

4. Клітинний рівень організації біологічних систем
Клітина — основна структурно-функціональна одиниця всіх живих організмів, для якої характерний індивідуальний метаболізм та здатність до відтворення. Прокаріотична клітина не має ядра (бактерії та синьо-зелені водорості), еукаріотична клітина має ядро (рослини, гриби, тварини).
Всі живі організми складаються з однієї або більшої кількості клітин. Відповідно, вони поділяються на одноклітинні, колоніальні та багатоклітинні. До одноклітинних належать бактерії, археї, деякі водорості, гриби а також найпростіші.
Розмір клітин обмежується співвідношенням поверхні до об’єму, тому клітини еукаріотів мають відносно невеликі розміри – у середньому в діаметрі близько 100 мкм. Крім того, певному розміру ядра відповідає певний об’єм цитоплазми: V ядра / V цитоплазми = const (постійне для даного типу клітин). Тiло людини складається з 10 трильйонiв клітин, форма, будова, тривалість їх життя залежать від функцій. Більшість клітин не округлі, а багатокутні, веретеновидні, мають вирости та інвагінації на поверхні. М’язові волокна можуть потовщуватись при інтенсивних тренуваннях внаслідок злиття одноядерних клітин-попередниць. Нервові клітини та клітини печінки при навантаженнях збільшують кількість хромосом і стають поліплоїдними, таким чином, збільшується об’єм ядра і об’єм цитоплазми.
Таблиця 1
Основні особливості організації клітин різних груп організмів
Органела |
Тварини |
Рослини |
Гриби |
Бактерії |
Клітинна стінка |
- |
+ |
+ |
+ |
Ядро |
+ |
+ |
+ |
- |
Мітохондрії |
+ |
+ |
+ |
- |
Пластиди |
- |
+ |
+ |
- |
Травні вакуолі |
+ |
- |
- |
- |
Скоротливі вакуолі |
+ |
+ |
- |
- |
Вакуолі |
- |
+ |
- |
|
Лізосоми |
+ |
+ |
+ |
- |
Апарат Гольджі |
+ |
+ |
+ |
- |
ЕПР |
+ |
+ |
+ |
- |
Центріолі |
+ |
- / + |
- |
- |
4.1. Поверхневий апарат клітини
Плазматична мембрана (плазмалема) оточує клітину та визначає її розміри і форму, являючи собою ліпідно-білковий комплекс з вуглеводними включеннями.
Рідинно-мозаїчна (рідинно-кристалична) модель будови мембрани: фосфоліпідний бішар є двовимірною рідиною, в якій вільно плавають білкові молекули, утворюючи рухому «мозаїку». Мембранні молекули змінюють розташування та конформацію в залежності від оточуючих хімічних факторів: води, речовин та іонів. Зовнішні мембрани взаємодіють з міжклітинною речовиною, а внутрішні утворюють єдину систему з цитоплазмою. Зовнішня і внутрішня сторони мембрани відрізняються по фосфоліпідному і білковому складу та відповідно своїх функцій.
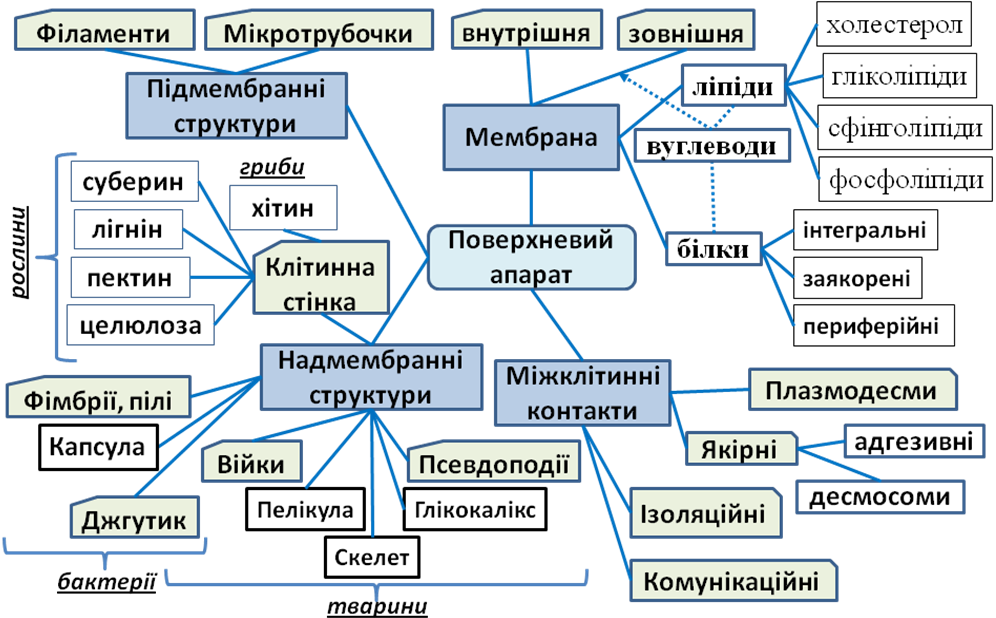
Рис. 8. Поверхневий апарат клітини
Зовнішня мембрана відмежовує внутрішній вміст клітини від навколишнього середовища, забезпечує сприйняття хімічних та фізичних подразнень, а також надходження необхідних речовин та видалення відходів.
Внутрішні мембрани ділять клітину на компартменти, кожен із яких призначений для певних метаболічних процесів (фотосинтез, гідроліз біополімерів, тощо). Крім того, деякі хімічні реакції можуть відбуватися тільки на самих мембранах, наприклад реакції світлової фази фотосинтезу або кінцевий етап аеробного окиснення. У прокаріотів внутрішні мембрани і компартменталізація відсутні, проте у деяких фотосинтезуючих та аеробних бактерій плазмалема утворює інвагінації всередину клітини, які виконують відповідні метаболічні функції.
Загальна будова мембран. Молекули фосфоліпідів утворюють безперервний подвійний шар завтовшки ≈ 4-7 нм, в якому гідрофільні «головки» (залишок фосфатної кислоти і холіну, серину, етаноламіну або іншої полярної сполуки) орієнтовані назовні у полярний міжклітинний розчин, а два гідрофобні «хвости» (залишки жирних кислот) спрямовані всередину один до одного. Крім фософоліпідів до складу мембран входять також інші типи ліпідів, такі як гліколіпіди, сфінголіпіди та холестерол. Ліпіди можуть переміщуватися в бічному напрямку, змінюючи своє положення до мільйона разів за секунду, а іноді можуть перестрибувати з одного шару в інший. Холестерол забезпечує стабільність мембран, вбудовуючись між молекулами фосфоліпідів. Вмонтовані в мембрану білки – додатковий механізм стабілізації подвійного шару ліпідів. Вміст білків у клітинній мембрані може коливатись від 18% (аксони) до 75% (тилакоїди). Мембранні білки бувають гідрофільними, амфіфільними, фібрілярними (у вигляді ниток) і глобулярними.
За розміщенням всі мембранні білки поділяються на 3 основні групи:
трансмембранні (інтегральні) білки мають у своєму складі гідрофобну частину, яка пронизує ліпідний бішар (всі іонні канали та більшість клітинних рецепторів);
білки заякорені в мембрані за допомогою ліпідів — ковалентно з'єднані з неполярною сполукою (найчастіше з глікозилфосфатидилінозитолом), яка входить у гідрофобну частину ліпідного бішару, і таким чином утримує білок на поверхні мембрани (G-білки);
периферійні білки нековалентно взаємодіють із іншими білками або гідрофільними головками фосфоліпідів, утримуючись поблизу поверхні мембрани тільки тимчасово (деякі ферменти).
Вуглеводи вмонтовуються у мембрану за 2 правилами:
завжди розташовуються на нецитоплазматичній поверхні мембрани;
приєднані до білків (93%) або ліпідів (7%).
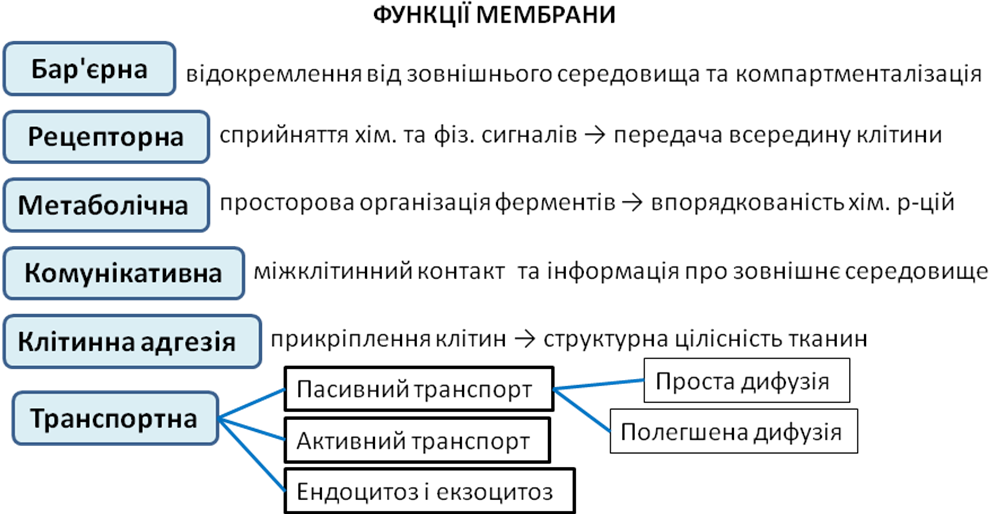
Рис. 9. Функції мембрани
Функції мембрани
Бар’єрна – відокремлює клітину від зовнішнього середовища, а також окремі компоненти клітин один від одного, забезпечуючи асиметричний розподіл речовин між внутрішньоклітинним і позаклітинним середовищами. Мембрани характеризуються вибірковою проникністю: через них можуть проходити неполярні молекули (наприклад кисень, вуглекислий газ) та гідрофобні речовини (стероїдні гормони, жирні кислоти), але вони непроникні для великих полярних та заряджених молекул (амінокислот, моносахаридів, іонів).
Транспортна — вибіркове надходження потрібних речовин до клітини та виведення з неї відходів. Розрізняють наступні види клітинного транспорту:
Пасивний транспорт — рух речовин через мембрану по градієнту концентрації (від зони з більшою концентрацією до ділянки з меншою концентрацією) без затрат енергії.
Проста дифузія — рух речовин (гідрофобні молекули або розчинені гази) безпосередньо через ліпідний бішар; різновидом простої дифузії є осмос.
Полегшена дифузія — рух молекул через спеціальні білкові канали або по градієнту концентрації за допомогою білків-переносників (іони неорганічних речовин). Ферменти транслокази і пермеази зв’язують своїм активним центром молекулу з одного боку мембрани і переносять її крізь гідрофобний шар, при цьому конформація білка-переносника може змінюватись, внаслідок чого в мембрані відкривається спеціальний гідрофільний канал.
Активний транспорт — рух речовин через мембрану проти градієнту концентрації, що відбувається з витрачанням енергії і здійснюється за допомогою спеціальних білків насосів. Розрізняють первинний активний транспорт, для якого використовується енергія гідролізу АТФ (натрій-калієвий насос) та вторинний активний транспорт, коли для переміщення однієї речовини проти градієнту концентрації інша транспортується за градієнтом (всмоктування глюкози клітинами тонкого кишечника).
Ендоцитоз і екзоцитоз — енергозатратні процеси транспортування речовин і часточок у клітину (ендоцитоз) або із клітини (екзоцитоз) за участю мембранних везикул (пухирців).
Рецепторна — за допомогою рецепторів (найчастіше глікопротеїнів), розташованих на мембранній поверхні, сприймаються та підсилюються хімічні та фізичні сигнали; таким чином здійснюється контакт сусідніх клітин між собою та позаклітинною речовиною, клітини багатоклітинного організму розпізнають одна одну. Рецепторні білки мембрани здатні специфічно зв’язуватись з гормонами, нейромедіаторами, цитокінінами.
Метаболічна — служить каркасом для просторової організації впорядкованих ферментативних процесів: інтегровані в мембрану ферменти можуть бути організовані у комплекси для здійснення низки послідовних метаболічних реакцій. Внутрішні мембрани розділяють клітину на відсіки (компатрменти), в кожному з яких проходять певні хімічні реакції, необхідні для виконання відповідних функцій (реакції світлової фази фотосинтезу та електронтранспортного ланцюга мітохондрій). Мембрана мітохондрій містить АТФ-синтетази – білки переносники електронів, які забезпечують синтез АТФ. На зовнішній мембрані містяться АТФ-ази, що рощеплюють АТФ, тому що для транспорту речовин витрачається енергія (24 ферменти на мембрані клітин печінки).
Клітинна адгезія – забезпечує прикріплення клітин багатоклітинного організму одна до одної або до позаклітинного матриксу (кадгерини - білки мембран тварин). Таким чином забезпечується структурна цілісність тканин тваринного організму і контакт з мікрооточенням.
Клітинна стінка оточує клітину, захищає її, надає сталої форми та запобігає осмотичному руйнуванню. У грам-позитивних бактерій (Staphylococcus, Bacillus, Lactobacillus) вона складається переважно з пептидоглікану муреїну, у грам-негативних (Salmonella, Escherichia, Azotobacter) – має додаткову зовнішню мембрану з фосфоліпідів, у архей в основному побудована з білків та полісахаридів.
Капсула — слизова оболонка бактерій, яка побудована переважно з вуглеводів, білків та уронових кислот; захищає від висихання та знищення, утримує клітини в колоніях, сприяє прикріпленню до субстратів.
Пілі або ворсинки — тонкі волоскоподібні вирости, що використовуються для прикріплення та утримання на субстраті (фімбрії у Neisseria gonorrhoeae та Helicobacter pylori), задіяні у процесі кон'югації (статеві пілі або F-пілі бактерій).
Більшість тварин здатні до руху, тому їх тканини мають бути еластичними. Проте у одноклітинних найпростіших організмів поверхневий апарат часто утворює твердий опорний шар (скелет) або пружну пелікулу.
Глікокалікс (грецьк. глікос – солодкий та каллум – товта шкіра) – надмембранний комплекс клітин, утворений вуглеводами, які знаходяться у складі глікопротеїдів та гліколіпідів мембран у тварин. Ці вуглеводи – насичена водою волокнисто-слизова маса, що може містити компоненти міжклітинної речовини або глікокаліксу сусідніх клітин