

В присутствии лактозы (положительная индукция):

В отсутствии лактозы (отрицательная индукция):
элонгация (рост цепи): скорость процесса в клетках E. coli in vivo 45-50 нуклеотидов в 1с при 37 0С;
терминация вызывается стоп-сигналом (определенной последовательностью нуклеотидов в ДНК, другое название – терминатор). Структура терминаторов полностью не выяснена: предполагают наличие длинных блоков типа: ТТТ…
ААА…
Особенности транскрипции у эукариот: если у прокариот транскрипция и трансляция сопряжены, то у эукариот они разобщены во времени и пространстве. Процесс этот изучен еще недостаточно.
Особенности регуляции транскрипции у эукариот:
существует три типа РНК-полимераз.
РНК-полимеразы
I II III
считывает гены считывает основную считывают гены
рРНК 18 S часть структурных генов, основных м яРНК,
28 S кодирующих белки, и тРНК, 5S рРНК
5.8 S гены, кодирующие
некоторые м яРНК
(малые ядерные)
2) РНК-полимеразы не могут самостоятельно инициировать транскрипцию, требуется большое количество белков – факторов транскрипции;
3) на скорость транскрипции эукариот могут влиять регуляторные белки, связывающиеся с ДНК за тысячи п. н. от промотора. Эти участки ДНК называются энхансерными (англ. enhance – усиливать). Механизм действия: ДНК образует петлю таким образом, что промотор и энхансер находятся друг напротив друга, а присоединяющийся к энхансеру специфический регуляторный белок является проводником сигнала на промотор:

5.4.4. Трансляция
Трансляция – это перевод информации, заложенной в последовательности нуклеотидных мРНК, в последовательность аминокислотных остатков полипептидных цепей.
Этапы:
1) активирование аминокислот под действием фермента аминоацил-тРНК-синтетазы и присоединение их к соответствующим тРНК;
2) собственно трансляция (сборка полипептидной цепи на рибосомах). Включает в себя: 1) инициацию – полипептидная цепь начинает строиться с N-конца и завершается С-концом. Инициирующий кодон в начале цепи АУГ (иногда ГУГ); 2) элонгацию (рост полипептидной цепи) – на рибосоме вновь поступившая аминокислота своим NH2-концом взаимодействует с СООН-концом предыдущей аминокислоты. Процесс протекает под действием фермента – пептидилтрансферазы. Элонгация заканчивается, когда в рибосому на мРНК приходят стоп-сигналы (УАА, УАГ, УГА); 3) терминацию.
У эукариот синтез белка включает 3 этапа:
- транскрипция,
- сплайсинг (см. выше),
- трансляция.
5.4.5. Особенности строения генов эукариот
У эукариот гены имеют мозаичную (экзон-интронную) структуру (экзоны кодируют белки, интроны не имеют прямого отношения к белковой последовательности, промежуточные участки). [Этот научно установленный факт опровергает универсальность принципа колинеарности генов – «что верно для E. coli, то верно и для слона»]. Но это открытие позволило дополнить этапы синтеза белка у эукариот еще одним процессом – сплайсингом (сплайсинг – созревание мРНК, когда вырезаются интроны из первоначальной копии, а экзоны соединяются между собой (см. схему)).
Сплайсинг


обычный |
альтернативный |
Э1 + Э2 + Э3 + … |
Э2 + Э4 + Э6 + Э1 + Э3 + Э5 + … |
Экзоны составляют 100 – 600 п.н.; интроны – несколько десятков тысяч п. н. Например, ген овальбумина курицы состоит из 7000 п. н. Из них на экзоны приходится 1872 п. н., а остальные 75 % приходятся на интроны.

В процессе созревания молодой мРНК (первичного РНК-транскрипта) на 5’- и 3’-концах возникают дополнения:
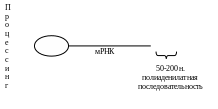
3’
5’ КЭП АААА…
Зачем нужны интроны?
– для осуществления альтернативного сплайсинга;
– внутри интронов могут быть спрятаны гены маленьких белков (например, у дрозофилы в гене Gart (47 тыс. п. н.) внутри большого интронного участка находится ген кутикулярного белка (0,9 тыс.п. н.);
– большая протяженность интронов обеспечивает защиту от мутаций;
– интроны – это бывшие экзоны, которые утратили в процессе эволюции свою роль.