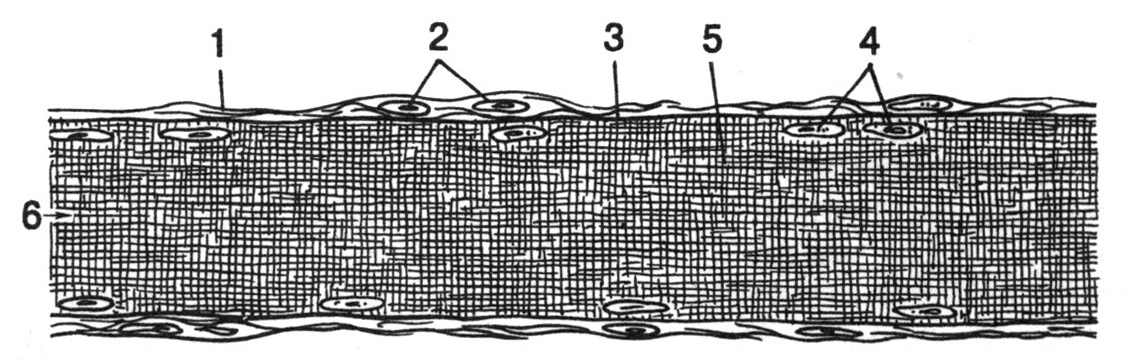
Рис. 22. Гладкая мышечная ткань (по Волковой О.В., Елецкому Ю.К., 1996). А – микрофотография препарата, Б – ультраструктура гладкого миоцита (схема). I – продольный разрез гладкомышечных волокон, II – поперечный разрез гладкомышечных волокон. 1 – гладкий миоцит (гладкомышечное волокно), 2 – ядро миоцита, 3 – эластические волокна окружающей миоцит соединительной ткани, 4 – плазмолемма миоцита, 5 – клеточный центр, 6 – митохондрии, 7 – плотные тельца (субстрат для прикрепления актиновых филаментов), 8 – миофиламенты (разбросаны диффузно, не образуют миофибрилл), 9 – мио-миоцитарный контакт, устроенный по типу электрического синапса (нексус), позволяющий непосредственно передавать возбуждение с одного гладкомышечного волокна (миоцита) на другое.
Следовательно, скелетное мышечное волокно – это многоядерное симпластическое образование, сердечное – многоклеточное многоядерное образование, а гладкое – одноклеточная одноядерная структура. Сократительный аппарат скелетного и сердечного мышечных волокон представлен миофибриллами, тогда как гладкого – их составными компонентами – миофиламентами (или протофибриллами), не образующими миофибрилл. Саркоплазматический ретикулум и Т-система в скелетном и сердечном мышечных волокнах хорошо развиты, в гладком – напротив, очень слабо.