

Гомотипические и гетеротипические реакции
Клементс и Шелфорд (1939) взаимодействиям между различными организмами, населяющими данную среду, дали название коакций. Коакции подразделили на два типа.
Гомотипические реакции, или взаимодействия между особями одного и того же вида. Реакции этого типа весьма разнообразны. Основные из них — групповой и массовый эффекты, внутривидовая конкуренция.
Гетеротипические реакции, т.е. взаимоотношения между особями разных видов. Выделяют различные формы биотических отношений. Воздействие одного вида на другой может быть положительным, отрицательным и нейтральным. При этом возможны разные комбинации типов воздействия.
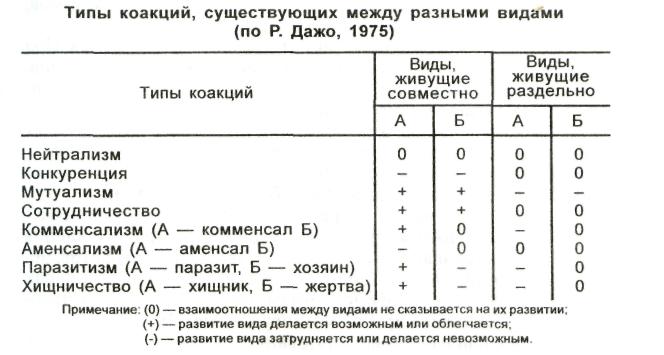
Нейтрализм — оба вида независимы и не оказывают друг на друга никакого влияния.
Конкуренция — каждый из видов оказывает на другой неблагоприятное действие. Виды конкурируют в поисках пищи, укрытий, мест кладки яиц и т. п. Оба вида называют конкурирующими.
Мутуализм — симбиотические взаимоотношения, когда оба сожительствующих вида извлекают взаимную пользу.
Сотрудничество — оба вида образуют сообщество. Оно не является обязательным, так как каждый вид может существовать отдельно, изолированно, но жизнь в сообществе им обоим приносит пользу.
Комменсализм — взаимоотношения видов, при которых один из партнеров получает пользу, не нанося ущерб другому.
Аменсализм — тип межвидовых взаимоотношений, при котором в совместной среде один вид подавляет существование другого вида, не испытывая противодействия.
Паразитизм — это форма взаимоотношений между видами, при которой организмы одного вида (паразита, потребителя) живут за счет питательных веществ или тканей организма другого вида (хозяина) в течение определенного времени.
Хищничество — такой тип взаимоотношений, при котором представители одного вида поедают (уничтожают) представителей другого, т. е. организмы одного вида служат пищей для другого.
Основные типы коакций приведены в табл. 6.1.
Среди взаимополезных взаимосвязей (+, +) среди видов (популяций) помимо мутуализма выделяют симбиоз и протокооперацию.
Межвидовые взаимоотношения
Они могут быть безразличными, вредными или полезными для партнеров. При нейтрализме оба вида живут на одной территории, не вступая в отношения друг с другом, например дятлы неподалеку от дроздов в буковом лесу или гидроидные полипы на раковине моллюска. Может существовать конкуренция за одинаковую пищу или жизненное пространство, например, между двумя видами воробьиных — славкой и соловьем.
При совместном содержании в культуре Paramaecium caudatum несколько быстрее вытесняется растущей популяцией P.aurelia, так как последняя выедает бактерий — пищу, необходимую и первому виду. Однако P.aurelia, питающаяся в поверхностной бактериальной пленке, не конкурирует с P. bursaria, поедающей микроорганизмы, которые опускаются на дно.
Мутуализм приносит выгоду обоим партнерам — при симбиозе жизненно важную, при протокооперации не очень значительную. Так, жвачные животные и микроорганизмы их рубца не могут существовать друг без друга, а гидра, напротив, может жить без водоросли хлореллы, как и та без нее.
Нередко польза и вред бывают односторонними. Для льва безразлично, поедают ли грифы и шакалы остатки его пищи (комменсализм); для жуков-навозников несущественно, что в полете они переносят нематод-копрофагов к новым навозным кучам — их субстрату.
Таблица 1
Гомотипические реакции. Групповой эффект проявляется у многих видов, которые могут нормально размножаться и выживать только в том случае, если представлены достаточно крупными популяциями. Например, для выживания африканских слонов стадо должно состоять не менее чем из 25 особей, а стадо северного оленя должно насчитывать не менее 300—400 голов. «Принцип минимального размера популяции» объясняет, почему нельзя спасти виды, которые стали слишком редкими. Известно, что белый журавль в Северной Америке, несмотря на все усилия, которые предпринимались в течение многих лет, насчитывает в настоящее время не более 30—50 особей.
При совместной жизни облегчаются поиски пищи, борьба с врагами. Групповой эффект более отчетливо проявляется при наличии у животных фазности, т. е. существование вида одновременно в двух фазах: одиночных особей и особей, объединенных в стада. Так, классическим примером фазности является саранча. Наличие фаз установлено у различных видов бабочек, жуков, тлей, таракана-прусака, сверчков, сеноедов и др. Во всех случаях наблюдаются значительные изменения в плодовитости, скорости развития, а нередко в морфологических и физиологических особенностях животных.
Массовый эффект. Этот термин, предложенный Грассе, обозначает эффект, вызванный перенаселением среды. Между групповым и массовым эффектами существуют в большинстве случаев переходы, но, как правило, массовый эффект влечет за собой вредные для животных последствия, в то время как групповой эффект на них воздействует благоприятно. Один из примеров массового эффекта дают исследования, проведенные Мак Наганом и Данном (Balachowsky, 1963) на амбарном долгоносике Sitophilus (Calanda) oryzae. У этого вида число яиц, откладываемых ежедневно, достигает максимума, когда отношение числа особей (сумма самцов и самок) к числу зерен становится равным 1/200. Размер кладки уменьшается, если на самку приходится меньшее число зерен. Это явление прежде всего связано с «насыщением» зерен, так как самки не решаются откладывать яйца в зерна, занятые уже другими яйцами или личинками. Оно связано и с массовым эффектом, поскольку насекомые мешают друг другу, что и тормозит кладку. Подобные явления, вызванные массовым эффектом, называют самоограничением.
Внутривидовая конкуренция. При внутривидовой конкуренции между особями сохраняются взаимоотношения, при которых они в состоянии размножаться и обеспечивать передачу свойственных им наследственных свойств. Внутривидовая конкуренция бывает в территориальном поведении, когда животное защищает место своего гнездовья или известную площадь в его округе. Так, в период размножения птиц самец охраняет определенную территорию, на которую кроме своей самки не допускает ни одной особи своего вида. Такую же картину можно наблюдать и у многих рыб (например, колюшки).
Проявлением внутривидовой конкуренции является существование у животных социальной иерархии, которая характеризуется появлением в популяции доминирующих и подчиненных особей. Например, у майского жука личинки трехлетнего возраста подавляют личинок одно- и двухлетнего возраста. Это является причиной того, что вылет взрослых жуков наблюдается только раз в три года, тогда как у других насекомых (например, посевных щелкунов Agriotes) продолжительность личиночной стадии также составляет три года, выход имаго происходит ежегодно из-за отсутствия конкуренции между личинками.
Конкуренция между особями одного вида из-за пищи по мере увеличения плотности популяции становится более острой. В некоторых случаях внутривидовая конкуренция может приводить к дифференциации вида, к распадению его на несколько популяций, занимающих разные территории. Так, у саванной овсянки (Passerculus sandwichensis) один экологический подвид размещается на сухих холмах, другой — на прибрежных солончаках. Конкуренция нередко является причиной переселения части популяции особей из одного географического района в другой. Этим объясняют перелеты различных зерноядных птиц, так называемых узких стенофагов тайги — кедровок, свиристелей, совершающих налеты в Западную Европу, когда не хватает пищи в районах их обычного распространения.
Гетеротипические реакции
Взаимоотношения между особями разных видов, или гетеротипические реакции, проявляются в виде нейтрализма, межвидовой конкуренции, мутуализма, сотрудничества, комменсализма, паразитизма и хищничества.
Межвидовой конкуренцией называют активный поиск двумя или несколькими видами одних и тех же пищевых ресурсов среды обитания. Конкурентные взаимоотношения, как правило, возникают между видами со сходными экологическими требованиями. При совместном обитании каждый из них находится в невыгодном положении в связи с тем, что присутствие другого вида уменьшает возможности в овладении пищевыми ресурсами, убежищами и другими средствами к существованию, имеющимися в местообитании. Конкуренция относится к форме экологических отношений, отрицательно сказывающейся на взаимодействующих партнерах. Конкурентные взаимоотношения могут быть самыми различными — от прямой физической борьбы до мирного совместного существования. И вместе с тем, если два вида с одинаковыми экологическими потребностями
оказываются в одном сообществе, то обязательно один конкурент вытесняет другого. Это одно из общих экологических правил, получившее название «закон конкурентного исключения», сформулированное Г. Ф. Гаузе (1910—1986) на основании результатов опытов по содержанию двух видов инфузорий на одном и том же ограниченном питании. Как оказалось, через некоторое время в живых остаются особи только одного вида, выжившие в борьбе за пищу, так как его популяция быстрее росла и размножалась. Победителем в конкурентной борьбе оказывается тот вид, который в данной экологической обстановке имеет хотя бы небольшие преимущества перед другим, а следовательно, и большую приспособленность к условиям окружающей среды.
Конкуренция является одной из причин того, что два вида, слабо различающихся спецификой питания, поведения, образа жизни и т. д., редко сожительствуют в одном сообществе. Здесь конкуренция носит характер прямой вражды. Самая жестокая конкуренция с непредвиденными последствиями возникает, если человек вводит в сообщества виды животных без учета уже сложившихся отношений.
Чаще же конкуренция проявляется косвенно, носит незначительный характер, так как различные виды неодинаково воспринимают одни и те же факторы среды. Чем разнообразнее возможность организмов, тем менее напряженной будет конкуренция.
Хищничество и паразитизм. Хищником называют свободно живущий организм, питающийся другими животными организмами или растительной пищей. Паразит не ведет свободной жизни, и хотя бы на одной стадии своего развития он связан с поверхностью (эктопаразит) или с внутренними органами (эндопаразит) другого организма, являющегося его хозяином.
Хищник, как правило, вначале ловит жертву, убивает ее, а затем поедает. Для этого у него имеются специальные приспособления (рис. 6.3).
У жертв также исторически выработались защитные свойства в виде анатомо-морфологических, физиологических, биохимических и других особенностей. Например, выросты тела, шипы, колючки, панцири, защитная окраска, ядовитые железы, способность быстро прятаться, зарываться в рыхлый грунт, строить недоступные хищникам убежища, прибегать к сигнализации об опасности. Вследствие таких обоюдных приспособлений формируются определенные группировки организмов в виде специализированных хищников и специализированных жертв. Так, основной пищей рыси (Felix lynx) служат зайцы, а волк (Canis lupus) — типичный многоядный хищник.
Большинство плотоядных животных, как было отмечено, в состоянии убить и сразу же съесть свою жертву, так как превосходят ее размерами и силой. Паразиты же выбрали несколько другой путь: они получают от хозяина все необходимое и тем самым подрывают его здоровье, от которого зависит их собственное благополучие.
Паразитов подразделяют на две основные категории: микропаразитов и макропаразитов (рис. 6.4) К микропаразитам относятся те, которые непосредственно размножаются внутри тела хозяина (вирусы, бактерии, простейшие). Микропаразиты растут в теле хозяина, но, размножаясь, образуют особые формы, которые покидают одного хозяина, чтобы заселить другого. К макропаразитам животных относятся круглые и ленточные черви (главные макропаразиты животных), вши, блохи, клещи, грибы.
Паразитизм, таким образом, возникает в процессе тесного контакта различных видов организмов на базе пищевых и пространственных связей, характерен для многих организмов, наиболее широко распространен среди простейших, червей, в несколько меньшей степени среди членистоногих.
Среди насекомых с полным превращением есть паразитоидные виды, которые вначале ведут себя как паразиты, щадя жизненно важные органы своего хозяина, а под конец развития, съедая своего хозяина, становятся настоящими хищниками. И хищник, и паразит могут существовать за счет одного или нескольких видов. При этом различают следующие виды.
Полифаги — нападающие на большое число видов. К ним относятся многие хищные млекопитающие и насекомые. Насекомые фитофаги питаются самыми различными растениями. Так, гусеница стеблевого мотылька Pyrausta nubilabis поражает более чем 200 видов растений.
Олигофаги — живущие за счет нескольких, часто близких видов. Колорадский жук питается картофелем и другими растениями, относящимися, главным образом, к пасленовым. Солитер эхинококк паразитирует в человеке, свинье, различных плотоядных и грызунах.
Монофаги — живущие за счет только одного хозяина. Монофагия является правилом для многих паразитических насекомых, например, таких, как афелинус (Aphelinus mali) — на кровяной тле, грушевый цветоед (Anthonomus pyri) — на груше, шелковичный червь — на тутовом дереве.
Принцип совпадения. Чтобы паразит мог развиваться, необходимо совпадение во времени поражаемой стадии хозяина и агрессивной стадии паразита. Паразит развиваться не может, если нет хозяина, если он недоступен или не находится на приемлемой стадии. Этот принцип Таленхорст (Thalenhorst, 1950) назвал совпадением. Изучение совпадения, например между насекомыми-фитофагами и их растениями-хозяевами, имеет практическое значение. Примером может служить яблоневый цветоед. Установлено, что период кладки яиц цветоеда тесно связан со стадиями развития цветочных почек яблони, т. е. существует фенологическое совпадение между кладкой долгоносика и развитием его растения-хозяина. Фенологическое совпадение должно существовать и между двумя видами животных, один из которых живет за счет другого. Совпадение должно быть очень точным в случае паразита, цикл развития которого охватывает нескольких промежуточных хозяев.
Классификация взаимоотношений. Взаимоотношения организмов разнообразны. Они разделяются на горизонтальные — между организмами одного трофического уровня (как внутри вида, так и между видами) и ветрикальные - между организмами разных трофических уровней. Взаимоотношения первого рода, как правило, носят характер конкуренции, но могут на некоторых этапах жизни организмов быть мутуализмом (т. е. взаимопомощью). Взаимоотношения второго рода более разнообразны: «фитофаг - растение», «хищник — жертва» (иногда эти два типа взаимоотношений объединяют, так как растения по существу тоже жертвы), «паразит -хозяин», мутуализм, комменсализм, аменсализм..
Кроме материальных взаимоотношений (конкуренции за ресурсы или передачи вещества и энергии при хищничестве или паразитизме), возможны сигнальные (информационные) взаимоотношения. Однако эти взаимоотношения тесно переплетены с материальными и лишь корректируют распределение ресурсов между особями или передачу вещества и энергии с одного трофического уровня на другой.
Разделение взаимоотношений в естественных экосистемах по полезности и вредности некорректно: любые взаимоотношения помогают поддерживать экологическое равновесие и в конечном итоге являются полезными для всех видов, которые входят в состав экосистемы. О полезности и вредности отношений между организмами можно говорить только в том случае, если в эти отношения вмешивается человек.
Понятие «экологическое равновесие» подвергается жесткой критике некоторых экологов (например, Гиляров, 2002) которые считают, что этот феномен не существует в природе, а само понятие уходит корнями в представления Карла Линнея о божественной гармонии природы (см. разд. 1.1). Оппоненты понятия «экологическое равновесие» считают, что любые взаимоотношения организмов и любая экосистема как сумма этих взаимоотношений всегда динамичны, так как испытывают влияние множества факторов.
Однако мы полагаем, что понятие экологического равновесия, если оно не абсолютизируется, а рассматривается как некоторая условность (подобная идеальному газу или ускорению под действием силы земного притяжения), полезно. Системы «хищник — жертва», «паразит — хозяин» и более сложные элементы экосистемы, такие как пищевые цепи, в состоянии экологического равновесия характеризуются соотношением численности и биомассы взаимодействующих организмов, которые колеблются вокруг некоторой средней величины, соответствующей экологическому равновесию. Это понятие необходимо при разработке системы рационального использования и охраны природы. Возможно, что более точно этот феномен отражают термины «динамическое равновесие» или «подвижное равновесие».
Конкуренция — это соревнование организмов одного трофического уровня (между растениями, между фитофагами, между хищниками и т.д.) за потребление ресурса, имеющегося в ограниченном количестве. Д. Тилман (TUman, 1982, 1983) подчеркивает, что особую роль играет конкуренция за потребление ресурсов в критические периоды их дефицита (например, между растениями за воду в период засухи или хищниками за жертвы в неблагоприятный год). Конкуренция играет большую роль в определении видового состава экосистем.
Принципиальных различий у межвидовой и внутривидовой (внутрипопуляционной) конкуренции нет. Возможны случаи, когда внутривидовая конкуренция является более острой, чем межвидовая, и наоборот. При этом интенсивность конкуренции внутри популяции и между популяциями может меняться в различных условиях. Если условия неблагоприятны для одного из видов, то конкуренция между его особями может усиливаться. В этом случае он может быть вытеснен (или чаще - потеснен) видом, для которого эти условия оказались более подходящими.
Однако в многовидовых сообществах пар «дуэлянтов» чаще всего не образуется и конкуренция носит характер диффузной: много видов одновременно конкурируют за один или несколько факторов среды. «Дуэлянтами» могут быть лишь массовые виды растений, которые делят один и тот же ресурс (например, деревья - липа и дуб, сосна и ель и т. д.).
У растений возможна конкуренция за свет, ресурсы почвы и за опылителей. На почвах, богатых ресурсами минерального питания и влагой, формируются густые сомкнутые растительные сообщества, где лимитирующим фактором, за который конкурируют растения, является свет. При дефиците в почве влаги или элементов минерального питания полог растений бывает разомкнутым и они не конкурируют за свет, а соревнуются за потребление почвенных ресурсов.
При конкуренции за опылителей побеждает тот вид, который более привлекателен для насекомого. Так, одним из факторов быстрого распространения в Европе гималайского заносного вида недотроги; железконосной УтраМепя glandulifera) является то, что он продуцирует больше нектара, чем его конкуренты в тех же влажных местообитаниях — чистец болотный, дербенник иволистный. Кроме того, нектар недотроги слаще (Chittka, Schurkens, 2001).
У животных конкуренция происходит за ресурсы пищи. Например, травоядные конкурируют за фитомассу. При этом конкурентами крупных копытных могут быть насекомые, подобные саранче, или мышевидные грызуны, способные в годы массового размножения уничтожить большую часть травостоя. Хищники конкурируют за жертвы.
Поскольку количество пищи зависит не только от экологических условий, но и от площади, где воспроизводится ресурс, конкуренция за пищу может перерастать в конкуренцию за занимаемое пространство, т. е. быть не только эксплуатационной, но и интерференционной (см. разд. 8.2). Снижение конкуренции в этом случае возможно при разделении территории на «охотничьи наделы» или на «загоны для выпаса». К примеру, косяки - семейные группы башкирской лошади, в поведении которых сохранились черты их диких предков, рассредоточиваются по «загонам для выпаса», что снижает конкуренцию за фитомассу и способствует равномерному использованию травостоев. По этой причине лошади - это идеальные фитофаги для охраняемых территорий в степной зоне.
Как и в отношениях между особями одной популяции, конкуренция между видами (их популяциями) может быть симметричной или асимметричной. При этом ситуация, когда условия среды одинаково благоприятны для конкурирующих видов, встречается довольно редко, и потому отношения асимметричной конкуренции возникают чаще, чем симметричной.
При флюктуирующих ресурсах, что обычно наблюдается в природе (увлажнение или элементы минерального питания для растений, первичная биологическая продукция для разных видов фитофагов, плотность популяций жертв для хищников), преимущества поочередно получают разные конкурирующие виды. Это также ведет не к конкурентному исключению более слабого, а к сосуществованию видов, которые попадают в более выгодную и менее выгодную ситуацию. При этом ухудшение условий среды виды могут переживать при снижении уровня метаболизма или даже перехода в состояние покоя.
Кроме того, на исход конкуренции часто влияет то, какой из видов первым начал заселять экотоп. Это особенно характерно для маловидовых сообществ водных растений, где более слабый вид может удерживать занятое место, в силу того, что и он его занял первым. Впрочем, лотерея способна влиять и на состав сообществ с большим числом претендентов на свободное место. В тропическом лесу на одном гектаре может быть до 150 видов деревьев, и потому занять место выпавшей особи могут представители разных видов.
Влияет на исход конкуренции и то, что победить в конкурентной борьбе больше шансов имеет популяция, в составе которой больше особей и которая, соответственно, будет более активно воспроизводить «свою армию» (так называемый массэффект).
Наконец, конкуренция между видами протекает на фоне отношений с организмами других трофических уровней (хищниками и паразитами). Это также влияет на исход конкуренции, поскольку более привлекательный как пищевой ресурс вид имеет меньше шансов победить в конкуренции. В итоге в естественных экосистемах виды сосуществуют даже при наличии асимметричной конкуренции, которая должна была бы привести к вытеснению одного из видов. Конкурентное исключение чаще всего наблюдается только в искусственных условиях «микрокосма», когда два конкурирующих вида изолированы и помещены в условия стабильной среды (например, в смешанном посеве двух культурных растений с разными конкурентными возможностями).
В естественных экосистемах существуют и специальные механизмы, которые снижают конкуренцию. Главный механизм — дифференциация экологических ниш (см. гл. 9), при которой различные организмы используют разные ресурсы.
Взаимоотношения «фитофаг — растение»
Взаимоотношения «фитофаг — растение» является первым звеном пищевой цепи, в котором вещество и энергия, накопленные продуцентами, передаются консументам. Для растений в равной мере «невыгодно», чтобы их съели до конца или не съели вовсе. По этой причине в естественных экосистемах проявляется тенденция формирования экологического равновесия между растениями и поедающими их фитофагами. Для этого растения: защищаются от фитофагов колючками, образуют розеточные формы с прижатыми к земле листьями, малодоступными для пасущихся животных;
защищаются от полного выедания биохимическим путем, продуцируя при усилении поедания токсичные вещества, которые делают их менее привлекательными для фитофагов (это особенно характерно для медленно растущих патиентов). У многих видов при их поедании образование «невкусных» веществ усиливается;
выделяют запахи, отпугивающие фитофагов.
Защита от фитофагов требует значительных затрат энергии, и потому во взаимоотношениях «фитофаг - растение» прослеживается трейдофф: чем растение быстрее растет (и соответственно, чем благоприятнее условия для его роста), тем оно лучше поедается, и, наоборот, чем растение медленнее растет, тем оно менее привлекательно для фитофагов. Интенсивное отрастание позволяет растениям с высокой поедаемостью сохраняться и даже доминировать в сообществах.
В то же время перечисленные средства защиты не обеспечивают полную сохранность растений от фитофагов, так как это повлекло бы за собой ряд нежелательных последствий для самих растений:
несъеденная степная трава превращается в ветошь -войлок, который ухудшает условия жизни растений. Появление обильного войлока ведет к накоплению снега, задержке начала развития растений весной и как итог — к разрушению степной экосистемы. Вместо степных растений (ковылей, типчака) обильно развиваются луговые травы и кустарники. У северной границы степи после этой луговой стадии вообще может восстановиться лес;
«лишние» листья многих видов трав и кустарников делают крону чрезмерно густой, что ухудшает условия для фотосинтеза (затененные листья «паразитируют», т. е. тратят на дыхание больше органического вещества, чем производят его в процессе фотосинтеза). Этот феномен, который исследовал А.А. Любищев, объясняет благоприятное влияние на урожайность посевов присутствия некоторого количества «вредителей»-фитофагов, которые осветляют полог растений. Поедание некоторого количества листьев побегов как бы запрограммировано у культурных растений в «память» о своем диком прошлом;
— в саванне уменьшение потребления побегов деревьев веткоядными животными (антилопами, жирафами и др.) приводит к тому, что их кроны смыкаются. В итоге учащаются пожары и деревья не успевают восстанавливаться, саванна перерождается в заросли кустарников.
Кроме того, при недостаточном потреблении растений фитофагами не освобождается место для поселения новых поколений растений.
Подобным образом регулируется равновесие между популяциями видов фитопланктона и зоопланктона. Активно поедаемые водоросли быстро размножаются. Некоторые водоросли, напротив, защищаются от выедания специальными выростами на твердых панцирях (как диатомовые) или объединением в большие колонии, которые не могут быть отфильтрованы рачками. Колониальными формами представлено большинство видов цианобактерий. Защите водорослей от выедания помогает их способность образовывать покоящиеся стадии, играющие ту же роль, что и банки семян у растений. Наконец, некоторые водоросли заглатываются планктонными животными-фитофагами, но не перевариваются и выделяются с экскрементами живыми.
«Несовершенство» отношений «фитофаг — растение» приводит к тому, что достаточно часто случаются кратковременные вспышки плотности популяций фитофагов и временное угнетение популяций растений, вслед за которыми следует и снижение плотности популяций фитофагов. Например, в степях Монголии один раз в 5—7 лет наблюдаются вспышки численности популяций полевки Брандта, которая полностью выедает надземные части степных растений и разрушает дернину. В итоге угнетаются популяции ковылей, а корневищные злаки быстро разрастаются за счет банка вегетативных зачатков (почек на корневищах). Вслед за вспышкой численности наступает депрессия популяций грызунов из-за массового заболевания, а популяции ковылей восстанавливаются.
Для обеспечения экологического равновесия в паре «фитофаг — растение» адаптации растений и животных, как правило, недостаточно. Оно возможно только в случае, если имеется зоофаг, который контролирует плотность фитофага. По этой причине невозможно экологическое равновесие, если из экосистемы исчез хищник или в нее внедрился вид-фитофаг, для которого нет контролирующего его хищника.
При разведении сельскохозяйственных животных плотность популяций фитофагов регулируется человеком, который сам устанавливает норму потребления фитомассы животными. В большинстве случаев при этом норма оказывается завышенной, что ведет к развитию процессов пастбищной дегрессии в экосистемах.
Взаимоотношения «хищник — жертва»
Взаимоотношения «хищник - жертва» представляют звенья процесса передачи вещества и энергии от фитофагов к зоофагам или от хищников низшего порядка к хищникам высшего порядка.
Как и при отношениях «фитофаг - растение», ситуация, при которой все жертвы будут съедены хищниками, что в конечном итоге приведет и к их гибели, в природе не наблюдается. Экологическое равновесие между хищниками и жертвами поддерживается специальными механизмами, исключающими полное истребление жертв. Так, жертвы могут:
- убегать от хищника. В этом случае в результате адаптации повышается подвижность и жертв, и хищников, что особенно характерно для степных животных, которым негде прятаться от преследователей («принцип Тома и Джерри»);
— приобретать защитную окраску («притворяться» листьями или сучками) или, напротив, яркий цвет, например красный, предупреждающий хищника о горьком вкусе. Общеизвестно изменение окраски зайца в разные времена года, что позволяет ему маскироваться летом в листве, а зимой — на фоне белого снега;
распространяться группами, что делает их поиск и промысел для хищника более энергоемким;
прятаться в укрытия;
переходить к мерам активной обороны (травоядные, имеющие рога, колючие рыбы), иногда совместной (овцебыки могут занимать «круговую оборону» от волков и т.д.).
В свою очередь хищники развивают не только способность к быстрому преследованию жертв, но и обоняние, позволяющее по запаху определить место нахождения жертвы. Многие виды хищников разрывают норы своих жертв (лисы, волки). В то же время сами они делают все возможное для того, чтобы не обнаружить своего присутствия. Этим объясняется чистоплотность мелких кошачьих, которые для устранения запаха много времени тратят на туалет и закапывают экскременты. Хищники одевают «маскировочные халаты» (полосатость щук и окуней, делающих их менее заметными в зарослях макрофитов, полосатость тигров и т.д.).
Полной защиты от хищников всех особей в популяциях животных-жертв тоже не происходит, так как это привело бы не только к гибели голодающих хищников, но в конечном итоге — к катастрофе популяций жертв. В то же время при отсутствии или снижении плотности популяции хищников ухудшается генофонд популяции жертв (сохраняются больные и старые животные) и ввиду резкого увеличения численности животных подрывается кормовая база.
Тем не менее даже при столь богатом арсенале средств защиты у жертв и нападения у хищников экологическое равновесие в паре «хищник — жертва» только за счет их биотических потенциалов невозможно. И в этом случае, как и в отношениях фитофагов и растений, необходимо третье звено пищевой цепи - естественный «враг» хищника - хищник более высокого порядка или паразит. В зависимости от характера жертвы и типа хищника (истинный, убивающий жертву сразу, или пастбищник, использующий ее многократно, как, например, оводы или слепни) возможно разное соотношение динамик их популяций. При этом картина осложняется тем, что хищники редко бывают монофагами (т. е. питающимися одним видом жертвы по типу «преследования»). Чаще всего, когда истощается популяция одного вида жертвы и ее добыча требует слишком больших затрат сил, хищники переключаются на другие виды жертв. Кроме того, одну популяцию жертв может эксплуатировать несколько видов хищников и паразитов.
По этой причине часто описываемый в экологической литературе эффект зависимости численности популяций жертв и хищников - пульсация численности популяции жертвы, за которой с некоторым запаздыванием пульсирует численность популяции хищника («эффект Лотки — Вольтерры»), — выражен далеко не во всех парах «хищник - жертва». П.В. Турчин (2002) считает осцилляторный (колебательный) характер динамики системы «ресурс - потребитель», которая включает отношения типа «фитофаг - растение» и «хищник — жертва», третьим законом популяционной экологии.
При интенсивной эксплуатации популяций фитофагов человек нередко исключает из экосистем хищников (в Великобритании, к примеру, есть косули и олени, но нет волков; в искусственных водоемах, где разводят карпа и другую прудовую рыбу, нет щук). В этом случае роль хищника выполняет сам человек, изымая часть особей популяции фитофага. При этом он должен руководствоваться экологическими нормативами «максимально допустимого урожая, (МДУ)», отражающими скорость восстановления плотности популяций. К примеру, без риска разрушить популяцию северного оленя из нее можно ежегодно изымать до 40% животных, а из популяции лосей - только 20%. Примерно также показатель МДУ используется при эксплуатации популяций растений.
Взаимоотношения «паразит — хозяин». Паразиты — это организмы, которые питаются за счет организма хозяина. Это очень разнообразная группа организмов (животные, растения, грибы, бактерии), которую изучает специальная наука — паразитология.
Паразиты не убивают хозяина, а длительное время используют его как пищевой ресурс и убежище, сокращая при этом продолжительность жизни и плодовитость «организма-дома». Паразиты близки к хищникам-пастбищникам, но в отличие от последних, которые используют несколько жертв, паразит связан с одним организмом-хозяином (а если с несколькими, то меняет их в ходе жизненного цикла). Различаются следующие группы паразитов.
Биотрофы и некротрофы. Биотрофы всю жизнь питаются за счет живого хозяина, а некротрофы губят его (или часть его тела, например лист растения) и завершают свою биографию как рассматриваемые далее детритофаги.
Микропаразиты и макропаразиты. Различаются по размерам. К микропаразитам относятся вирусы, бактерии, микроскопические грибы и простейшие, к макропаразитам — все прочие.
Истинные паразиты и паразитоиды. Истинные паразиты всю свою жизнь питаются за счет организма-хозяина (или нескольких хозяев, если в течение жизненного цикла переселяются из организма одного вида в другой). Паразитоиды (как правило, насекомые) на определенных стадиях жизненного цикла ведут свободный образ жизни (питаются, как фитофаги или зоофаги). После этого они откладывают яйца в тело организма-хозяина, в котором паразитируют личинки. Паразитоиды представляют переход к хищникам. Насекомые-паразитоиды используются для биологического метода контроля насекомых-вредителей в сельском хозяйстве (трихограмма, теленомус-наездник и др.). Эндотрофные и эктотрофные паразиты. Эндотрофные паразиты живут внутри организма-хозяина (глисты в кишечнике позвоночных, стеблевые нематоды в стеблях хлебных злаков), а эктотрофные - на поверхности организма-хозяина (блохи, вши, клещи и др.). Особый случай эктотрофного паразитизма — образ жизни карликовых самцов глубоководных удильщиков, которые внедряются острыми зубами в тело самки, после чего тела хозяина и паразита сливаются воедино (объединяются даже кровеносные системы). Тело самки в 13 раз больше тела самца. Экологический смысл этой адаптации заключается в повышении вероятности оплодотворения. Удильщики живут в полной темноте, поэтому иные варианты поиска «спутника жизни» реализовать крайне сложно.
Суперпаразиты — «паразиты паразитов». Существуют суперпаразиты от первого до четвертого порядка (их можно представить в виде матрешки), верхний уровень представлен бактериями или вирусами. В этом случае из паразитов формируется пищевая цепь.
Различаются группы паразитов и по сложности жизненного цикла. Одни виды паразитов передаются при непосредственном контакте особей хозяина (например, вирусы и бактерии, вызывающие болезни человека). Другие паразиты перед заражением основного хозяина, в теле которого они образуют потомство, проходят через один или несколько видов промежуточных хозяев (например, широкий лентец - паразит человека, но в течение жизненного цикла он проходит через стадии жизни в рачках-циклопах и рыбах).
Наконец, кроме паразитов существуют еще и полупаразиты. Значительное число видов растений (в средней полосе в первую очередь из семейства норичниковых) сочетает автотрофное питание и паразитирование на корнях растений-хозяев. При этом если каждый вид растений-паразитов (из семейств заразиховых, повиликовых и др.) имеет своего «персонального» хозяина, то число видов-хозяев для одного вида полупаразита исчисляется десятками и сотнями. В этом случае нет необходимости в тонком подстраивании метаболизма хозяина и полупаразита, как при абсолютном паразитизме, полупаразит получает от растения-хозяина только неспецифические элементы питания.
Среди полупаразитов есть виды, наносящие вред хозяйству человека. Например, погремки (Rhinanthus) при доминировании в луговых сообществах становятся основными продуцентами органического вещества. В этом случае за счет потери энергии при переходе с одного трофического уровня на другой биологическая продукция травостоя снижается в 2-3 раза. Причем погремок не поедается сельскохозяйственными животными ни на пастбище, ни в сене.
В естественных экосистемах взаимоотношения «паразит — хозяин» являются одним из важных факторов поддержания экологического равновесия. Особенно велика их роль при контроле плотности популяций крупных животных, у которых нет естественных врагов-хищников (слон, бегемот, крокодил, лев и др.). При отсутствии паразитов их отношения с жертвами могли бы быть нарушены.
В процессе длительной коэволюции (взаимо приспособления) паразитов и хозяев вырабатываются специальные механизмы, которые позволяют им устойчиво сосуществовать.
Защитные реакции хозяев могут быть следующими:
иммунный ответ организма, т. е. возникновение биохимических реакций, которые сдерживают массовое развитие паразитов;
сбрасывание зараженных частей (это особенно характерно для растений-хозяев, которые сбрасывают сильно зараженные листья). В этом случае паразиты продолжают жить уже как детритофаги;
выработка устойчивости к влиянию паразитов за счет быстрого роста здоровых тканей взамен пораженных (это имеет место при паразитировании тли);
изоляция органов поражения как «зеленых островов» (формирование галлов у дуба, орешника и других растений после того, как насекомое-паразитоид отложит в ткани листа яйцо);
уменьшение плотности популяций хозяев, что снижает вероятность распространения паразита и заражения им. Зараженные животные менее подвижны и становятся более легкой
добычей хищников, которые таким образом снижают долю зараженных особей в популяции;
- формирование гетерогенных популяций хозяев, в составе которых есть экотипы, устойчивые к паразитам. Эти экотипы являются основой адаптивной селекции на повышение устойчивости культурных растений к грибковым заболеваниям.
В естественных экосистемах формирование экологического равновесия между популяциями паразитов и их хозяев — нормальное явление. При этом в отличие от отношений «фитофаг — растение» или «хищник - жертва» оно возможно без третьего участника. В силу того, что паразиты связаны с ограниченным кругом хозяев, эта связь математически описывается проще, чем связь между хищниками и их жертвами. Во многих случаях проявляется модель Лотки-Вольтерры: плотность популяций обоих видов изменяется циклически, но пики плотности паразитов запаздывают по отношению к пикам плотности хозяев.
Ситуация изменяется в антропогенных экосистемах, особенно в сельскохозяйственных, где заражение скота паразитами может привести к гибели многих животных. Представляют опасность взаимоотношения паразитов и человека, который может заболевать гельминтозами, вызываемыми разными видами глистов, лямблиозом (при заражении простейшим — лямблией), болезнями бактериальной и вирусной природы.
Катастрофическими бывают последствия заноса паразитов в новые районы, где у их потенциальных хозяев отсутствуют механизмы защиты от паразитов. В XX в. произошли ботанические катастрофы в Америке (гибель зубчатого каштана Castanea dentata от занесенного туда из Китая паразитического гриба Endothia parasitica, вызывающего «рак каштана») и Европе, где от «голландской болезни» почти полностью исчез вяз. Болезнь вызывает гриб Ophiostoma ulmi, который переносится жуком-короедом.
К еще более серьезным последствиям привело распространение вируса «коровьей чумы» в саваннах Африки (McNaughton, 1992). Из-за этого вируса, который первоначально вызвал болезни у домашнего скота, а затем поразил многие виды диких крупных копытных животных, резко снизилась интенсивность поедания фитофагами растительности саванны, и в первую очередь ее древесно-кустарникового компонента, что сказалось на его отношениях с травяным покровом саванны. Кустарники и деревья стали бурно разрастаться, подавляя травы, что увеличило количество пожаров, которые при таком сомкнутом древесном пологе стали более частыми. После пожаров деревья отрастали плохо и замещались кустарниками, корневища которых позволяли им сохраняться во время пожара. Только в 70-е годы, когда удалось снять влияние на экосистемы вируса «коровьей чумы», вылечив от болезни домашний скот, поставлявший паразита популяциям диких животных через выделение слюны на пастбищах, где их выпас чередовался, процесс был остановлен. Восстановилась плотность популяций животных, питающихся ветками, и, соответственно, восстановился баланс между древесно-кустарниковым и травяным компонентами экосистемы саванны, которая приобрела первозданный облик. Есть примеры более сложных взаимоотношений «паразит - хозяин» с посредником. Так, гетеротрофное растение-паразит подъельник паразитирует на грибах, разлагающих мертвое органическое вещество, но, кроме того, по гифам микоризного гриба, как по шлангу, выкачивает питательные элементы из корней ели.
Мутуализм — это форма взаимоотношений организмов, при которых партнеры получают пользу. Отношениями мутуализма связаны организмы, не конкурирующие за ресурсы. Мутуализм включает разнообразные формы сотрудничества - от облигатного (симметричного или асимметричного), при нарушении которого гибнут оба или
один из сотрудничающих партнеров, до факультативного, которое помогает выживать партнерам, но не является для них обязательным (так называемая протокооперация). Рассмотрим основные варианты мутуализма.
Растения и микоризные грибы. Такие взаимоотношения с грибами (микотрофия) свойственны большинству видов наземных сосудистых растений (цветковых, голосеменных, папоротников, хвощей, плаунов), что во многом облегчило заселение растениями суши (Заварзин, 2000). Микоризные грибы могут оплетать корень растения и проникать в ткани корня, не нанося ему при этом существенного ущерба (эндотрофные и эктотрофные микоризы).
Грибы, не способные к фотосинтезу, получают из корней растений органические вещества, а у растений за счет разветвленных грибных нитей в сотни и тысячи раз увеличивается всасывающая поверхность корней. Кроме того, некоторые микоризные грибы не просто пассивно всасывают элементы питания из почвенного раствора, но одновременно выступают в роли редуцентов и разрушают сложные вещества до более простых. При этом, микоризные грибы, выделяя антибиотики, защищают корни растений от патогенов.
Микоризные грибы - «дорогое удовольствие» для растений, так как использование их в качестве посредников для обеспечения элементами питания и водой сопряжено со значительными затратами продуктов фотосинтеза (1/3 или даже 1/2 валовой первичной продукции). По этой причине при улучшении условий минерального питания, например удобрении лугов, даже типичные микотрофные растения отказываются от микориз и переходят на «самообслуживание». Не тратятся на содержание микориз виды-нитрофилы (распространенные на почвах с высоким содержанием нитратного азота) из семейств маревых, крестоцветных и некоторых других, поселяющиеся на нарушенных местообитаниях, где за счет минерализации органического вещества в почве резко возрастает количество нитратов. При этом микоризы, которыми обладают виды следующих стадий сукцессии восстановления экосистем после нарушений (см. разд. 12.6), выделяют вещества, подавляющие «самостоятельные» растения. Это ускоряет процесс вытеснения нитрофилов.
Микоризы нет у водных растений, сравнительно редко она встречается у растений, обитающих в экстремальных условиях - пустынях, горных и арктических тундрах. Как подчеркивает ТА. Работнов (1992), большинство микотрофов - это мезофиты умеренно богатых почв.
Микоризы у травянистых растений, как правило, не видоспецифичны (т. е. один вид грибов может формировать микоризу у разных видов растений), а у древесных - видоспецифичны. Таким образом, плодовые тела подберезовика, подосиновика, масленка или рыжика образуются за счет продукции фотосинтеза соответствующих видов деревьев.
Поскольку микоризные грибы оплетают корни нескольких рядом произрастающих растений, по ним возможен горизонтальный перенос элементов питания от одного растения к другому по «гифопроводам». A.M. Гиляров (2003) рассматривает это как «экзаптации на уровне сообществ», т. е. как побочный эффект адаптации микоризного гриба к нескольким видам растений. Данных о количестве веществ, перекачиваемых по микоризам из одного растения в другое, мало. Можно полагать, что оно невелико, тем не менее смягчает отношения конкуренции и повышает общую устойчивость экосистем.
Растения и микроорганизмы-азотфиксаторы. Возможны две формы такого мутуализма — облигатный мутуализм и протокооперация. В первом случае азотфиксирующие микроорганизмы живут в корнях растений (бобовых, облепихи, ольхи и некоторых других), вызывая образование клубеньков. Процесс связывания атмосферного азота облигатными азотфиксатора-ми называется симбиотической азотфиксацией. Во втором случае - при протокооперации - азотфиксирующие микроорганизмы населяют примыкающую к корням часть почвы (ризосферу) и усваивают органические вещества, которые, как в проточном культиваторе, постоянно выделяются корнями. Такая азотфиксация называется ассоциативной. В целом ассоциативная азотфиксация преобладает в естественных экосистемах, симбиотическая — в агроэкосистемах.
Симбиотические микроорганизмы могут жить и в листьях, пример — водный папоротник азолла, распространенный в тропическом поясе. Связанная с азоллой цианобактерия анабена способна за год фиксировать до 1000 кг/га азота (что является бесспорным рекордом, достойным книги Гиннеса). Для сравнения: посев клевера в средней полосе способен за год фиксировать до 200 кг/га азота, а люцерны в жарких районах с удлиненным полевым периодом и при поливе — до 700 кг/га. Оптимальная доза внесения азотных удобрений в разных условиях в зависимости от культуры колеблется от 50 до 200 кг/га; в настоящее время вносится в среднем 10 кг/га азотных удобрений в действующем веществе.
Обеспечение новых («мертвых») субстратов азотом является необходимым условием для их зарастания. В теплом климате азот в субстрате накапливается в результате симбиотической азотфиксации: пионерами заселения лавовых потоков, отложений речного аллювия, горных осыпей являются бобовые растения (особенно часто из рода люпин). В более прохладном климате азот поставляется в результате ассоциативной азотфиксации: новые субстраты зарастают злаками и осоками. В самых суровых условиях Севера пионерами оказываются цианобактерии, которые обладают уникальной способностью и к фотосинтезу, и к азотфиксации.
Мутуалистические взаимоотношения с азотфиксаторами, так же как и содержание микориз, обходятся растениям очень дорого: на них затрачивается значительное количество продуктов фотосинтеза (около 1/3). Большими затратами органического вещества на симбиотическую азотфиксацию объясняются более низкие урожаи зернобобовых культур по сравнению со злаками.
Тем не менее на биологическую азотфиксацию экологи возлагают большие надежды, она должна во многом заменить техногенную азотфиксацию промышленных предприятий, при которой на производство минеральных азотных удобрений затрачивается очень много энергии. Кроме того, экологически грязным является не только само производство удобрений, но и их использование: при внесении азотных удобрений на поля до 50% их вымывается в окружающую среду, вызывая ее загрязнение (в первую очередь эвтрофикацию водных экосистем, см. разд. 12.7).
Растения и насекомые-опылители. Насекомые, переносящие пыльцу, питаются нектаром или пыльцой. Отмечены случаи участия насекомых в опылении даже таких типично ветроопыляемых растений, как злаки. Насекомые-опылители переносят пыльцу с одного цветка на другой на большие расстояния, чем ветер. Если пыльца деревьев за время, пока рыльцевая поверхность сохраняет способность ее воспринимать, может быть перенесена ветром не более чем на 70 м (у трав - менее 10 м), то за это время шмели переносят пыльцу на расстояние до 3 км. Радиус переноса пыльцы пчелами обычно ограничен 1 км.
Существуют два основных направления развития мутуализма растений и насекомых: узкая и широкая специализация (т. е. в направлении облигатного мутуализма и протокоопера-ции). При узкой специализации эволюция ведет к ограничению числа опылителей: происходит усложнение строения цветка (как у бобовых или губоцветных) таким образом, что нектар становится доступным только для насекомых с определенным типом строения тела (в первую очередь ротового аппарата). Высшее достижение этого варианта эволюции - взаимоотношения опылителей и некоторых представителей орхидных, которые привлекают самцов насекомых-опылителей, имитируя облик и половые феромоны самок.
При широкой специализации спектр опылителей возрастав, такой спектр опылителей имеют представители семейства сложноцветных. Этим объясняется их высокая устойчивость в антропогенно нарушенных экосистемах с обедненным видовым составом опылителей. По этой причине в современном нарушаемом человеком мире обязательный мутуализм растений и насекомых менее выгоден для обоих партнеров, чем протокооперация.
Эффективность протокооперации возрастает благодаря неодновременному цветению разных видов растений, опыляемых одним видом насекомых. Более того, как правило, насекомые посещают цветки именно в апогей их цветения, когда продукция нектара максимальна. В тропиках опылителями некоторых растений являются птицы и летучие мыши.
Растения и животные, распространяющие их семена. Распространение плодов (и семян) растений с помощью животных (зоохория) широко представлено в природе. Агентами-распространителями являются птицы, поедающие сочные плоды, медведи, копытные, насекомые. При прохождении через пищеварительный тракт животных семена зоохорных растений не только не перевариваются, но даже повышают всхожесть.
Кроме плотных покровов, защищающих семена от переваривания, существуют другие приспособления для зоохории. Так, на семенах растений (многих губоцветных, лилейных, маковых, молочайных, лютиковых, сложноцветных), распространяемых муравьями, имеются специальные придатки, богатые маслом, которые привлекают муравьев и используются ими в пищу. Сухие зоохорные плоды снабжены различными крючочками и щетинками для прикрепления к шерстному покрову животных, например у репейничка, череды, чернокорня.
С помощью животных распространяются споры некоторых видов грибов и мхов.
Водоросли и грибы в лишайнике. В этом случае мутуализм столь обязателен и функции сотрудничающих организмов столь прилажены друг к другу, что по существу возникает новый «организм второго порядка». Водоросль обеспечивает гриб органическими веществами, гриб водоросль - водой и минеральными элементами.
Этот вариант обязательного мутуализма представлен весьма широко: имеются сотни видов лишайников. Лишайники первыми заселяют поверхность скал и широко распространены на Севере — в условиях крайней ограниченности ресурсов тепла и элементов минерального питания.
Долгие годы в литературе ведется дискуссия о симметричности отношений гриба и водоросли в лишайнике. В последнее время все чаще эти отношения рассматриваются как асимметричные: большую «выгоду» от симбиоза получает гриб («водоросль - это царевна, плененная жестоким драконом»).
Млекопитающие и микроорганизмы, населяющие их пищеварительный тракт. Большинство животных, включая человека, но особенно травоядные, сами не в состоянии переваривать пищу, так как не имеют ферментов, разрушающих целлюлозу, и эту роль играют микроорганизмы - бактерии и некоторые простейшие, которые живут в их желудочно-кишечном тракте. В кишечном тракте гладких китов среди 1000 видов бактерий были найдены даже те, которые могут разрушать органические вещества, присутствующие в нефтепродуктах. Возможно, что наличием таких симбионтов объясняется сравнительно высокая устойчивость этого вида китов к нефтяному загрязнению океана.
Хемоавтотрофные бактерии и низшие животные. Червеобразные животные-вестиментиферы (тип погонофоры) в стадии личинки являются типичными гетеротрофами со ртом, пищеварительным каналом и анусом. Однако после того как они заглатывают серобактерии, происходит редукция органов пищеварения, клетки животного заполняются серобактериями и вестиментиферы становятся «симбиотическими автотро-фами». В результате мутуализма бактерии получают защиту, а животное — органическое вещество.
Содержание сероводорода в среде, где обитают вестиментиферы, таково, что может погибнуть любой другой организм. Их спасает особый тип гемоглобина, который связывает не только кислород, но и серу. Мутуализм позволяет вестименти-ферам очень быстро расти и достигать длины 2,5 м.
В.В.Малахов (2001) считает, что мутуализм вестиментифер и бактерий возник в результате развития пищевых отношений типа «хищник — жертва». В гидротермальных оазисах океана и сейчас существуют свободноживущие серобактерии, которые формируют «маты», служащие пищей для многих животных. Начав с питания такими свободноживущими бактериями, вестиментиферы со временем вступили с ними в отношения мутуализма.
Подобным образом питаются и другие погонофоры, связанные отношениями мутуализма уже не с серобактериями, а с метанобактериями. Бактерии используют метан, образующийся в нефтяных пластах и поступающий в океан по трещинам в плитах литосферы. Это позволяет использовать погонофор в качестве биологических индикаторов месторождений нефти.
Кишечнополостные и водоросли. Водоросли поселяются в теле кораллов, заключенных в известковый скелет, и снабжают животное органическим веществом. Животное поставляет водорослям питательные элементы и дает убежище. В отличие от погонофор кораллы являются гетеротрофами, которые питаются зоопланктоном. Водоросли лишь помогают наиболее эффективно использовать вещества, полученные при гетеротрофном питании. Этим мутуализмом объясняется быстрый рост коралловых рифов.
Человек и сельскохозяйственные животные и культурные растения. Этот вариант мутуализма является протокооперацией, тем не менее ни человек при современной плотности населения на планете не может обойтись без сельскохозяйственных животных и растений, ни корова, пшеница или рис не могут выжить без человека. Причиной мутуализма является искусственный отбор, в результате которого из «эгоистических» побуждений человек усиливал у растений и животных эксплерентность и снижал патиентность и виолентность, что лишило эти организмы способности жить без его опеки. Особенно активно это направление селекции развивалось в 60—70-е годы. XX в., когда в странах субтропического и тропического поясов произошла Зеленая революция.
Широко распространен мутуализм водорослей и простейших в океанических экосистемах. Некоторые простейшие после поедания водорослей используют их хлоропласты, которые продолжают работать в теле простейшего до тех пор, пока не износятся, после чего перевариваются.
Как протокооперация (хотя и весьма слабая) могут рассматриваться взаимоотношения между бобовыми и злаками в сеяных травостоях: бобовые за счет связи с симбиотическими азотфиксирующими бактериями улучшают условия обеспечения злаков азотом, а вертикально ориентированные листья злаков, пронзающие густой травостой бобовых, понижают уровень конкуренции за свет.
Известно множество других «экзотических» вариантов мутуализма, например:
разведение грибов муравьями и жуками;
отношения африканской птицы медоуказчика и капского медоеда (птица находит пчелиное гнездо, а медоед вскрывает его);
отношения чистильщиков (птиц, рыб) и их «клиентов»:
отношения муравьев и акаций (муравьи охраняют акацию, соком которой питаются, от других фитофагов).
Комменсализм и аменсализм. Мутуализм связан плавным переходом с другим вариантом отношений организмов - комменсализмом, при котором сотрудничество выгодно только одному из партнеров. При этом на разных стадиях сотрудничества (или чаще в разных условиях) большую выгоду может получать то один, то другой партнер. Рассмотрим некоторые наиболее характерные случаи комменсализма.
«Растения-няни» и их подопечные. Береза и ольха могут быть «няней» для ели: «няня» защищает всходы ели от прямых солнечных лучей, так как на открытом месте елочки вырасти не смогут. В таких же отношениях состоят кустарники из семейств губоцветных и сложноцветных с южноамериканскими кактусами. Кактусы для снижения дневного испарения воды фотосинтезируют при закрытых устьицах (они используют углерод кислот, образованных из захваченного устьицами за ночь углекислого газа) и потому не «потеют». Взрослые кактусы имеют большую фитомассу, и им перегрев не страшен, а молодые могут развиваться только в тени засухоустойчивых кустарников, так как на открытом солнечном свете они гибнут. Весьма характерно, что в более благоприятных климатических условиях (например, при подъеме в горы, где климат более прохладный) кактусы не нуждаются в растениях-нянях.
Эпифиты и деревья. Как комменсализм можно рассматривать отношения эпифитов и деревьев-хозяев: они выгодны для эпифита, но безразличны для хозяев, поскольку эпифиты используют лишь отмершие ткани дерева или скопления пыли и опавших листьев.
Группа эпифитов достаточно разнообразна и включает лишайники, водоросли, папоротниковидные, цветковые растения. При этом сосудистые эпифиты распространены почти исключительно в теплом тропическом и субтропическом климате. В умеренном климате эпифиты представлены только лишайниками, которые селятся на стволах деревьев, и реже мхами. Отсутствие эпифитов в лесах умеренной полосы связано в первую очередь с их неспособностью переносить холодные периоды года.
Как правило, эпифиты — чистые комменсалы, но есть случаи, когда установлено их отрицательное влияние на дерево-хозяина. При этом отрицательное влияние оказывает не сам эпифит, а его микоризный гриб, который ведет себя как паразит. Если эпифитов очень много, то они создают дополнительную нагрузку на ветви, что может вызвать их поломку. Таким образом, «безвредность» эпифитов для деревьев относительна.
Детритофагия. На те популяции, организмы которых поедаются в мертвом состоянии, детритофаги практически не влияют. Однако это положение нельзя абсолютизировать: участвуя совместно с редуцентами в процессе минерализации органического вещества, детритофаги способствуют его возврату в окружающую среду и тем самым росту растений и всех организмов, которые связаны с ними в пищевых цепях.
В почве сконцентрировано огромное разнообразие детрито-фагов: число их видов на 1 м2 почвы в лесу умеренной полосы может превышать 1000, а общая плотность популяций простейших и нематод — 10 млн, ногохвосток (Collembola) и почвенных клещей (Acari) - 100 тыс. и т. д.
Почвенные детритофаги разделяются на группы микрофауны (в нее входят простейшие, нематоды и коловратки, а также специализированные животные, питающиеся микроорганизмами, т. е., строго говоря, к детритофагам не относящиеся), ме-зофауны (животные с шириной тела от 100 мкм до 2 мм), макрофауны (ширина тела от 2 до 20 мм) и мегафауны (ширина тела более 20 мм). В последнюю группу входят мокрицы (Isopoda), многоножки (Diplopoda), дождевые черви (Megadrili), улитки и слизни (Mollusca), личинки некоторых мух (Diptera) и жуков (Coleoptera). Ч. Дарвин считал, что только дождевые черви перерабатывают такое количество растительного детрита, что ежегодное поступление в почву их экскрементов достигает 50 т/га. В тропических районах это количество может быть в три раза выше.
Именно благодаря этой армаде детритофагов из мертвого органического вещества (в травяных экосистемах в первую очередь корней растений, в лесных - опада) формируется почва. При этом многие детритофаги одновременно являются и хищниками, так как питаются «бутербродами» из мертвого вещества и содержащихся в нем живых бактерий.
Особая группа детритофагов - копрофаги, питающиеся экскрементами. Большинство копрофагов питается экскрементами травоядных животных, которые усваивают поедаемую растительную массу не более чем на 10%, остальное выделяется ими в окружающую среду в непереваренном состоянии (существуют даже варианты детритофагии в сельском хозяйстве при повторном скармливании животным специально подготовленных экскрементов). Фауна копрофагов очень разнообразна, особенно в тропиках (в частности, в ее составе есть навозный жук (Heliocopris dilloni), имеющий размер до 6 см).
Копрофаги часто специализированы для «переработки» определенного типа экскрементов, что создало проблемы после появления в Австралии крупного рогатого скота, навоз которого местные копрофаги, привыкшие к «вкусу» помета кенгуру, перерабатывать отказались. Пришлось отправлять в Австралию жуков-копрофагов из Африки! Копрофагов, питающихся экскрементами хищников, много меньше. Это объясняется тем, что в данном случае корм переваривается более полно, и потому потенциальная пища детритофагов менее питательна.
Крупные детритофаги, питающиеся трупами животных, называются мусорщиками. Это - ворон, коршун, гриф, шакал, росомаха, барсук, песец и др. К этой же группе относятся некоторые крупные беспозвоночные, например жук-мертвоед некрофорус, который способен (вдвоем с самкой) закопать труп мыши на глубину до 20 см и там «скормить» его своим личинкам.
Среди детритофагов водных экосистем по способу добывания и переработки пищи различают размельчителей, собирателей, соскребателей, фильтраторов.
В заключение отметим, что взаимоотношения организмов могут меняться не только в зависимости от условий среды (обострение конкуренции при уменьшении ресурсов), но и на разных стадиях жизненного цикла. Пример тому - взаимоотношения семги и двустворчатого моллюска жемчужницы. Личинки жемчужницы паразитируют в жабрах семги, а взрослые моллюски живут независимо на дне и, фильтруя воду, улучшают условия для жизни рыбы, очень чувствительной к загрязнению. Кроме того, между скоплениями раковин прячется от хищников молодь семги. Уже описанный положительный «эффект группы» и отношения растений-нянь и их подопечных со временем также сменяется острой конкуренцией: в группе начинается процесс самоизреживания, а подопечные подавляют своих «нянь».
Положительные взаимодействия играют важную роль в организации экосистем, так как уравновешивают антагонизм конкуренции, хищничества и паразитизма.
Тема Демэкология или экология популяций
Лекция 6 Понятие о популяции
Первой надорганизменной биологической системой является популяция — группа совместно обитающих особей, элементарная единица эволюционного процесса и форма существования вида. Главным объектом исследований демэкологии является изучение взаимодействий на уровне отношений популяция - среда. Ее задача – изучение морфологических особенностей популяции, их возрастного состава численности, плотности рождения, смертность, расширение и характер расселения организмов, а также изучение внутривидовых и межвидовых отношений популяции.
Биологический вид — это совокупность особей, обладающих наследственным сходством морфологических, физиологических и биохимических особенностей, способных к скрещиванию с образованием плодовитого потомства, приспособленных к определенным условиям жизни и занимающих в природе определенную область (ареал). Виды часто занимают большой ареал, в пределах которого особи распределены неравномерно, группами — популяциями. Целостность вида поддерживается связями между популяциями.
Популяция — совокупность особей одного вида, способных к самовоспроизводству, которая длительно существует в определенной части ареала относительно обособленно от других совокупностей того же вида. Контакты между особями одной популяции чаще, чем между особями разных популяций. Например, уровень панмиксии (свободного скрещивания) внутри популяции выше, чем между особями разных популяций. Популяция является структурной единицей вида и единицей эволюции.
Популяция — это элементарная группировка организмов определенного вида, обладающая всеми необходимыми условиями для поддержания своей численности необозримо длительное время в постоянно изменяющихся условиях среды.
Популяция – совокупность особей одного вида, которые воспроизводят себя на протяжении большого количества поколений и долгое время занимают определенную территорию, функционируя и развиваясь в одном или нескольких биоценозах.
Популяция – группа особей одного вида способных свободно скрещиваться и достаточно долго поддерживать свое существование в данном месте обитания.
Популяция – элементарная эволюционная единица экологическим принципом, которой является плотность, разделенная по возрасту, полу и характер размещения в границах экосистемы или сообщества.
Ареал Пространство, на котором популяция или вид в целом встречается в течение всей своей жизнедеятельности, называется ареалом — областью распространения. Ареал может быть сплошным или разорванным (дизъюнктивным), если между его частями возникают различные преграды (водные, орографические и др.), пространства, не заселенные представителями данного вида. Выделяют различные центры ареалов: геометрический центр; центр возникновения вида в пределах ареала; центр обилия — часть ареала, на которой сосредоточено наибольшее количество особей.
В зависимости от величины ареала и характера распространения различают космополитов, убиквистов, эндемиков. Космополиты — виды растений и животных, представители которых встречаются на большей части обитаемых областей Земли (например, комнатная муха, серая крыса). Убиквисты — виды растений и животных с широкой экологической валентностью, способны существовать в разнообразных условиях среды, имеют обширные ареалы (например, тростник обыкновенный, волк). Эндемики — виды растений и животных, которые имеют небольшие ограниченные ареалы (часто встречаются на островах океанического происхождения, в горных районах и изолированных водоемах).
Для животных также различают трофический и репродуктивный ареалы, между которыми существует связь в виде путей пролета для птиц или путей миграции для некоторых млекопитающих и рыб.
Классификация популяций. Популяции различаются по размерам и степени генетической самостоятельности, длительности существования, способу размножения особей и т.д.
По размерам занимаемой популяцией территории и степени связи между особями различают элементарные (локальные), экологические и географические популяции.
элементарная (локальная) – совокупность особей вида, населяющих небольшой участок однородной площади. Количество этих участков может быть различным, зависит от конкретного биогеоценоза.
Экологическая – совокупность элементарных популяций, т.е. видовые сообщества, прикрепленные к конкретным биогеоценозам.
географическая - совокупность групп особей, предшествующего ранга, которые заселяют значительную территорию с географическими однородными условиями среды.
По способности к самовоспроизведению и самостоятельной эволюции популяции бывают перманентные (постоянные) и темпоральные (временные). Перманентные (постоянные) — популяции, относительно устойчивые в пространстве и во времени, способные к неограниченно длительному самовоспроизведению, являются элементарными единицами эволюции. Темпоральные (временные) — популяции, неустойчивые в пространстве и во времени, неспособные к длительному самовоспроизведению, с течением времени либо преобразуются в перманентные, либо исчезают.
По способу размножения популяции делят на панмиктические, клональные и клонально-панмиктические. Панмиктические популяции состоят из особей, размножающихся половым путем, для которых характерно перекрестное оплодотворение. Клональные популяции состоят из особей, для которых характерно только бесполое размножение. Клонально-панмиктические популяции образованы особями с чередованием полового и бесполого размножения.
Каждая популяция выполняет определенную функцию в экосистеме, одновременно взаимодействуя с популяциями того же вида, которые находятся за границами данного биоценоза. На основании этого выделяют следующие популяции:
1. Независимая - Распоряжается достаточным потенциалом рождаемости, который дает возможность пополнять потери ее численности и долго существовать без миграций особей извне.
2. Полузависимая – может существовать размножению собственных особей в условиях низкой численности. Иммиграция извне чувствительно влияет на последствия заселения
3. Зависимая – эта популяция, в которой рождаемость не покрывает потерь численности, такая популяция не существует без миграции извне
4. Псевдопопуляция – группа особей какой-либо популяции, которая не имеет возможности размножаться в данном месте и которая образуется благодаря миграции особей соседних сообществ, такие популяции могут существовать долго, участвовать в биоценотических процессах, но без возможного расширения.
5. Периодическая – популяция, появляющаяся в незаселенных биотоках или между границами данного вида в течение нескольких месяцев или лет. Ее существование связанно с периодическим возникновением благоприятных условий среды.
6. Гемиопопуляция – это популяция, в которой, отчетливо проявляются отличия жизненных потребностей в разных фазах жизненного цикла, отдельные индивиды занимают разное место в природе (комары).
Популяции, будучи групповыми объединениями, обладают рядом специфических свойств, которые не присущи каждой отдельной особи: численность, плотность, рождаемость, смертность, скорость роста и др. Кроме того, популяции свойственна определенная организация: половая, возрастная, генетическая, пространственно-этологическая и другие структуры.
Статические показатели популяции: численность, плотность, показатели структуры
Количественные показатели (характеристики) популяции можно разделить на статические и динамические. Статические показатели характеризуют состояние популяции на данный момент времени. Основные из них: численность, плотность, а также показатели структуры.
Численность популяции — общее количество особей на данной территории или в данном объеме, относящихся к одной популяции. Различают непериодические (редко наблюдаемые) и периодические (постоянные) колебания численности популяций.
Плотность популяции — среднее число особей на единицу площади или объема. Она измеряется числом особей или биомассой популяции на единицу производительности или объема, например 400 деревьев на 1 га , 2 млн. водорослей на 1 м3 воды, 100 кг рыбы на 1 га поверхности водоема.
Средняя плотность — это число особей (или биомасса) на единицу всего пространства, а специфическая, или экологическая, плотность — число особей (или биомасса) на единицу заселенного пространства (т. е. доступной площади или объема, которые фактически могут быть заняты популяцией). Как и многие другие показатели популяции, плотность варьирует в определенных переделах и эти изменения не бесконечны, различают верхние и нижние пределы. Верхний предел плотность популяции определяется потоком энергии в экосистемах, трофическим уровнем, а также объемом и интенсивностью его метаболизма. Нижний предел выделить труднее, особенно в стабильных экосистемах, в которых, действует механизмы, поддерживающие плотность организма в довольно ограниченных пределах.
При увеличении численности плотность популяции не возрастает лишь в случае ее расселения, расширения ареала.
Как биологическая единица, популяция обладает определенной структурой и функцией.
Экологическая структура популяции – это ее состояние на данный момент времени. Структуру популяции отражают такие ее показатели, как численность и распределение особей в пространстве, соотношение групп по полу, возрасту, их морфологические, поведенческие и другие особенности. Структура популяции формируется как на основе общебиологических свойств вида, так и под влиянием абиотических факторов среды.
Половая структура (половой состав) — соотношение особей мужского и женского пола в популяции. В популяциях различают однополовые и двуполовые структуры. Однополовая структура состоит из женских особей, и размножаются партеногенезом (размножение яйцеклетки без оплодотворения) и характерна для пчел, для споровых и семенных растений. Более распространены двуполовые популяции.
Половая структура свойственна только популяциям раздельнополых организмов. Теоретически соотношение полов должно быть одинаковым: 50 % от общей численности должны составлять мужские особи, а 50 % — женские особи. Фактическое соотношение полов зависят от действия различных факторов среды, генетических и физиологических особенностей вида.
Различают первичное, вторичное и третичное соотношения. Первичное соотношение — соотношение, наблюдаемое при формировании половых клеток (гамет). Обычно оно равно 1:1. Такое соотношение обусловлено генетическим механизмом определения пола. Вторичное соотношение — соотношение, наблюдаемое при рождении. Третичное соотношение — соотношение, наблюдаемое у взрослых половозрелых особей. Например, у человека во вторичном соотношении несколько преобладают мальчики, в третичном — женщины: на 100 мальчиков рождается 106 девочек, к 16—18 годам из-за повышенной мужской смертности это соотношение выравнивается и к 50 годам составляет 85 мужчин на 100 женщин, а к 80 годам — 50 мужчин на 100 женщин.
У некоторых рыб (р. Пецилия) различают три типа половых хромосом: Y, X и W, из них Y-хромосома несет гены мужского пола, а X и W-хромосомы — гены женского пола, но разной степени «мощности». Если генотип особи имеет вид YY, то развиваются самцы, если XY — самки, если же WY, то в зависимости от условий среды развиваются половые признаки самца или самки.
В популяциях меченосцев соотношение полов зависит от значения рН среды. При рН = 6,2 количество самцов в потомстве составляет 87-100 %, а при рН = 7,8 - от 0 до 15 %.
Для млекопитающих высших беспозвоночных характерно разнополовая система. Для млекопитающих (когда один самец оплодотворяет несколько самок) – важным условием является численность самок, а не общее количество особей т.к. лишь некоторые виды образуют на период размножения отдельную пару, которая сохраняется до конца жизни одного из партнеров.
Соотношение полов – это отношение количества самцов к количеству самок или количества самцов, к общей численности самцов и самок. Для характеристики используют запись в виде десятичной дроби или в соотношении в долях.
Возрастная структура (возрастной состав) — соотношение в популяции особей разных возрастных групп. Абсолютный возрастной состав выражает численность определенных возрастных групп в определенный момент времени. Относительный возрастной состав выражает долю или процент особей данной возрастной группы по отношению к общей численности популяции. Для каждого вида и популяции, характерны свои соотношения возрастных групп, на эти соотношения влияют общая продолжительность жизни, время достижения половой зрелости, интенсивность размножения, т.е. особенности вырабатывания в процессе эволюции как приспособление к определенным условиям.
В популяции обычно выделяют три экологических возраста: предрепродуктивный (особи еще не способные размножаться), репродуктивный (особи способные размножаться), пострепродуктивный (особи уже не способные размножаться). Потому в зависимости от соотношения количества особей относящихся к различным экологическим возрастам способность популяции к изменению численности будет не одинаково. Как правило, в быстрорастущих популяциях преобладают особи младших возрастных групп, в стабильных – распределение между возрастными группами относительно равномерно, а в популяциях с уменьшающейся численностью, преобладают особи старших, возрастов. Однако возрастная структура популяций может изменяться и без заметных колебаний общей численности: каждый пол имеет определенную возрастную структуру, к которой направлено изменение ее реальной возрастной структуры. Если по какой то причине произошло изменение состава структуры популяции, то через определенный промежуток времени она возвращается в исходное состояние. Для описания возраста структуры в популяции выделяют возрастные группы, состоящие из организмов одного возраста, и оценивают численность этих групп. Результаты представляют в виде диаграммы или пирамиды, выделяют три типа возрастных пирамид:
растущая популяция – пирамида отражает высокий процент молодых особей в популяциях.
стабильная популяция – пирамида отражает умеренное соотношение молодых и старых особей.
старая популяция – пирамида отражает низкий процент молодых особей.
Если в популяции размножение происходит постоянно, то по возрастной структуре устанавливают сокращается или увеличивается численность популяций. Если основание пирамиды широкое – это значит, что рождаемость превышает смертность и численность растет. Если же основание узкое, то молодых возрастных групп меньше, чем более старых и численность будет уменьшаться.
Возрастной состав связан с интенсивностью смертности и величиной рождаемости организма, который зависит от внешних факторов, которые контролируют мах рождаемость и смертность. По возрастному составу оценивается способность популяции к самоподдержанию численности и ее устойчивости к внешним фактором. Чем сложнее возрастной состав популяции, тем устойчивее ее воспроизводство. Анализ возрастного состава популяции позволяет прогнозировать численность популяции на ряд ближайших лет. Популяции, включающие в себя множество возрастных групп в меньшей степени подвержены влиянию факторов определяющих успешность размножения т.к. так как самые не благоприятные условия размножения способные приводить к полной гибели приплода того или иного года, не являются катастрофическими для популяций, потому, что одни и те же рождающие пары участвуют в воспроизводстве много раз.
Возрастные группы могут быть подразделены и на более мелкие категории. Например, у растений выделяют следующие состояния: покоящееся семя, проростки и всходы, ювенальное состояние, имматурное состояние, виргинильное состояние, раннее генеративное, среднее генеративное, позднее генеративное, субсенильное, сенильное (старческое), состояние полутрупа.
У растений-монокарпиков, включая однолетних, а иногда и у поликарпиков отсутствует постгенеративный период. Распределение особей ценопопуляции по возрастным состояниям называется ее возрастным спектром. Счетной единицей могут являться отдельные особи, парциальные кусты (длиннокорневые растения) или одиночные побеги (некоторые длиннокорневищные и корнеотпрысковые растения).
Ценопопуляцию, в возрастном спектре которой в момент наблюдения представлены только семена и молодые особи, называют инвазионной. Обычно это молодая ценопопуляция, только что внедрившаяся в фитоценоз. Поддержание ее обеспечивается заносом зачатков извне: Ценопопуляция называется нормальной, если она представлена всеми или практически всеми возрастными группами. Она способна к самоподдержанию вегетативным или семенным путями. Нормальной полночленной называют популяцию, которая состоит из особей всех возрастных групп. Если особи каких-либо возрастных состояний отсутствуют, например проростки или сенильные, то такая ценопопуляция называется нормальной неполночленной. Нормальные неполночленные ценопопуляции имеют монокарпики. Ценопопуляцию, не содержащую молодых особей, а представленную сенильными и субсенильными или даже цветущими особями, но не образующими семян, называют агрессивной. Регрессивная ценопопуляция не способна к самоподдержанию и зависит от заноса зачатков извне.
Возрастная структура ценопопуляции в значительной степени определяется биологическими особенностями вида: периодичностью плодоношения, числом продуцируемых семян и вегетативных зачатков, способностью вегетативных зачатков к омоложению, скоростью перехода особей из одного возрастного состояния в другое, длительностью возрастного состояния, способностью образовывать клоны и др.
У многих животных, так же как и у растений, более длительным является предрепродуктивный период. Так, у поденок (Ephemeridae) он продолжается несколько лет из-за длительного развития личинок. Репродуктивный же их возраст не превышает нескольких дней — время размножения взрослых особей. Пострепродуктивный период здесь практически отсутствует. Популяции быстро восстанавливают свою численность, если особи имеют короткий предрепродуктивный период.
B сокращающихся популяциях преобладают старые особи, которые уже не способны интенсивно размножаться. Данная возрастная структура свидетельствует о неблагоприятных условиях, В быстро растущих популяциях преобладают интенсивно размножающиеся молодые особи. В стабильных популяциях это соотношение, как правило, составляет 1:1. При благоприятных условиях в популяции имеются все возрастные группы, и поддерживается сравнительно стабильный уровень численности. На возрастной состав популяции помимо общей продолжительности жизни влияют длительность периода размножения, число генераций в сезон, плодовитость и смертность разных возрастных групп. Так, у полевок взрослые особи могут давать потомство 3 раза в год и более, а молодые особи способны размножаться через два—три месяца
Правило стабильности возрастной структуры популяции: любая естественная популяция стремится к стабильной возрастной структуре, четкому количественному распределению особей по возрастам. Это правило сформулировано А. Лоткой в 1925 г.
Пространственное распространение популяций
Пространственная структура — характер распределения особей в пределах ареала. Она зависит от особенностей окружающей среды и этологии (особенностей поведения) вида. Каждая популяция занимает пространство, обеспечивающее средствами к жизни лишь определенное число особей. При этом полнота использования имеющихся ресурсов зависит как от общей численности популяции, так и от пространственного размещения особей. Различают случайное, равномерное и неравномерное (групповое) распределение особей и их групп (рис. 1).
Случайное распределение в природе встречается редко; оно наблюдается в тех случаях, когда среда очень однородна, а организмы не стремятся объединиться в группы и выражается в неодинаковом, расстоянии между особями. Является результатом вероятностных процессов, однородности среды и слабых социальных связей между особями.
Равномерное распределение бывает там, где между особями очень сильна конкуренция или существует антагонизм. Характеризуется равным удалением каждой особи от всех соседних. Свойственно популяциям, существующим в условиях равномерного распределения факторов среды или состоящих из особей, проявляющих друг к другу антагонизм.
Чаще всего наблюдается неравномерное (групповое) распределение — образование различного рода скоплений. Проявляется в образовании группировок особей, между которыми остаются большие незаселенные территории. Характерно для популяций, обитающих в условиях неравномерного распределения факторов среды или состоящих из особей, ведущих групповой (стадный) образ жизни. Если особи в популяции стремятся образовывать группы определенного размера (например, пары у животных, вегетативные клоны у растений), то распределение групп может оказаться более близким к случайному.
Неравномерное распределение индивидуумов возникает:
1) вследствие местных различий условий среды и неравномерности занимаемого пространства,
2) под влиянием суточных и сезонных изменений погоды,
3) в связи с процессом размножения,
4) в результате взаимного притяжения (у высших животных).
Установление типа распределения, степени агрегированности, а также размеров и продолжительности сохранения групп необходимо для понимания природы популяции и, особенно для точного измерения ее плотности.
Способы, которыми достигается рациональный характер размещения, определяются системой взаимоотношений между членами популяции. Агрегация может усиливать конкуренцию между особями за питательные вещества, корм или жизненное пространство, однако это неблагоприятное следствие агрегации более чем уравновешивается тем, что она способствует выживанию группы в целом. По сравнению с одиночными особями у особей, объединенных в группу, наблюдается снижение смертности в неблагоприятные периоды или при нападении других организмов, поскольку в группе поверхность их соприкосновения со средой по отношению к массе меньше и поскольку группа способна изменять микросреду в благоприятном для себя направлении. Степень агрегированности (так же как и общая плотность), при которой наблюдается оптимальный рост и выживание популяции, варьирует в зависимости от вида и условий, поэтому как «недонаселенность» (или отсутствие агрегированности), так и перенаселенность могут оказывать лимитирующее влияние (так называемый принцип Олли).
Факторы, способствующие изоляции или разделению в пространстве особей либо мелких пар, составляющих популяцию, распространены не так широко, как факторы, содействующие агрегированности, но, тем не менее, они весьма важны, особенно для регуляции размеров популяций (посредством внутривидовых и межвидовых механизмов или же тех и других). Обычно изоляция возникает как следствие конкуренции между особями за пищу при ее недостатке или прямого антагонизма. В обоих случаях это может привести к случайному или равномерному распределению особей, поскольку близкие соседи уничтожаются либо изгоняются. Активность особей, пар или семейных групп у позвоночных и высших беспозвоночных обычно ограничена определенной зоной, называемой индивидуальным (или семейным) участком территории.
У высших животных внутрипопуляционное распределение регулируется системой инстинктов. Им свойственно особое территориальное поведение — реакция на местонахождение других членов популяции (у птиц, млекопитающих, ряда рыб, насекомых, крабов, осьминогов и др.). В наибольшей степени территориальность выражена у позвоночных и некоторых членистоногих, обладающих сложным репродуктивным поведением, проявляющимся, в частности, в гнездостроении, откладывании яиц, заботе о потомстве и его защите. Однако Ю. Одум включает в понятие территориальности любой активный механизм, посредством которого происходит разобщение в пространстве особей или групп особей. Так, свойственная растительным организмам изоляция уменьшает конкуренцию, способствует сохранению энергии в критические периоды, предотвращает перенаселенность, и истощение запасов пищи для низших животных и биогенных веществ, воды и света для растений. У животных благодаря их подвижности способы упорядочения территориальных отношений более разнообразны по сравнению с растениями. Территориальность благоприятствует регуляции численности популяции на уровне, более низком, чем уровень насыщения. В зависимости от характера использования пространства все подвижные животные разделяются на две основные группы: оседлые и кочевые.
Оседлые животные в течение всей или большей части жизни используют довольно ограниченный участок среды. Им присущи инстинкты привязанности к своему участку, регулярное возвращение к месту размножения после длительных и дальних миграций. Это свойство носит название хоминга (чувство дома). При этом у оседлых видов выработались такие приспособительные особенности поведения, которые обеспечивают «закрепление» мест обитания за определенными особями, семьями или другими внутрипопуляционными группировками (охрана границ занимаемого пространства от чужаков, ритуальное поведение типа угрозы, специальные предупредительные сигналы, маркировка территории запаховыми, зрительными и другими метками). Территориальное поведение животных особенно проявляется в период размножения.
Кочевые животные совершают постоянные передвижения в пространстве, так как они зависят от запаса корма на конкретной территории. Кочевой образ жизни характерен преимущественно для стад и стай (кочующие группы многих рыб во время нагульных миграций, стада слонов, зебр, антилоп, северных оленей и т. п.). Перемещение кочующих видов по площади обычно происходит за сроки, достаточные для восстановления кормовых ресурсов на пастбищных участках. В целом при кочевом образе жизни животные совершают более или менее закономерные циклические перемещения по обширной территории с повторным использованием одних и тех же участков через определенные интервалы. Масштабы и длительность таких миграций обусловлены обилием пищи и численностью стада.
По типу использования пространства все подвижные животные подразделяются на оседлых и кочевых. Оседлый образ жизни имеет ряд биологических преимуществ, таких как свободная ориентация на знакомой территории при поиске пища или укрытия, возможность создать запасы пищи (белка, полевая мышь). К его недостаткам относится истощение пищевых ресурсов при излишне высокой плотности популяции.
По форме совместного существования животных выделяют одиночный образ жизни, семейный, колониями, стаями, стадами. Одиночный образ жизни проявляется в том, что особи в популяциях независимы и обособленны друг от друга (ежи, щуки и др.). Однако он характерен только для определенных стадий жизненного цикла. Полностью одиночное существование организмов в природе не встречается, так как при этом было бы невозможно размножение. Семейный образ жизни наблюдается в популяциях с усилением связей между родителями и потомством (львы, медведи и др.). Колонии — групповые поселения оседлых животных, как длительно существующие, так и возникающие лишь на период размножения (гагары, пчелы, муравьи и др.). Стаи — временные объединения животных, облегчающие выполнение какой-либо функции: защиты от врагов, добывания пищи, миграции (волки, сельдь и др.). Стада — более длительные, чем стаи, или постоянные объединения животных, в которых, как правило, выполняются все жизненные функции вида: защита от врагов, добывание пищи, миграции, размножение, воспитание молодняка и т.д. (олени, зебры и др.).
Этологическая структура популяций. Характерной особенностью популяций животных является система взаимоотношений между составляющими ее членами. Закономерности поведения организмов изучает наука этология. В зависимости от образа жизни вида формы совместного существования особей в популяции чрезвычайно разнообразны. Различают одиночный образ жизни, при котором особи популяции независимы и обособлены друг от друга, но лишь временно, на определенных стадиях жизненного цикла. Полностью одиночное существование организмов в природе не встречается, так как было бы невозможным осуществление их основной жизненной функции — размножения. У видов с одиночным образом жизни часто образуются временные скопления особей в местах зимовок (бабочки-крапивницы, жужелицы, божьи коровки) либо в период, предшествующий размножению. В последнем" случае у многих видов возникают ритуальные демонстрации, драки самцов и проявляются другие типы специализированного поведения, направленного на устранение конкурентов.
При семейном образе жизни заметно усиливаются связи и взаимоотношения между родителями и их потомством: забота об отложенных яйцах, молодых птенцах, охране мальков. Различают семьи отцовского, материнского и смешанного типов. При семейном образе жизни начинает заметно проявляться территориальное поведение животных, когда путем различных сигналов, маркировкой, формами угрозы и т. п. обеспечивается владение участком, достаточным для выкармливания потомства. В основе формирования более крупных объединений животных (стая, стадо, колония) лежит дальнейшее усложнение поведенческих связей в популяциях.
Стая — это временное объединение животных, которые проявляют биологически полезную организованность действий (для защиты от врагов, добычи пищи, миграции и т. п.). Наиболее широко стайность распространена среди рыб и птиц, встречается и у млекопитающих (собачьи стаи). В стаях сильно развиты подражательные реакции и ориентация на соседей.
Стадо — длительное или постоянное объединение животных, в котором осуществляются все основные функции жизни вида: добывание корма, защита от хищников, миграции, размножение, воспитание молодняка. Основу группового поведения в стадах составляют взаимоотношения доминирования — подчинения, базирующиеся на индивидуальных различиях между особями. Для стада характерно наличие временного либо относительно постоянного лидера, на котором концентрируется поведение других особей и которые, подражая лидеру, своим поведением определяют поведение стада в целом. В отличие от лидера вожаки характеризуются поведением, непосредственно направленным на активное руководство стадом (специальные сигналы, угрозы или прямое нападение).
В крупных стадах (стада диких оленей, зебр, павианов и др.) выделяют также объединения, представляющие собой семейные или возрастные группы со своим внутренним иерархическим соподчинением особей разных рангов. Иерархически организованному стаду свойственен закономерный порядок перемещения, определенные позиции при защите от врагов, расположении на местах отдыха и т. п.
Колония — это групповое поселение оседлых животных. Колонии могут существовать длительно или возникать лишь на период размножения. По сложности взаимоотношений между особями колонии животных чрезвычайно разнообразны. Это могут быть:
а) простые территориальные скопления одиночных форм (морские желуди, мидии);
б) более сложные поселения, в которых некоторые функции жизни животных выполняются сообща (защита от врагов и предупредительная сигнализация в колониях чаек, кайр, ласточек, грачей, альбатросов);
в) наиболее сложный тип поселений, характерный для общественных насекомых — термитов, муравьев, пчел, ос и др. Такие колонии возникают на основе сильно разросшейся семьи. При этом насекомые выполняют сообща большинство основных функций: размножения, защиты, обеспечения кормом себя и потомства, строительства и т. п. с обязательным распределением труда и специализацией особей или возрастных групп на выполнении определенных операций. Члены колонии действуют на основе постоянного обмена информацией друг с другом.
Динамика популяций
Популяции изменчивы во времени при ее изменении особый интерес представляет не только ее состав, но и величина в каждый определенный момент времени. Зная скорость изменения популяции можно судить об особенностях и условиях развития, а так же о состоянии экосистемы в целом. Скорость изменения числа популяции можно определить путем деления величины изменения количества особей на период за которое оно произошло. Полученная величина будет характеризовать скорость в которой численность изменяется во времени. Средняя скорость изменения популяций
V =
![]()
Где - N – размер популяций;
t – время за которое изменяется популяция;
Н – популяция из 100 особей в некоторых объемах воды за 1 ч увеличивается до 150 особей.
ΔН = 50
V = 50 особей/час
Δt = 1 ч
Выделяют 4 типа динамики численности популяции:
Показательный – интенсивный рост в форме геометрической прогрессии и может происходить от биологических особей организма и от условий места обитания. После достижения максимума количество особей уменьшается.

Логистический – характеризует популяцию, которая только заселяет территорию популяция проходит сразу фазу роста, до момента овладения средой, затем переходит в стадию равновесия.




Циклический – характеризует динамику численности популяций, которая регулярно повторяется под влиянием циклических изменений в среде.



Стабильный – характеризует популяции, которые развиваются в постоянных условиях среды и имеют отлаженный механизм регулирования численности.


Экспериментальные исследования позволили выделить три фазы роста численности популяций:
а) фаза роста характеризуется приростом общей численности популяции на протяжении определенного периода времени.
б) фаза спада характеризуется снижением числа особей за определенный период времени.
в) фаза равновесия характеризует периодом времени, когда средняя численность популяции остается на одном уровне.
Рост популяции характеризуется рождаемостью, смертностью миграций (эмиграцией - иммиграцией)
Рождаемость. Это показатель, характеризующий способность популяции к увеличению численности в результате размножения особей или просто скорость размножения.
Т.о. рождаемость показывает число новых особей в популяции. Рождаемость выразить в относительных показателях, т.е. числом родившихся особей за единицу времени, отнесенная к общей численности популяции в начале сезона размножения. Когда жизнь популяции протекает в благоприятных условиях, и отсутствуют факторы, ограничивающие доступность используемых ресурсов, рождаемость повышается, результатом этого может быть быстрое, но кратковременное увеличение численности популяций. Рождаемость в это время мах величина. Мах рождаемости определяется числом самок в популяции и их способностью производить определенное количество детенышей за единицу времени. Т.е. физиологической плодовитостью в дальнейшем по мере роста численности популяций, условия жизни ухудшаются, а скопление особей, препятствует эффективному использованию ресурсов. Накапливаются вредные отходы, рост численности замедляется и в конечном итоге она достигает уровня, который соответствует сложившимся экологическим условиям, одновременно происходит снижение рождаемости, которая обеспечивает лишь поддержание численности популяции, такая рождаемость называется экологической. Т.о. мах (абсолютная) рождаемость это появление теоретически Мах возможного количества особей в идеальных условиях. Эта величина является постоянной для конкретной популяции. Реализуемая рождаемость обозначает увеличение популяции при фактических или специфических условиях среды. Она зависит от состава размера популяции и фактическим условиям среды. Рождаемость обычно выражается в виде скорости определяемой путем деления числа новых особей за определенный период времени.
N∕∆T
Абсолютную рождаемость выражают в расчете на 1 самку, для человека в расчете на 1000 чел рождаемость может быть нулевой или положительной, но не отрицательной. Мах рождаемость использует, как критерий для сопоставления с реальной рождаемостью.
Смертность. Это количество особей, погибших за определенный период, различают специфическую смертность число смертей по отношению к числу особей в популяции. Экологическая (реализуемая) – это гибель особей в определенных условиях среды. Величина непостоянная изменяется в зависимости от состояния природной среды и состояния популяции. Существует тин величина, характеризующая гибель особей в идеальных условиях, когда на популяцию воздействует лимитирующие факторы. В этих условиях мах продолжительность жизни равна их физиологической жизни. Специфическая смертность обычно выражается числом особей, погибших за определенное время и выражено в % от начальной численности популяции тин смертность принимают за теоретическую константу т.к. даже в идеальных условиях в любой популяции происходит смертность от старости или связанные с дефектами развития. В конкретных экологических условиях смертность выше минимальной т.к. влияние внешних факторов, хищники, отсутствие корма, загрязненность среды, вызывают дополнительные причины гибели организма.
Выделяют три типа смертности:
1 тип характеризуется одинаковой смертностью во всех возрастах. Данный тип встречается редко и только у популяций, находящих в оптимальных условиях.
2 тип характеризуется повышенной гибелью особей на ранних стадиях развития. Характерен для большинства животных и растений мах гибель у животных в личиночной стадии и в молодом возрасте, у растений – на стадии произрастания семян и всходов.
3 тип характеризуется повышенной гибелью взрослых особей и в первую очередь старых. В природе средняя продолжительность жизни особей намного меньше, чем потенциальная и фактическая величина смертности выше минимальной.
Кривые выживания
На основе данных о рождаемости и смертности в популяциях строят кривые выживания. По оси абсцисс отмечают процент выживания (продолжительность жизни от общей продолжительности) и по оси ординат (время или скорость).
Рис. Кривые выживания (по Ф. Дрё, 1976):
I — кривая дрозофилы; II — кривая гидры; III — кривая устрицы
характерна для сильных организмов смертность низкая, высокая смертность в конце жизни, характерно для крупных видов животных и человека
редкая смена выживания в разном возрасте и разных фазах развития, характерна для насекомых.
постоянное выживание, в каком либо возрасте реально осуществима и может только приближаться к идеальной кривой оси.
сильно вогнутая – наибольшая смертность в начале жизни, а в дальнейшем низкая (для растений гибель семян)
Понижающая при ускорении форма кривой выживания связанна со степенью заботы о потомстве и другими способами защиты молодых особей. Форма кривой выживания, часть изменяется при изменении плоти популяции и при возрастании плотности она становится более вогнутой, при увеличении численности организмов их смертность возрастает.
Экологические стратегии популяции. Приспособление особей в популяции в конечном счете направлены на повышение выживания и оставления потомства. Экологическая стратегия популяций это общая характеристика ее роста и размножения сюда входят темпы роста особей, время достижения половой зрелости, плодовитости, периодичность размножения вида и занимающие нарушения места обитания и имеющие большое значение ч-видами или виды оппортунисты. Виды с относительно низким значением ч и для в которых скорость размножения чувствительность к плотности популяции называется К-видами. Характерные особенности представлены в таблице:
r |
К |
Различают быстро (высокая плодовитость, время генерации короткое) |
Размножение медленно низкая плодовитость продолжительность времени генерации |
V размен не зависит от плотности популяции |
V размен зависит от плотности популяций если р падает V быстро увеличивается |
Энергия и вещество распределены между многими потомками |
Энергия и вещество концентрируются в немногих потомках, родители заботятся о потомстве |
Размеры популяций некоторое время могут превышать поддерживающую емкость среды |
Размеры популяций близки к равномерному уровню |
Вид не всегда устойчив на данной территории |
Вид устойчив на данной территории |
Расселяются широко и в больших количествах |
Расселяются медленно |
Размножение идет с большой затратой энергии и вещества |
Размножение идет с относительно малыми затратами, большая часть энергии расходуется на вегетативный рост |
Малые размеры особей |
Крупные размеры, у деревянных стеблей и большие корни |
Малая продолжительность жизни, могут рассеяться на открытых местах, место обитания сохраняется не долго |
Большая продолжительность жизни плохо приспособлены к росту на открытых местах , место обитания устойчивое и сохраняется долго |
Слабые конкуренты, слабо развиты защитные приспособления, лучше приспособлены к изменению окружающей среды (бактерии, тля, мучные хрущаки, однолетние растения) |
Сильные конкуренты хорошие защитные механизмы, менее устойчивы к изменениям условий среды (крупные тропические бабочки, кондор, альбатрос, человек, деревья) |
Виды с r- стратегией быстрее заменяют нарушенное местообитания (горная порода, лесные вырубки, выгоревшие участки леса) Они занимают данное место обитания в течение жизни одного или нескольких поколений, в дальнейшем они переселяются на новое место. Виды с К-стратегией более конкурентно способные и обычно вытесняют К-виды. Могут использовать одно и то же место обитания по разному, то они могут сосуществовать вместе.
СТРУКТУРА И ДИНАМИКА ПОПУЛЯЦИЙ
Каждый вид занимает тот или иной ареал, в различных частях которого наблюдаются разные условия. В связи с этим Н. П. Наумов предложил концепцию иерархии популяций в зависимости от размеров занимаемой ими территории:
элементарная (локальная) популяция — совокупность особей вида, занимающих небольшой участок однородной площади. Количество таких популяций пропорционально разнообразию условий биогеоценоза;
экологическая популяция — совокупность элементарных популяций, т. е. внутривидовые группировки, приуроченные к конкретным биогеоценозам;
географическая популяция — совокупные группы особей предыдущего ранга, заселяющие значительную территорию с географически однородными условиями существования, в пределах которой наблюдаются единый ритм жизненных явлений и другие функциональные особенности, создающие морфофизиологический тип, отличающий данную популяцию от соседних, находящихся в иных географических условиях.
Соотношение особей по полу в популяции (особенно доля размножающихся самок) имеет большое значение для дальнейшего роста ее численности. Генетический механизм обеспечивает примерно равное соотношение образующихся зигот по признаку пола, которое вскоре заметно нарушается в результате различий физиологических, поведенческих, экологических реакций самцов и самок, приводящих к неравномерной смертности.
Внутрипопуляционные процессы. Рождаемость и смертность в популяциях
Рождаемость (плодовитость) — число новых особей, появившихся за единицу времени в результате размножения. Средняя величина плодовитости каждого вида определилась исторически как приспособление, обеспечивающее восполнение убыли популяций.
Огромная потенциальная возможность к размножению никогда не реализуется организмами вследствие гибели значительного числа яиц, личинок, влияния болезней, хищников и т. п. Максимальная рождаемость (иногда ее называют абсолютной, или физиологической) — образование теоретически максимально возможного количества новых особей в идеальных условиях (когда отсутствуют лимитирующие экологические факторы и размножение ограничивается только физиологическими факторами); для данной популяции это величина постоянная.
Экологическая рождаемость — увеличение популяции при фактических или специфических условиях среды. Эта величина изменяется в зависимости как от размера и состава популяции, так и от сопутствующих факторов.
Смертность характеризует гибель особей в популяции. Как рождаемость выражается числом новых особей, так смертность отражает число особей, погибших за данный период, по отношению к числу особей, составляющих всю популяцию или ее часть. Экологическая или реализуемая, смертность — гибель особей в данных условиях среды, величина, которая, подобно экологической рождаемости, не остается постоянной, а изменяется в зависимости от условий среды и состояния самой популяции. Существует некоторая теоретическая (минимальная смертность) — постоянная величина, характеризующая гибель особей в идеальных условиях, при которых популяция не подвергается лимитирующим воздействиям.
Максимальная продолжительность жизни особей в этих оптимальных условиях равна их физиологической продолжительности жизни, которая в среднем обычно намного превышает экологическую.
Специфическую смертность обычно выражают как процент числа особей, погибших за данное время, от начальной численности популяции.
На численные показатели рождаемости и смертности популяций существенное влияние оказывают эмиграция (выселение), иммиграция (пополнение) и общая миграция особей. Обычно первые два из названных потоков мигрантов уравновешивают друг друга, но массовые перемещения особей между популяциями могут изменить их структуру и основные свойства (предотвратить гибель популяции, находящейся на грани вымирания, либо, наоборот, привести ее к катастрофическому сокращению). Миграции (суточные, сезонные) позволяют организмам использовать оптимальные условия среды в таких местах, где постоянное проживание их невозможно. Немигрирующие виды с наступлением неблагоприятного периода обычно переходят в неактивное состояние (диапауза, зимняя спячка у животных, состояние покоя у растений).
Возрастная структура популяций. В каждой популяции есть группы разновозрастных особей, соотношение которых характеризует способность популяции к размножению. Чем сложнее такая структура, тем выше приспособительные возможности популяции.
По отношению к популяции можно выделить три экологических возраста: предрепродуктивный, репродуктивный и пострепродуктивный. Длительность этих возрастов относительно продолжительности жизни сильно варьирует у разных организмов. У многих животных и растений особенно длительным бывает предрепродуктивный период. При благоприятных условиях в популяции присутствуют все возрастные группы, обеспечивая относительно стабильный уровень ее численности. На возрастной состав популяции влияют продолжительность жизни особей, период достижения половой зрелости, длительность периода размножения, количество генераций или приплодов в сезон, плодовитость и смертность разных возрастных групп.
Наиболее устойчивая структура популяции возникает при длительном сосуществовании различных возрастных групп. При этом размножающаяся часть особей составляет «запас» популяции. Учет и анализ возрастной структуры природных популяций имеют большое значение для рационального охотничьего промысла и прогнозирования популяционно-экологической ситуации.
Типы роста популяций. Характер увеличения численности популяции может быть неодинаковым, и в связи с этим выделяют различные типы роста популяций. По форме кривых роста, построенных на арифметической шкале, можно выделить два основных типа роста:
- рост, описываемый т-образной кривой,
- рост, описываемый S-образной кривой.
Эти два противоположных типа роста в соответствии с особенностями разных организмов и условий среды могут различным образом сочетаться и модифицироваться. При т-образной форме кривой роста плотность быстро увеличивается по экспоненте, но затем, когда начинает действовать (обычно более или менее резко) сопротивление среды или другой лимитирующий фактор, рост популяции внезапно прекращается (рис 3).
Рис. 3. Динамика роста численности (N) одной и той же гипотетической популяции микроорганизмов при использовании линейного (а) и логарифмического (б) масштабов
При S-образной форме кривой роста увеличение плотности популяции вначале происходит медленно (начальная фаза, или фаза положительного ускорения), затем быстрее (эту фазу называют фазой приближения к логарифмическому росту), но вскоре под влиянием сопротивления среды рост популяции постепенно замедляется (фаза отрицательного ускорения). По мере увеличения сопротивления среды замедление роста становится все более выраженным, и в конце концов достигается и поддерживается более или менее устойчивое равновесие.
Лекция 8 - Экологические стратегии популяций. Сравнительная характеристика г- и К- стратегий
Экологическая стратегия выживания — комплекс свойств популяции, направленных на повышение вероятности выживания и оставление потомства, называется. Это общая характеристика роста и размножения. Сюда входят темпы роста особей, время достижения половозрелости, плодовитость, периодичность размножения и т.д.
Так А.Г. Раменский (1938) различал три основные типа стратегий выживания среди растений: виоленты, патиенты и эксплеренты.
Виоленты (силовики) — подавляют всех конкурентов, например, деревья, образующие коренные леса.
Патиенты — виды, способные выжить в неблагоприятных условиях («тенелюбивые», «солелюбивые» и т.п.).
Эксплеренты (наполняющие) — виды, способные быстро появляться там, где нарушены коренные сообщества, — на вырубках и гарях (осины), на отмелях и т.д.
Экологические стратегии популяций отличаются большим разнообразием. Но при этом все их многообразие заключено между двумя типами эволюционного отбора, которые обозначаются константами логистического уравнения: r-стратегия и К-стратегия.
г-стратеги (r-виды, г-популяции) — популяции из быстро размножающихся, но менее конкурентоспособных особей. Имеют j-образную кривую роста численности, не зависящую от плотности популяции. Такие популяции быстро расселяются, но они малоустойчивы. К ним относятся бактерии, тли, однолетние растения и др. (табл. 6).
К-стратеги (К-виды, К-популяции) — популяции из медленно размножающихся, но более конкурентоспособных особей. Имеют S-образную кривую роста численности, зависящую от плотности популяции. Такие популяции населяют стабильные местообитания. К ним относятся человек, кондор, деревья и др. Следует отметить, что одну и ту же среду обитания разные популяции могут использовать по-разному, поэтому в одном и том же местообитании могут сосуществовать виды с r- и К-стратегиями. Между этими крайними стратегиями существуют переходы. Ни один из видов не подвержен только r- или только К- отбору
Гомеостаз популяции — поддержание определенной численности (плотности). Изменение численности зависит от целого ряда факторов среды — абиотических, биотических и антропогенных. Однако всегда можно выделить ключевой фактор, наиболее сильно влияющий на рождаемость, смертность, миграцию особей и т.д.
Факторы, регулирующие плотность популяции, делятся на зависимые и независимые от плотности.
Зависимые от плотности факторы изменяются вместе с изменением плотности, к ним относятся биотические факторы.
Независимые от плотности факторы остаются постоянными с изменением плотности, это абиотические факторы.
Популяции многих видов организмов способны к саморегуляции своей численности. Выделяют три механизма торможения роста численности популяций:
при возрастании плотности повышается частота контактов между особями, что вызывает у них стрессовое состояние, уменьшающее рождаемость и повышающее смертность;
при возрастании плотности усиливается эмиграция в новые местообитания, краевые зоны, где условия менее благоприятны и смертность увеличивается;
при возрастании плотности происходят изменения генетического состава популяции, например, быстро размножающиеся особи заменяются медленно размножающимися.
Понимание механизмов регуляции численности популяций чрезвычайно важно для возможности управления этими процессами. Деятельность человека часто сопровождается сокращением численности популяций многих видов. Причины этого в чрезмерном истреблении особей, ухудшении условий жизни вследствие загрязнения окружающей среды, беспокойства животных, особенно в период размножения, сокращение ареала и т.д. В природе нет и не может быть «хороших» и «плохих» видов, все они необходимы для ее нормального развития.
Типы стратегии жизни (поведения) организмов. Типы стратегии жизни (поведения) организмов — это самая важная оценка экологии вида, интегральная характеристика, которая отражает и жизненные циклы, и жизненные формы, и экологические группы. Для каждого типа стратегии характерен свой комплекс (синдром) адаптивных признаков.
«r-отбор» и «К-отбор». Слово «стратегия», исходно обозначающее определенную систему спланированных военных действий, пришло в экологию во второй половине XX столетия, причем первоначально говорили только о стратегии поведения животных.
П. Макартур и Е. Уилсон (Macаrthur, Wilson, 1967) описали два типа стратегий организмов как результаты двух типов отбора, связанных отношениями трейдоффа:
r-отбор — эволюция в направлении увеличения затрат на размножение организма, итогом которой являются г-стратеги; К-отбор — эволюция в направлении увеличения затрат на поддержание жизни взрослого организма, ее итогом являются К-стратеги.
Популяции К-стратегов, крупных организмов, живущих в стабильных «предсказуемых» условиях, имеют достаточно постоянный показатель численности, и среди взрослых особей наблюдается интенсивная конкуренция, на противодействие которой (т. е. на выживание) затрачивается основная доля ресурсов. Влияние конкуренции испытывают на себе и молодые особи, однако оно ослаблено, так как у животных - К-стратегов, как правило, отмечается забота родителей о потомстве, количество которого ограничено (слон, лев, тигр и др.).
Популяции г-стратегов состоят из мелких организмов с высоким вкладом в размножение, они формируются в «непредсказуемых» флюктуирующих условиях (домовая мышь, рыжий таракан, домашняя муха и др.). Периоды быстрого роста этих популяций при избытке ресурсов и слабой конкуренции чередуются с периодами «кризисов», когда количество ресурсов резко уменьшается. По этой причине размер таких популяций зависит в первую очередь от количества ресурсов и потому колеблется вне связи с конкуренцией. У г-стратегов — короткий жизненный цикл, позволяющий им успеть дать потомство до наступления очередного «кризиса», и специальные приспособления для переживания «кризисов» в покоящемся состоянии.
Э. Пианка (1981), рассматривая типы стратегий Макартура - Уилсона, подчеркнул, что «мир не окрашен только в черное и белое» и в природе преобладают организмы с переходными между г- и К-типами стратегий. У таких организмов отмечается некоторый компромисс между полярными составляющими трейдоффа, однако не существует организмов со стратегией, включающей целиком синдромы К-стратегов и г-стратегов («нельзя быть одновременно салатом и кактусом»).
У Макартура - Уилсона было, по крайней мере, два независимых и неизвестных этим ученым предшественника, имевших такие же взгляды.
Во-первых, о принципах дифференциации эволюции в направлениях поддержания организмами собственного существования и «продолжения себя в потомках» писал Г. Спенсер (1870). При этом Спенсер рассматривал данные направления эволюции как антагонистические, т. е. как трейдофф. В качестве примеров результатов такой эволюции он рассматривал слона и мелких животных.
Во-вторых, предтечей системы К- и г-стратегов был и ботаник Дж. Маклиод (McLeod, 1884, по Hermy, Stieperaere, 1985), который разделил растения на «пролетариев» и «капиталистов». (Разумеется, столь экстравагантные названия для типов были данью моде - именно в этот период в Европу пришел марксизм, тем не менее аналогии Маклиода весьма удачны).
Растения-капиталисты затрачивают основную энергию на поддержание взрослых особей, они уходят зимовать с капиталом из фитомассы многолетних тканей — древесных стволов и ветвей, корневищ, клубней, луковиц и пр. Растения-пролетарии, напротив, зимуют в стадии семян, т.е. без капитала, так как энергия в основном затрачивается на размножение. Это — однолетники, которые образуют большое количество семян и выживают за счет того, что всегда какая-то их часть попадает в благоприятные условия. Кроме того, «пролетарии» имеют семена, способные формировать почвенные банки, в которых подолгу сохраняют всхожесть и годами ждут «своего часа».
Растения с переходным типом стратегии, например многолетние луговые травы, характеризуются достаточно высокой плодовитостью и умеренной долей зимующих органов.
Система типов стратегий Раменского—Грайма. Выдающийся российский эколог Л.Г. Раменский (1935) разделил все виды растений на три «ценотипа» (к тому времени термин «стратегия» еще не вошел в обиход экологов): виолентов, патиентов и эксплерентов, дав им емкие образные эпитеты — «львы», «верблюды», «шакалы».
Работа Раменского осталась незамеченной не только за рубежом, но даже в России. На долю переоткрывшего те же типы стратегий Дж. Грайма (Grime, 1979), напротив, выпал грандиозный успех. При этом если Раменский описал свою систему все го на нескольких страницах, то Грайм посвятил ей две объемистые монографии (Grime, 1979; Grime et al., 1988). Сегодня эту систему стратегий называют «система Раменского - Грайма».
В отличие от одномерной системы г- и К-стратегов, система Раменского - Грайма двумерна и отражает отношение организмов к двум факторам: обеспеченности ресурсами (суммарным отражением действия этого комплексного градиента является биологическая продукция, см. разд. 10.6) и нарушениям. Нарушением является результат действия любого внешнего по отношению к экосистеме фактора, который вызывает разрушение ее части или уничтожает ее целиком. Факторами нарушения являются интенсивный выпас скота (особенно в лесу), распашка целинной степи, проход тяжелой техники в тундре и т. д. Нарушения в масштабе сотен квадратных километров могут вызвать землетрясения, извержения вулканов, большие лесные пожары, кислотные дожди.
Эта система типов стратегий изображается в виде «треугольника Грайма» (рис. 1). Буквы в углах треугольника обозначают три первичных типа стратегии, сочетания из двух и трех букв - переходные (вторичные) типы. Несмотря на «растительное» происхождение, систему стратегий Раменского - Грайма успешно используют не только ботаники, но и зоологи, и микробиологи.
Рис. 1. Треугольник Грайма (пояснения в тексте)
Первичные типы стратегий как и г- и К-стратегии. Первичные типы стратегий Раменского – Грайма как г- и К-стратегии связаны отношениями трейдоффа, т.е. синдромы их адаптивных признаков альтернативны.
Тип С (от англ. competitor — конкурент) — виолент, «силовик», «лев». Это мощные организмы, затрачивающие большую часть энергии на поддержание жизни взрослых особей, интенсивность размножения низкая.
Растения-виоленты — чаще деревья (бук, дуб), реже кустарники или высокие травы (например, канареечник в прирусловой пойме рек умеренной полосы или тростник в дельтах южных рек полупустынной и пустынной зон), которые произрастают в благоприятных условиях (полное обеспечение водой, элементами питания, теплый климат) при отсутствии нарушений. Они имеют распростертую крону (или корневища, как у канареечника и тростника), за счет которой держат под контролем условия среды и полностью (или почти полностью) используют обильные ресурсы таких местообитаний.
Виоленты всегда абсолютно доминируют в сообществах, и примесь других видов растений незначительна. В буковых лесах под пологом деревьев сумрачно и почти нет трав и кустарников. В зарослях тростника в дельте Волги биомасса доминанта составляет 99%, другие виды встречаются единично.
При ухудшении условий (просыхание почвы, засоление и т. д.) или их нарушении (рубка леса, высокие рекреационные нагрузки, пожары, воздействие техники и т. д.) «львы» растительного мира погибают, не имея приспособлений для переживания действия этих факторов.
Тип S (от англ. stress-tolerant — устойчивый к стрессу) — патиент, «выносливей», «верблюд». Это разнообразные организмы, способные за счет специальных адаптации переживать сильный стресс. Растения-патиенты обитают при дефиците ресурсов или наличии условий, которые ограничивают их потребление (засуха, засоление, дефицит света или ресурсов минерального питания, холодный климат и т. д.).
Не менее разнообразен арсенал адаптации растений к переживанию стресса дефицита элементов почвенного питания. Патиенты-олиготрофы имеют многолетние листья, питательные вещества из которых переходят в стебель перед их опаданием (пример - брусника). У сфагнового мха, обладающего способностью к бесконечному росту вверх, элементы питания постоянно перекачиваются из отмирающей части в живые стебли и листья. Патиентами являются почти все лишайники.
Адаптации растений к дефициту света — более тонкие, темно-зеленые листья, в которых содержание хлорофилла выше, чем в листьях растений, обитающих в условиях хорошего освещения.
Растения-патиенты не образуют сомкнутых сообществ, обычно их покров разрежен и число видов в этих сообществах небольшое. В некоторых сообществах патиенты сообитают с виолентами, занимая ниши под их густым пологом, например копытень в широколиственном лесу или мхи в еловом лесу.
Тип R (от лат. ruderis - сорный) - эксплерент, рудерал, «шакал». Эти организмы замещают виолентов при сильных нарушениях местообитаний или используют ресурсы в стабильных местообитаниях, но в периоды, когда они оказываются временно не востребованными другими видами.
Большинство растений-эксплерентов — однолетники (реже - двулетники), образующие большое количество семян (т. е. виды-«пролетарии», в терминологии Маклиода, или г-стратеги, по МакАртуру и Уилсону). Они способны формировать банк семян в почве (например, виды родов полынь, марь, лебеда) или имеют приспособления для распространения плодов и семян (например, летучки - у одуванчика, бодяка или прицепки — у липучки и лопуха, плоды которых разносят животные и человек).
Таким образом, растения-рудералы первыми начинают восстанавливать растительность при нарушениях: семена одних видов уже имеются в почвенном банке, семена других быстро доставляются на место нарушения ветром или иными агентами. Эту важную для экосистем группу растений можно сравнить с «ремонтной бригадой», которая, как живица на пораненном стволе сосны, залечивает нанесенные природе раны.
К эксплерентам относятся и виды, которые периодически дают вспышки обилия в стабильных сообществах без нарушений. Это происходит в двух случаях:
при обильных ресурсах, когда временно ослаблено конкурентное влияние постоянно обитающих в сообществах виолентов (весенние эфемероиды в лесах, которые развиваются до распускания листвы на деревьях);
при постоянно ослабленном режиме конкуренции и внезапно резко увеличивающемся количестве ресурса, который патиенты, постоянно присутствующие в сообществе, не могут освоить. В пустыне однолетники-эфемеры за короткий период вегетации после дождей покрывают поверхность почвы зеленым ковром.
Вторичные типы стратегий. Пластичность стратегий. Многим видам присущи вторичные стратегии, т. е. они, сочетают признаки синдромов двух или трех первичных типов стратегий. Однако поскольку синдромы виолентности, патиентности и эксплерентности связаны трейдоффом, а величина «суммарного адаптивного потенциала» ограничена, ни один вид с вторичной стратегией не может обладать полным набором признаков двух и тем более трех первичных стратегий (это напоминает ситуацию с фондовым портфелем: в его составе могут быть акции одной или нескольких компаний, но их общая стоимость определяется величиной капитала).
Видов растений с вторичными типами стратегий больше, чем видов с первичными типами стратегий. Примером вида, имеющего стратегию виолент-патиент (CS), является сосна, которая хорошо растет на бедных песчаных почвах, а также все виды рода ель, которые произрастают в холодном климате на бедных кислых (но хорошо увлажненных) почвах.
Стратегию виолент-рудерал (CR) имеют такие виды, как ольха серая (Alnus incana), которая разрастается на вырубках, и крапива двудомная — обычный доминант почв, богатых азотом. Виды со стратегией рудерал-патиент (RS) можно наблюдать на вытоптанных площадках вокруг колодцев в пустынной зоне (например, виды из рода Peganum).
Большинство луговых и степных растений представляют смешанный тип стратегии — CRS, т.е. сочетают в своем поведении черты виолентности, патиентности и эксплерентности, хотя эти качества у различных видов представлены в разном соотношении. Например, у видов солончаковых лугов - ячменя короткоостого {Hordeum brevisubulatum), бескильницы расставленной {Puccinellia distans) или типичных доминантов степей — ковылей и типчака — больше признаков патиентности, а у пырея ползучего — эксплерентности.
Многие виды обладают свойством пластичности стратегии. Например, дуб черешчатый в местообитаниях с оптимальными условиями — типичный виолент, а у южной границы ареала он представлен кустарниковой формой и является патиентом. Патиентом на засоленных почвах является тростник, который в этих условиях представлен стелющейся формой с узкими листьями. В плавнях дельт южных рек (Волга, Дон, Днепр, Урал) в условиях изобилия элементов минерального питания и теплого климата этот же вид имеет стратегию настоящего виолента, его высота достигает 3 и даже 4 м, а ширина листа порядка 3-4 см.
Японское искусство выращивания карликовых деревьев («бонсай») основано на превращении виолентов в патиенты. Природный «бонсай» создается из сосны на верховых болотах. На сфагновых кочках растут сосенки (Pinus sylvestris forma pumilis Abolin), которые в возрасте 90—100 лет имеют высоту меньше метра и диаметр «ствола» в пределах 5—8 мм, а длину хвои — 1 см. На таких «деревьях» образуются шишки со всхожими семенами (иногда на одном «дереве» — всего одна шишка).
Особенности стратегий культурных растений и животных. Сельское хозяйство имеет возраст около 10 тысяч лет, и весь этот период окультуриваемые растения и животные испытывали влияние искусственного отбора, который человек вел, исходя из «эгоистических» соображений.
Н.И. Вавилов считал, что большинство предков культурных растений обитало на горных осыпях, где вследствие постоянных естественных нарушений могли обитать только эксплеренты с низкой конкурентной способностью. Обработка почвы для возделывания таких эксплерентов моделировала нестабильные условия, подавлявшие растения с иными стратегиями. Искусственный отбор был направлен на повышение продукционного потенциала культурных растений, т. е. усиление свойства эксплерентности.
Поскольку эксплерентность образует трейдофф с виолентностью и патиентностью, то по мере повышения продукционного потенциала ослаблялась способность новых сортов противостоять действию неблагоприятных условий. Растениям потребовались удобрения, полив и защита от сорняков, вредителей и болезней. Затраты энергии на их возделывание увеличивались, что прямо или косвенно вело к разрушению среды (снижению плодородия почв, загрязнению, уменьшению биоразнообразия и т. д.). Эти тенденции наиболее ярко раскрылись в период Зеленой революции 60—70-х годов XX в.
В последние 10-20 лет направление селекции культурных растений изменилось, ее задачей стало повышение адаптивного потенциала сортов, т. е. их патиентности и виолентности (появился даже термин «дедоместикация», Kampf, 2000). Адаптивные сорта, приспособленные к определенным условиям среды, отличаются несколько меньшей урожайностью, но требуют несравненно меньших затрат на выращивание и потому менее опасны для окружающей среды.
Большие возможности биотехнологии, создающей генетически модифицированные сорта растений (ГМР), первоначально также были направлены на повышение продукционного потенциала. Однако в последние годы усилия биотехнологов направлены в первую очередь на повышение устойчивости ГМР к болезням, вызываемым грибами, и к насекомым-фитофагам. Большим успехом биотехнологов, к примеру, является картофель серии «новый лист», который устойчив против колорадского жука.
Такой же была история сельскохозяйственных животных. Долгое время их селекция была направлена на повышение продукционного потенциала (привеса, удоев, настрига шерсти и т. д.). В результате резко ослабла устойчивость этих животных к неблагоприятным воздействиям, для их содержания потребовались обильные корма, теплые помещения, целый набор препаратов для профилактики и лечения болезней. В настоящее время также наблюдается тенденция дедоместикации животных. В качестве селекционного материала используются животные «народных» пород, адаптированные к местным климатическим условиям.
Тема Синэкология - экология сообществ
Лекция девятая Определение биоценоза
Популяции разнообразных видов живых организмов, которые заселяют общие местожительства, обычно, вступают в определенные взаимоотношения, потому что вынужденные пользоваться общим жизненным пространством и пищевыми ресурсами. Поскольку стойкие взаимоотношения возможны только между популяциями видов, а стабильный характер таких отношений являет собой результат адаптаций, то после длительного общего существования формируются многовидовые группировки - биоценоз. Необходимо отметить, что набор видов в этих группировках не является случайным, а определяется возможностью поддержки круговорота веществ. Только на этой основе в принципе оказывается возможным стойкое существование любой формы жизни.
Биоценоз представляет собой совокупность растений, животных, грибов и микроорганизмов, которые совместно населяют участок земной поверхности и характеризуются определенными отношениями как между: собой, так и с совокупностью абиотических факторов.
Срок «биоценоз» предложен немецким зоологом К. Мебиусом в 1877 году.
K. Мебиус определил биоценоз - это объединение живых организмов, которое отвечает по своему составу, количеством видов и особей некоторым средним условиям среды, где организмы связаны взаимной зависимостью и сохраняются благодаря постоянному размножению в определенных местах. Если хотя бы одно из условий отклоняется хоть на некоторое время от средних условий, то изменяются и весь биоценоз.
Масштабы биоценотических группировок организмов чрезвычайно разнообразны - от группировок на стволе дерева или на болотной моховой кочке до биоценоза ковыльной степи.
Составными частями биоценоза является фитоценоз (совокупность растений), зооценоз (совокупность животных), микоценоз (совокупность грибов) и микробиоценоз (совокупность микроорганизмов). Синоним биоценоза - сообщество.
Участок поверхности планеты (суши или водоемы) с однотипными абиотическими условиями (рельефом, климатом, почвами, характером увлажнения и др.), которую занимает тот или другой биоценоз, называется биотопом (от грецьк. topos - место). Биотоп, с которым связанные организмы, которые здесь живут, условия их существования, испытывают изменения под воздействием биоценоза.
Однородность климатических условий биоценоза определяет климатоп, грунтовых - эдафотоп, увлажнение - гидротоп. При полевых исследованиях предел биоценозу устанавливают за фитоценозом, который легко распознать. Например, луговую группировку легко отличить от лесного, еловые леса – от сосновых, а верховое болото - от низинного.
Выделить пределы между двумя биоценозами несложно, если их абиотические и биотические факторы заметно отличаются (озеро и лука, лес и поле, болото).
Биоценоз - это совокупность растений, животных и микроорганизмов, которые заселяют один участок суши, водоема, характеризуются определенными отношениями между собой и приспособлениями к окружающей среде. Т.е. биоценоз – это стойкая система совместных существований автотрофных и гетеротрофных организмов (биоты) на определенном пространстве суши или акватории с конкурентными условиями (биотоп). Термин биоценоз предложил в 1877 Мебиус.
Биоценоз — совокупность популяций разных видов, обитающих на определенной территории. Растительный компонент биоценоза называется фитоценозом, животный — зооценозом, микробный — микробоценозом. Ведущим компонентом в биоценозе является фитоценоз. Он определяет каким будет зооценоз и микробоценоз.
Различают видовую, пространственную и экологическую структуры биоценоза.
В 1978 году, Троян предложил следующие критерии биоценоза:
биоценоз имеет характерный видовой состав (доминантные, субдоминантные виды)
имеет необходимый набор видов биоценоз – является системой, в границах которой реализуется оборотные материи и энергия, которой осуществляется между компонентов биоценоза и среды, по этому биоценоз должен иметь все элементы необходимые для оборота материи. Т.е. биоценоз должен содержать автотрофы (продуценты), гетеротрофы (редуценты), деструкторы.
характеризуется определенной продолжительностью во т биоценоз со своими видами составом является стойкой и долговечной, однако его обитатели имеют различную продолжительность жизни (микробы-минуты, мелкие беспозвоночные – дни, крупные – года, лесные деревья сотни лет). Биоценозы тропических лесов отличаются геологической историей, а на местах пожарищ развиваются новые биоценозы.
Имеет свою территорию и границы. Территория, функционирования отдельного биоценоза отличается однородностью и особенностью условий. Малые биоценозы могут существовать на нескольких метрах (источник – ручей). Крупные биоценозы занимают несколько сот км2. Выделить границы между двумя биоценозами не трудно, если их абиотический и биотический факторы заметно отличаются (луг, озеро) и в границах этих биоценозов при более тщательном рассмотрении можно выделить более тщательным рассмотрением сообщества. Сложность в изучении биоценозов является в том, что животные могут мигрировать в соседние биоценозы, потому нельзя утверждать, что определенному растительному сообществу обязательно соответствует определенное сообщество животных.
Пространственная структура биоценоза
Пространственная структура — распределение организмов разных видов в пространстве (по вертикали и по горизонтали). Пространственная структура образуется, прежде всего, растительной частью биоценоза. Различают ярусность (вертикальная структура биоценоза) и мозаичность (структура биоценоза по горизонтали).
Все биоценозы суши и воды характеризуются не равномерностью пространственного размещения. Составляют исключение агроэкосистемы или культурные биоценозы.
Пространственная неоднородность биоценоза обусловлена неоднородностью или мозаичностью места произрастания, которые колонизировали организмы в вертикальной структуре и формируют разные условия освещенности, обусловливающие эффективность образования первичной продукции биоценоза. Основным фактором, формирующим градиент вертикального расчленения среды, является количество фотосинтетической энергии, поступающей в разные экосистемы.
Для водных биоценозов характерно интенсивное поглощение солнечной энергии толщей воды. Выделяют фотичный слой, глубина которого превышает 400 м от зеркала воды, в нем выделяют слой воды, в котором солнечный свет присутствует в количествах превышающих компенсационную точку фотосинтеза растений и составляющий приблизительно 80 м.
Градиент проникновения солнечных лучей в биоценозах суши имеет вторичный характер, в отличие от водных биоценозов и зависит от развития растительности. Древесный ярус, в котором реализуется большинство фотосинтетических процессов, охватывает ярус крон. Он отдален от поверхности земли на десятки метров. Существующие под пологом верхнего яруса световые условия зависят от биологических особенностей деревьев, доминирующих в данном типе лесов. В тропических и хвойных лесах количество солнечного света изменяется не заметно, в лиственных в зависимости от смены листового покрова.
В вертикальной структуре биоценозов (на примере леса) выделяют четыре основных яруса:
ярус крон, высота (толщина) которого, зависит от видового состава деревьев.
кустарниковый ярус – охватывает как кустарники, так и деревья, которые в данных условиях могут расти в виде кустарника.
травяной ярус – включает в себя одно и многолетние травы, а так же мелкие кустарники.
Приземный ярус – включающий мхи и лишайники.
Таким образом, ярусность - это расчленение фитоценозов на структурные или функциональные горизонты, которые имеют разную степень замкнутости и играют неодинаковую роль в ассимиляции и аккумуляции веществ и энергии, в непосредственном и опосредованном влиянии на жизнедеятельность организмов сообщества.
Горизонтальная структура или мозаичность биоценоза связанна, прежде всего, с разнообразием подстилающей поверхности. Мозаичность биоценоза развивается вторично, т.к. в начале появляются виды аборигенов (пионеры). Одной из характерных особенностей горизонтальной структуры биоценозов является, переходные полосы от одного биоценоза к другому и которые называются экотоном (полоса берега - луг). В экотонах представлены виды обоих соседних биоценозов, хотя зачастую в этих полосах селятся виды, не встречающиеся ни в том, ни в другом биоценозе.
Важными характеристиками структуры биоценоза являются консорция, синузия и парцелла (рис. 1). Консорция — структурная единица биоценоза, объединяющая автотрофные и гетеротрофные организмы на основе пространственных (топических) и пищевых (трофических) связей вокруг центрального члена (ядра). Например, отдельно стоящее дерево или группа деревьев (растение-эдификатор) и связанные с ним организмы. Биоценоз — это система связанных между собой консорций. Синузия — структурная часть в вертикальном расчленении биоценоза, ограниченная в пространстве (или во времени). Пространственно синузия может совпадать с горизонтом, пологом, слоем, ярусом биогеоценоза. Например, в сосновом лесу можно выделить синузию сосны, синузию брусники, синузию зеленых мхов и т.д.
Рис. 1. Консорция, парцелла и синузия (Н.Ф. Реймерс, 1990)
Парцелла — структурная часть в горизонтальном расчленении биоценоза, отличающаяся от других частей составом и свойствами компонентов. Парцеллу выделяют (ограничивают) по ведущему элементу растительности. Например, участки широколиственных деревьев в хвойном лесу.
Отношение организмов в биоценозах. Живые организмы определенным образом связаны друг с другом. Различают следующие типы связей между видами (В.Н. Беклемешев, 1951): трофические, топические, форические, фабрические. Наиболее важными являются трофические и топические связи, так как именно они удерживают организмы разных видов друг возле друга, объединяя их в сообщества.
Трофические связи возникают между видами, когда один вид питается другим: живыми особями, мертвыми остатками, продуктами жизнедеятельности. Трофическая связь может быть прямой и косвенной. Прямая связь проявляется при питании львов живыми антилопами, гиен трупами зебр, жуков-навозников пометом крупных копытных и т.д. Косвенная связь возникает при конкуренции разных видов за один пищевой ресурс.
Трофические связи наблюдаются, когда один вид питается другим (либо мертвыми остатками, либо продуктами их жизнедеятельности). Прямая трофическая связь осуществляется с видами, предоставляющими им пищу.
Косвенная трофическая связь возникает конкуренции двух видов из-за объектов питания. Т.к. деятельность одних отражается на снабжении кормом других. Воздействие одного вида на поедаемость другого или доступность для него пищи, является косвенной трофической связью.
Топические связи проявляются в изменение одним видом условий обитания другого вида. Например, под хвойным лесом, как правило, отсутствует травянистый покров. К ним относятся:
создание одним видом среды обитания для другого – паразитирующего.
формирование субстрата, на котором поселяются или избегают поселяться другие виды.
влияние на движение воды, воздуха, изменение температуры, освещаемости в различных положительных или отрицательных точках взаимоотношений.
В биоценозах трофические и топические связи имеют наибольшее значение и составляют основу его существования.
Форические связи возникают, когда один вид участвует в распространении другого вида. Перенос животными семян, спор, пыльцы растений называется зоохория, а мелких особей — форезия.
В роли транспортеров выступают животные перенос животными семян, спор, пыльцы – называется зоохорий. Перенос животными более мелких животных называется форезией. Форезия распространена среди животных, в основном в членистоногих и свойственная тем видам, для которых перенос из одного биотипа в другой жизненно необходим для сохранения и процветания.
Фабрические связи заключаются в том, что один вид использует для своих сооружений (фабрикаций) продукты выделения, мертвые остатки или даже живых особей другого вида. Например, птицы при постройке гнезд используют ветки деревьев, траву, пух и перья других птиц.
Каждый конкретный вид из-за сложности взаимоотношений между видами может преуспевать не везде, где складываются подходящие для него условия физической среды. Различают физиологический и синэкологический оптимумы распространения вида. Физиологический оптимум – это благоприятное для вида сочетание всех абиотических факторов, при которых возможно наиболее быстрые темпы роста и размножения. Синэкологический оптимум – это биотическое окружение, при котором вид испытывает наименьшее давление со стороны врагов и конкурентов, что позволяет ему успешно размножиться. Физиологический и синэкологический оптимумы не всегда совпадают.
Межвидовые связи, формирующие биоценоз, обуславливают закономерные соотношения в нем видов, их экологических особенностей, численности, распределения в пространстве, или, можно сказать, позволяют создать определенною структуру биоценоза.
Экологическая структура — соотношение организмов разных экологических групп. Биоценозы со сходной экологической структурой могут иметь разный видовой состав. Это связано с тем, что одни и те же экологические ниши могут быть заняты сходными по экологии, но далеко не родственными видами. Такие виды называются замещающими или викарирующими.
Любой биоценоз, естественный или искусственный, имеет определенный набор популяций различной численности. Однако в определении природы и функций сообщества не все организмы имеют одинаковое значение. Из большого количества видов, входящих в какое-либо сообщество, часто лишь небольшое их число оказывает на него определяющее воздействие, обусловленное их численностью, размерами, продукцией и другими параметрами. В сообществе роль каждого вида не зависит от таксономических отношений, так как организмы, оказывающие регулирующее воздействие, часто принадлежат к разным таксономическим группам.
Лекция десять Видовая структура биоценоза
Экологическая структура — соотношение организмов разных экологических групп. Биоценозы со сходной экологической структурой могут иметь разный видовой состав. Это связано с тем, что одни и те же экологические ниши могут быть заняты сходными по экологии, но далеко не родственными видами. Такие виды называются замещающими или викарирующими.
Любой биоценоз, естественный или искусственный, имеет определенный набор популяций различной численности. Однако в определении природы и функций сообщества не все организмы имеют одинаковое значение. Из большого количества видов, входящих в какое-либо сообщество, часто лишь небольшое их число оказывает на него определяющее воздействие, обусловленное их численностью, размерами, продукцией и другими параметрами. В сообществе роль каждого вида не зависит от таксономических отношений, так как организмы, оказывающие регулирующее воздействие, часто принадлежат к разным таксономическим группам.
Видовая структура — число видов, образующих данный биоценоз, и соотношение их численности или массы. То есть видовая структура биоценоза определяется видовым разнообразием и количественным соотношением числа видов или их массы между собой.
Видовое разнообразие — число видов в данном сообществе. Встречаются бедные и богатые видами биоценозы. Видовое разнообразие зависит от возраста сообщества (молодые сообщества беднее, чем зрелые) и от благоприятности основных экологических факторов — температуры, влажности, пищевых ресурсов (биоценозы высоких широт, пустынь и высокогорий бедны видами). Различают α- и β-разнообразие. α-разнообразие — видовое разнообразие в данном местообитании, β-разнообразие — сумма всех видов всех местообитаний в данном районе.
Высоким видовым разнообразием отличаются экотоны — переходные зоны между сообществами, а увеличение здесь видового разнообразия называется краевым эффектом.
В сообществе различают следующие виды: доминантные, преобладающие по численности, и «второстепенные», малочисленные и редкие. Среди доминантов особо выделяют эдификаторов (строителей) — это виды, определяющие микросреду (микроклимат) всего сообщества. Как правило, это растения.
О значимости отдельного вида в видовой структуре биоценоза судят по нескольким показателям: обилие вида, частота встречаемости и степень доминирования. Обилие вида — число или масса особей данного вида на единицу площади или объема занимаемого им пространства. Частота встречаемости — процентное отношение числа проб или учетных площадок, где встречается вид, к общему числу проб или учетных площадок. Характеризует равномерность или неравномерность распределения вида в биоценозе. Степень доминирования — отношение числа особей данного вида к общему числу всех особей рассматриваемой группировки.
Основные сообщества состоят из продуцентов, консументов и редуцентов. В пределах названных групп отдельные виды могут принимать главенствующее участие в регуляции энергетического обмена и оказывать более заметное влияние на среду обитания других видов. Такие виды называются экологическими доминантами. Степень доминирования в сообществе одного или нескольких видов выражается показателем доминирования, который отражает значение каждого вида для сообщества.
Упрощенную классификацию биотического сообщества можно показать на следующем примере: на пастбище могут сосуществовать мятлик, клевер, дуб, мясной скот, куры, индейки, овцы, лошади. Простой перечень сообщества не дает полного представления о нем, необходима количественная оценка видов. Например, мятлик луговой — 20 га, клевер ползучий — 0,8 га, дуб — 2 дерева, мясной скот — 2 особи, молочный скот — 48 особей, козы — 6 особей, овцы — 1 особь, индейки — 1 особь, лошади — 1 особь. Теперь очевидно, что среди продуцентов доминирует мятлик, а среди консументов — молочный скот. Естественно, более полную картину дала бы информация о сезонных вариациях в использовании пастбища, о годовой продуктивности и т.д. Доминантами можно назвать те виды, которые на своем трофическом уровне наиболее продуктивны, преобладают над другими видами в сообществе по массе или численности.
Классификация элементов сообщества. Любой биоценоз как естественный, так и искусственный может определять набор популяций различной численности. Из большого количества видов, входящих в сообщество, лишь небольшое их число оказывает на него определяющее воздействие, обусловленное их численностью размерами и др. параметрами в пределах сообщества отдельные виды занимают главенствующее участие в регуляции энергетического обмена, и оказывает более заметное влияние на среду обитания других видов, такие виды называют экологическими доминантами. Степень доминирующего в биоценозе одного или несколько видов выражает показатели доминирования, который отражает значение каждого вида для сообщества. Рассмотрим упрощенную классификацию биотического сообщества на примере пастбища. На пастбище могут сосуществовать мятник, клевер, дуб, мясной скот, куры, индейки, лошади. Простой перечень сообщества не дает полного представления о нем, необходима количественная оценка вида. Т.е. метлик луговой 20 га, клевер ползучий 0,8 га, дуб 2 дерева, мясной скот 2 особи, молочный скот 48, козы 6 особей, овцы 1 особь. Индейки 1 особь, 1 лошадь, курей 3 особи. Отсюда видно, что среди продуцентов доминируют мятлик, среди консументов молочный скот. Доминантами можно назвать те виды, которые на своем уровне наиболее продуктивны, преобладают над другими видами в сообщество по массе и численности.
Показатель доминирования (с), отражающий степень доминирования каждого вида в сообществе, определяется как сумма квадратов отношений показателей значимости видов (n,) к общей оценке значимости (N):
C=∑(ni/N)2
Доминирование некоторых видов в сообществе не означает, что многочисленные виды не играют никакой роли. Они определяют видовое разнообразие, что не менее важно для структуры сообщества.
Для оценки видового разнообразия можно использовать следующие критерии: видовое богатство (d); показатель выравненности (е), показатель общего разнообразия (Н), или показатель Шеннона:
Видовое богатство (d):
d1 = (S -1) / log N; d2 = S/100 особей,
где S — число видов, N — общая оценка значимости вида.
Показатель выравненности (е):
e = H/logS.
Показатель общего разнообразия (Н), или показатель Шеннона:
Н=∑(п; / N) log(ni/ N), или Н = ∑PilogPi,
где ni,— оценка «значительности» для каждого вида;
Р — вероятность «значительности» для каждого вида Рi< n/N.
Создать классификацию сообществ популяции по какому-то одному показателю довольно сложно, так как видовое разнообразие, соотношение между видами и структура биотического сообщества, как правило, специфичны для определенных условий. Биотические сообщества удобно именовать и классифицировать в соответствии:
с основными структурными показателями, такими как доминирующие виды, жизненные формы или виды-индикаторы;
с условиями обитания сообществ;
с функциональными особенностями, такими как тип метаболизма сообщества.
В сообществе не многие из составляющих его видов, имеют значительную численность, большую биомассу, продуктивность. Если основное количество энергии потребляют доминирующие виды, то видовое разнообразие этой группы определяют основные более редкие виды.
Показатель видового разнообразия – это соотношение между численностью видов, и каким либо показателем значимости численности, биомасса, продуктивность в состав сообществ чаще всего входят несколько видов с высоко численностью и несколько видов с малой численностью.
Количество соотношений, между которыми варьируют в широких пределах, несмотря на то, что общий поток энергии, поступающий в экосистему, влияет на видовое разнообразие. Эти величины не связанны линейной зависимостью. Высоко продуктивное сообщество могут характеризоваться очень высокими показателями видовых разнообразий.
Стабильность экосистемы тесно связанна видовым разнообразием или продуктивностью. На видовое разнообразие большое влияние оказывает функции связи между трофичными уровнями и таксономическими единицами (масштабы выпаса или хищничества существенно влияют на разнообразие травостоя на пастбищах; умеренное хищничество снижает плотность, доминантов, предоставляя возможность конкурирующим видам лучше использовать пространство и ресурсы).
Виды, живущие за счет доминантов, получили название предоменантов. Например, в сосновом лесу таковыми являются кормящиеся на сосне насекомые, мышевидные грызуны. В биоценозах существуют виды, которые называются эдификаторами, создающие своей жизнедеятельностью среду для всего сообщества и без которых в связи с этим существование большинства других видов невозможно. Удаление вида эдификатора из биоценоза влечет за собой изменение физической среды и в первую очередь микроклимата биотопа т.о. все виды, слагающие биоценоз в определенной степени связанны с доминирующими видами и эдификаторами.
Консорции. Внутри биоценоза формируются группировки комплекса популяций, которые зависят от растения-эрификатора или других элементов биоценоза и создают своеобразные структурные единицы биоценоза – консорции (Раменский 1952 год). Консорция - это совокупность популяций организмов, жизнедеятельность которых в пределах одного биоценоза, трофически или топически связана с центральным видом - автотрофным растением. В роли центрального вида выступают эдификатор - основной вид, который определяет особенности биоценоза. Популяции остальных видов консорции образуют ее ядро за счет, которого существуют виды, разрушающие органическое вещество, создаваемое автотрофами. Популярность автотрофа растения березы на базе, которого формируется консорция и называется детерминантом, а виды, объединенные вокруг него, консортами.
Группы консортов того или иного порядка, объединенные вокруг детерминанта называются концентрами. Каждая консорция охватывает большое количество видов, среди которых есть виды, входящие в состав только одной консорции и виды, которые являются членами 20 и более консорций, тем самым способствуют объединению организмов биоценоза в единый комплекс. Состав консорций – результат длительного процесса подбора видов, способных существовать в условиях местообитания детерминанта. Каждая консорция представляет собой особую структурную единицу биоценоза и экосистемы.
Рис. 2. Схема строения консорции:
/, //, III — концентры, / — центральный вид — детерминант консорции (автотроф), 2 — консорты первого концентра, 3 —«, консорты второго и третьего концентров.
Обычно консорцию изображают в виде концентрической фигуры (рис.), помещая в ее центре автотрофное растение — детерминант консорции. Вокруг него располагаются группы организмов, непосредственно взаимодействующих с детерминантом,— это так называемые консорты. К ним относятся животные-фитофаги, микроорганизмы, грибы, совокупность которых образует первый концентр.
Второй концентр составляют животные-зоофаги, поедающие животных-фитофагов, а также виды идущие в свите консортов первого концентра. По мере повышения трофического уровня автономность консорции все более и более снижается, так как хищник-зоофаг может поедать фитофагов, питающихся разными растениями. Кроме того, существует много всеядных животных.
В настоящее время вполне правомерным считается вычленение как автотрофных (ядром консорции является зеленое растение), так и гетеротрофных (ядро — особь гетеротрофного вида) консорции. Существование последних обеспечивается за счет энергии центральной живой особи, независимо от того, ассимилирует она эту энергию путем фотосинтеза или за счет поедания живой или мертвой органической массы. Таким образом, любой организм не только автотрофного, но и гетеротрофного способа питания является источником энергии для других, связанных с ним консортивными связями организмов. Поэтому консорцию рассматривают как явление общебиологическое. В связи с этим оспаривается мнение ботаников о том, что ядром консорции следует считать только автотрофный организм.
С животными трофически связано не только множество симбиотических и паразитических микроорганизмов и насекомых. С ними форически связано много высших и низших растений (семена, споры, микроскопические грибы, водоросли). Ярко выраженным ядром консорции являются морские животные, ведущие неподвижный образ жизни
Экологическая ниша (Гриннел1917 Элтон 1933). Любая популяция (вид) занимает определенное местообитание и определенную экологическую нишу.
Местообитание — это территория или акватория, занимаемая популяцией (видом), с комплексом присущих ей экологических факторов. Местообитание вида является компонентом его экологической ниши. Применительно к наземным животным местообитание вида называется стация, местообитание сообщества — биотоп.
Экологическая ниша — совокупность всех факторов среды, в пределах которых возможно существование вида в природе. То есть экологическая ниша — это место вида в природе, включающее не только его положение в пространстве и отношение к абиотическим факторам, но и его функциональную роль в сообществе (прежде всего трофический статус). Местообитание — это как бы «адрес» организма, а экологическая ниша — это его «профессия».
Для характеристики экологической ниши обычно используют два важных показателя: ширина ниши и степень перекрывания ее с соседними. Экологические ниши разных видов могут быть разной ширины и перекрываться в различной степени.
Разделение экологических ниш между видами происходит за счет приуроченности разных видов к разным местообитаниям, разной пищи и разному времени использования одного и того же местообитания. Принцип конкурентного исключения (принцип Гаузе) гласит: «Два вида не могут сосуществовать в одной и той же местности, если их экологические потребности идентичны. Такие виды обязательно должны быть разобщены в пространстве или во времени».
Группы видов в сообществе, обладающие сходными функциями и нишами одинакового размера, т.е. роль которых в сообществе одинакова или сравнима, называются гильдиями. Например, лианы тропического леса представлены многими видами растений. Между видами внутри гильдии наблюдается особенно острая конкуренция.
Виды, занимающие одинаковые ниши в разных географических областях, называются экологическими эквивалентами. Например, крупные кенгуру Австралии, бизоны Северной Америки, зебры и антилопы Африки и т.д. являются экологическими эквивалентами. В настоящее время они значительно замещены коровами и овцами.
Выделяют 4 типа расположения экологических ниш 2-х видов одного биотопа:
Рис. 1 Випадки можливих взаємних піш, проілюстровані за допомогою поняття щільності пристосованості (зліва) та моделей теорії множин (справа):
а — ніша всередині ніші. Ніші виду 2 (S2) розташовані всередині ніші (St). Можливі два наслідки конкуренції: 1) якщо вид 2 має перевагу (переривчаста лінія), то він буде співіснувати при неповному використанні спільних ресірсів з видом і; 2) якщо переваги має вид 1 (суцільні лінії), то він буде використовувати весь градієнт ресірсів, а вид 2 буде витіснений;
6 — перекривання ніш однакової ширини. Конкуренція однакова в обох напрямках;
в — перекривання ніш неоднакової ширини. Конкуренція неоднакова у двох напрямах, оскільки частина простору ніш, яка входить до області перекривання, у виду 2 більша, ніж у виду 1;
г — прилягання ніш. Пряма конкуренція неможлива, але подібне може бути наслідком її уникання;
д — повний розділ ніш. Конкуренція неможлива і її важко навіть передбачити. Чому ж тут не спостерігається кормова конкуренція? А тому, що кожний із видів в процесі еволюції пристосувався до своєї екологічної ніші.
Екологічна ніша — фізичний простір з властивими йому екологічними умовами, що визначають існування будь-якого організму, місце виду в природі, що включає не лише становище його в просторі, а й функціональну роль у біоценозі та ставлення до абіотичних факторів середовища існування. Екологічна ніша характеризує ступінь біологічної спеціалізації даного виду.
Различают фундаментальную (потенциальную) нишу, которую организм мог бы занимать в отсутствие конкурентов, хищников и других врагов и в которой физические условия оптимальны; и реализованную – фактический диапазон условий существования организма, который или меньше фундаментальной ниши или равен ей.
Фундаментальную нишу называют иногда предконкурентной, а реализованную – постконкурентной.
Два вида с совершенно одинаковыми потребностями не могут существовать вместе: один из них через какое то время обязательно будет вытеснен. Это положение получило статус закона известного под названием принципа конкурентного вытеснения или принцип Гаузе. Исключение – бакланы.
Выделяют четыре направления конкуренции влияющие на:
географическое расширение видов (интродукция)
на размещение вида в различных биотопах одной и той же местности.
влияние на морфологию и продуктивность.
влияние на эволюцию видов биоценоз конкурентные отношения
Тема - Экология экосистем
Лекция одиннадцать Определение экосистемы
Предметом экологии является изучение условий закономерностей существования, формирования и функционирования экосистем. Объектом экологии является экосистема.
Термин экосистема был предложен А. Тенсли в 1935 году, который считал, что экосистема - это единая открытая функциональная система, образованная организмами и средой их обитания с которой они активно взаимодействуют.
Экосистема – это любая совокупность живых существ и среда их обитания, объединенная в единое функциональное целое, возникающая на основе взаимозависимости и причинно-следственных связей, существующих между отдельными экологическими компонентами.
Совокупность специфического физико-химического окружения (биотопа) с сообществом живых организмов (биоценозом) и образует экосистему.
Тенсли предложил следующее соотношение
Биотоп + биоценоз = экосистема.
Экосистема — система живых организмов и окружающих их неорганических тел, связанных между собой потоком энергии и круговоротом веществ (рис. 1). Биотоп — определенная территория со свойственными ей абиотическими факторами среды обитания (климат, почва). Биоценоз – совокупность живых организмов на определенной территории.
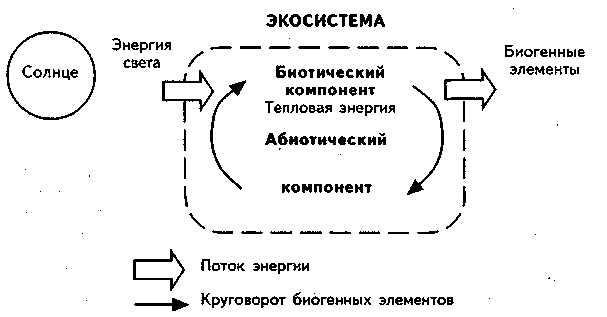
Рис. 1. Функциональная схема экосистемы
Рис. -2 – Структура биогеоценоза
Биогеоценоз — совокупность биоценоза и биотопа (рис. 1). Термин «экосистема» был предложен английским ученым А. Тенсли (1935), а термин «биогеоценоз» — российским ученым В.Н. Сукачевым (1942).
«Экосистема» и «биогеоценоз» — понятия близкие, но не синонимы. Биогеоценоз — это экосистема в границах фитоценоза. Экосистема — понятие более общее. Каждый биогеоценоз — это экосистема, но не каждая экосистема — биогеоценоз.
В нашей стране и за рубежом идея о взаимосвязи и единстве всех явлений и предметов на земной поверхности, т. е. о природных комплексах, возникла в какой-то мере почти одновременно, с той лишь разницей, что в СССР она развивалась как учение о биогеоценозе, а в других странах — как учение об экосистеме.
Биогеоценоз и экосистема — понятия сходные, но не тождественные. В обоих случаях — это взаимодействующие совокупности живых организмов и среды, но экосистема — понятие безразмерное. Муравейник, аквариум, болото, биосфера в целом, кабина космического корабля и т. п.— все это экосистемы. В отечественной литературе принято характеризовать биогеоценоз как экосистему, границы которой определены фитоценозом, т. е. суженный к пределам фитоценоза участок биогеоценотического покрова Земли. Иными словами, биогеоценоз — это частный случай, определенный ранг экосистемы. Биогеоценоз — сложный природный комплекс живых существ, находящихся в зависимости от неорганической среды и взаимодействующих с ней материально-энергетическими связями. По своей сущности это динамическая, уравновешенная, взаимосвязанная и стойкая во времени система, которая является результатом длительной и глубокой адаптации составных компонентов и в которой может осуществляться круговорот веществ. Биогеоценоз — не простая совокупность живых организмов и их среды обитания, а особая, согласованная форма существования организмов и окружающей среды, диалектическое единство всех экологических компонентов, объединенных в единое функциональное целое на основе взаимозависимости и причинно-следственных связей. Биогеоценозы земного шара образуют биогеоценотический покров, который изучает биогеоценология. Основателем этой науки был выдающийся советский ученый В. Н. Сукачев. Совокупность всех биогеоценозов (экосистем) нашей планеты создает гигантскую глобальную экосистему, называемую биосферой.
Биогеоценозы могут формироваться на любом участке земной поверхности, сухопутном и водном. Биогеоценозы бывают степными, болотными, луговыми и т. д. Единая экосистема нашей планеты называется биосферой. Биосфера — экосистема высшего порядка.
Выделяют микро, мезо и мегаэкосистемы экосистемы. При этом меньшие по размеру входят в качества подсистем в более крупные функции, образуя иерархию в которой каждый уровень организации взаимосвязаны с отсутствием четких граней между ними т.о. иерархия экосистемы в биосфере та их взаимоподчиняемость в порядке укрупнения и усложнения. Отсюда вытекает по мере объединения компонентов в более крупные у новых единиц, возникают качественно новые свойства, которые отсутствуют на предыдущем уровне. Примером мегаэкосистемы (глобальной) является биосфера.
Так как все экосистемы, включая биосферу, являются открытыми, то для своего функционирования, они должны получать и отдавать энергию, т.е. реальная функционирующая экосистема должна иметь вход и выход переработанной энергии. Энергия солнечного света поступает в экосистему, где фотоавтотрофными организмами превращается в химическую энергию, используемую для синтеза органических веществ из неорганических. В экосистеме поток энергии направлен в одну сторону: часть поступающей энергии солнца, преобразуется растениями и переходит на качественно более высокую ступень, превращаясь в органическое вещество, которое представляет собой более концентрированную форму энергии. Большая часть солнечной энергии приходит через экосистемы и покидает ее. В отличие от энергии, вода и элементы питания, необходимые для жизни, могут использоваться многократно (после отмирания органические вещества превращаются в неорганические). В состав экосистемы входит два компонента: сообщество живых организмом или биоценоз (биотический компонент) и физико-химическая среда или биотоп (абиотический компонент).

Р ис.3
- Функциональная схема экосистемы.
ис.3
- Функциональная схема экосистемы.
Таким образом, экосистему можно представить как единое целое, в котором биогенные вещества из абиотического компонента включаются в биотический и обратно, т.е. происходит постоянный круговорот веществ с участием биотического и абиотического компонентов.
Любая природная система может развиваться только за счет использования материально-энергетических и информационных возможностей окружающей ее среды (закон развития природной системы за счет окружающей среды).
Структурой экосистемы принято называть совокупность ее системообразующих связей. Учитывая характер взаимодействий между биотическим и абиотическим компонентами, можно выделить несколько аспектов единой внутренней структуры экосистемы:
- энергетическую (совокупность энергетических потоков в экосистеме);
- вещественную (совокупность потоков вещества);
- информационную (совокупность внутриэкосистемных информационных потоков);
- пространственную (характеризующую пространственное распределение потоков энергии, вещества и информации внутри экосистемы);
- динамическую (определяющую изменение внутриэкосистемных потоков во времени).
Среди различных аспектов структуры экосистемы на первый план выступает именно энергетическая структура, так как поток энергии в значительной степени определяет и характер потоков вещества и особенности информационных взаимодействий, и пространственно-динамические закономерности.
Структурную организацию экосистемы можно рассмотреть с трофической и биологической точек зрения.
С точки зрения трофической структуры экосистему можно разделить на два яруса — автотрофный и гетеротрофный (по Ю. Одуму, 1986).
1. Верхний автотрофный ярус, или «зеленый пояс», включающий растения или их части, содержащие хлорофилл, где преобладают фиксация энергии света, использование простых неорганических соединений и накопление сложных органических соединений.
2. Нижний гетеротрофный ярус, или «коричневый пояс» почв и осадков, разлагающихся веществ, корней и т.д., в котором преобладают использование, трансформация и разложение сложных соединений.
С биологической точки зрения в составе экосистемы удобно выделить следующие компоненты (по Ю. Одуму, 1986):
неорганические вещества;
органические соединения;
воздушную, водную и субстратную среду;
продуцентов;
макроконсументов;
микроконсументов.
Неорганические вещества (С02, Н20, N2, 02, минеральные соли и др.), включающиеся в круговороты.
Органические вещества (белки, углеводы, липиды, гумусовые вещества и др.), связывающие биотическую и абиотическую части.
Воздушная, водная и субстратная среда, включающая абиотические факторы.
Продуценты — автотрофные организмы, способные производить органические вещества из неорганических, используя фотосинтез или хемосинтез (растения и автотрофные бактерии).
5. Консументы (макроконсументы, фаготрофы) — гетеротрофные организмы, потребляющие органическое вещество продуцентов или других консументов (животные, гетеротрофные растения, некоторые микроорганизмы). Консументы бывают первого порядка (фитофаги, сапрофаги), второго порядка (зоофаги, некрофаги) и т.д.
6.Редуцееты (микроконсументы, деструкторы, сапротрофы, осмотрофы) — гетеротрофные организмы, питающиеся органическими остатками и разлагающие их до минеральных веществ (сапротрофные бактерии и грибы).
Следует учитывать, что и продуценты, и консументы частично выполняют функции редуцентов, выделяя в окружающую среду минеральные вещества — продукты их метаболизма.
Таким образом, как правило, в любой экосистеме можно выделить три функциональные группы организмов: продуцентов, консументов и редуцентов. В экосистемах, образованных только микроорганизмами, консументы отсутствуют. В каждую группу входит множество популяций, населяющих экосистему. В экосистеме пищевые и энергетические связи идут в направлении: продуценты —» консументы —> редуценты.
Энергетическая структура экосистемы
Согласно закону максимизации энергии (Г. и Э. Одумов, 1972) выживает та экосистема, которая обеспечивает максимальное поступление и наиболее эффективное использование энергии.
Поток энергии в экосистемах осуществляется через сложный механизм, включающий процессы ассимиляции, накопление, транспортирование и высвобождение энергии. Важнейший этап в этом механизме - ассимиляция энергии, которая представляет собой превращение кинетической энергии фотонов в биологически доступную потенциальную энергию химических связей в органических соединениях. По способности к ассимиляции энергии организмы делятся на два типа:
- автотрофы (продуценты) - организмы, самостоятельно синтезирующие органические вещества, поглощая минеральные вещества и энергию, поступающую из вне;
- гетеротрофы (консументы и редуценты) – организмы, получающие необходимые органические вещества и энергию с пищей, представляющую собой живую или мертвую биомассу.
К автотрофам относятся зеленые растения и некоторые виды бактерий.
Гетеротрофами являются все животные, грибы, подавляющее большинство бактерий, некоторые водоросли и бесхлорофилльные высшие растения.
Транспорт энергии в экосистемах осуществляется через пищевые цепи. Пищевая цепь – это перенос энергии в форме пищи от ее источника (автотрофа) через ряд организмов, происходящий путем поедания одних организмов другими. Обычно различают два типа пищевых цепей:
- пастбищная цепь, начинающаяся живыми растениями (продуцентами), живой тканью которых питаются растительноядные животные (фитофаги);
- детритная цепь, начинающаяся мертвым органическим веществом (детритом), потребляемым мертвоядными организмами (детритофагами).
Пастбищная;
фитофаги
Зоофаг 2
Растен.





детритофаги
зоофаг 1
Детритная;
Рис. 4 - Типы пищевых цепей
В сложных природных сообществах пищевые цепи тесно переплетаются друг с другом, образуя пищевые сети. Организмы, отстоящие от начала пищевой цепи на одинаковое число звеньев, образуют трофический уровень. На первом трофическом уровне находятся только продуценты. Ко второму трофическому уровню принадлежат фитофаги и детритофаги (первичные консументы), к третьему – зоофаги (вторичные консументы) и т.д.
Границы между трофическими уровнями нерезкие. Многие животные потребляют энергию более чем с одного трофического уровня, т.е. используют любую пищу, которая подходит им по размеру и другим признакам. Передача энергии по пищевой цепи подчиняется второму закону термодинамики, согласно которому процессы, связанные с превращениями энергии, могут происходить самопроизвольно только при условии, что часть этой энергии рассеивается в форме тепла. В связи с этим, в каждом звене пищевой цепи теряется значительная доля (80-95%) энергии, заключенной в доступной пище. На каждый последующий трофический уровень поступает в среднем не более 10% доступной энергии (правило 10%).
Зеленые растения ассимилируют от 1 до 5% получаемой солнечной энергии, а так как при дыхании растений разрушается до 80% синтезируемых углеводов, эффективность фотосинтеза составляет не более 1,5%. Фитофаги для образования своей биомассы используют в среднем всего 1% энергии, содержащейся в поедаемых ими растениях. На других трофических уровнях эффективность усвоения доходит до 10% (иногда даже больше).
Из «правила десяти процентов» вытекают важные следствия.
Чем ближе организм находится к началу пищевой цепи, тем большим количеством доступной энергии он располагает.
Пищевые цепи, включающие более 4-5 трофических уровней, встречаются крайне редко.
Закономерности транспорта энергии в пищевых цепях удобно изображать графически с помощью диаграмм которые называются «экологические пирамиды». Они представляют собой соотношение численности организмов, их биомасс, эквивалентных этим биомассам энергий или продукции организмов на различных трофических уровнях пищевой цепи.
Существует три основных типа пирамид
-пирамида чисел - показывает численность отдельных организмов;
-пирамида биомассы - характеризует общий сухой вес, калорийность или другую меру общей численности живого вещества;
-пирамида энергии отвечает величине потока энергии или продуктивности на последовательных трофических уровнях.
Пирамиды чисел и биомассы могут быть обратными, т.е. основа может быть меньшей, чем один или несколько верхних уровней. Так бывает когда средние размеры продуцентов меньше чем размеры консументов. Пирамида энергии всегда сужается к верху при условии, что будут учтены все источники питания в системе.
Продуктивность экосистемы тесно связанна с потоком энергии, проходящим через нее. В каждой экосистеме только часть поступающей энергии накопляется в виде органических соединений. Скорость ассимиляции энергии называется продукцией, а величина продукции, отношение к единице площади экосистемы называется продуктивностью.
Общая скорость, с которой энергия усваивается продуцентами и накапливается в форме органического вещества, называется валовой первичной продукцией. Чистая первичная продукция – это скорость накопления органического вещества в растительных тканях за вычетом расхода энергии на жизнеобеспечение организма. Вторичная продукция создается гетеротрофами.
Величина продукции, отнесенная к единице площади экосистемы, носит название продуктивности. Так как в экосистемах только растения ассимилируют солнечную энергию, трансформируя ее в органическое вещество, а остальные только ее потребляют, поэтому продукция экосистемы в целом зависит от соотношения расхода энергии растениями на фотосинтез и дыхание. В неблагоприятных условиях, при стрессовых нагрузках расходы на дыхание организмов увеличивается, и продукция экосистемы снижается.
Пищевые цепи и сети. Питаясь друг другом, живые организмы образуют цепи питания. Цепь питания — последовательность организмов, по которой передается энергия, заключенная в пище, от ее первоначального источника. Каждое звено цепи называется трофическим уровнем. Первый трофический уровень — продуценты (автотрофные организмы, преимущественно зеленые растения). Второй трофический уровень - консументы первого порядка (растительноядные животные). Третий трофический уровень — консументы второго порядка (первичные хищники, питающиеся растительноядными животными). Четвертый трофический уровень — консументы третьего порядка (вторичные хищники, питающиеся плотоядными животными). В пищевой цепи редко бывает больше 4—5 трофических уровней. Последний трофический уровень — редуценты (сапротрофные бактерии и грибы). Они осуществляют минерализацию — превращение органических остатков в неорганические вещества.
Различают два типа пищевых цепей (рис. 10). Цепи выедания (или пастбищные) — пищевые цепи, начинающиеся с живых фотосинтезирующих организмов. Например, фитопланктон —» зоопланктон—> рыбы микрофаги —» рыбы макрофаги —> птицы ихтиофаги. Цепи разложения (или детритные) — пищевые цепи, начинающиеся с отмерших остатков растений, трупов и экскрементов животных. Например, детрит —» детритофаги —> хищники микрофаги —» хищники макрофаги. Таким образом, поток энергии, проходящий через экосистему, разбивается как бы на два основных направления. Энергия к консументам поступает через живые ткани растений или через запасы мертвого органического вещества. Цепи выедания преобладают в водных экосистемах, цепи разложения — в экосистемах суши.
.
Рис. 5. Y-образная модель потока энергии, показывающая связь между пастбищной и детритной пищевыми цепями (Ю. Одум, 1986)
В сообществах пищевые цепи сложным образом переплетаются и образуют пищевые сети. В состав пищи каждого вида входит обычно не один, а несколько видов, каждый из которых в свою очередь может служить пищей нескольким видам. С одной стороны, каждый трофический уровень представлен многими популяциями разных видов, с другой стороны, многие популяции принадлежат сразу к нескольким трофическим уровням. В результате благодаря сложности пищевых связей выпадение какого-то одного вида часто не нарушает равновесия в экосистеме
Любой экосистеме свойственен круговорот веществ и прохождение через нее потока энергии.
В экосистеме органические вещества синтезируются автотрофами из неорганических веществ. Затем они потребляются гетеротрофами. Выделенные в процессе жизнедеятельности или после гибели организмов (как автотрофов, так и гетеротрофов) органические вещества подвергаются минерализации, т.е. превращению в неорганические вещества. Эти неорганические вещества могут быть вновь использованы автотрофами для синтеза органических веществ. Так осуществляется биологический круговорот веществ.
В то же время, энергия не может циркулировать в пределах экосистемы. Поток энергии (передача энергии), заключенной в пище, в экосистеме осуществляется однонаправленно от автотрофов к гетеротрофам.
Рис. 6. Схема потока энергии в пищевой цепи, показывающая три трофических уровня
(по Ю. Одум, 1986)
I— поступление энергии на трофический уровень; L — солнечная энергия, попадающая на растения; LA — энергия, поглощаемая растениями (около 0,5 L); Lu — неиспользуемая часть энергии солнечного света; А — ассимиляция, т.е. энергия, ассимилированная (запасенная) в органическом веществе; R — траты на дыхание; PG — валовая первичная продукция (около 0,05 LA); PN — чистая первичная продукция; Я - вторичная продукция (консументов); Nu — неиспользуемая консументами энергия (накапливаемая или экспортируемая); NA — неассимилируемая консументами энергия (выделенная с экскрементами). Цифры внизу - порядок величины потерь энергии при каждом переносе, начиная с поступления солнечного излучения в количестве 3000 ккал/м2 в сутки.
На рис. 11 представлена упрощенная схема потоков энергии на трех трофических уровнях пищевой цепи. Трофические уровни изображены как резервуары, размер которых соответствует энергии, заключенной в их биомассе (5), размер соединяющих их каналов — величине потоков энергии. На входе в систему поступает поток солнечной энергии (/). На первом трофическом уровне зелеными растениями поглощается около 50 % солнечной энергии (LA). Только часть ее в процессе фотосинтеза преобразуется в энергию химических связей органических веществ. Это валовая первичная продукция (Р^) - Большая часть поглощенной растениями, но не усвоенной энергии, рассеивается в окружающую среду в виде тепловой энергии. Часть образованных органических веществ окисляется, а высвобождающаяся энергия расходуется на поддержание всех метаболических процессов. Это так называемые траты на дыхание (R). Эта энергия, в конечном счете, также рассеивается в виде тепла. Оставшаяся часть новообразованных органических веществ составляет прирост биомассы растений (DB) и называется чистой первичной продукцией (PN) - В чистую первичную продукцию превращается только 1 % поглощенной растением энергии. Таким образом, Pw = PQ — R.
До второго трофического уровня доходит только часть чистой первичной продукции. Некоторая ее часть (Nu) не используется консументами первого порядка. Она может накапливаться или экспортироваться за пределы системы. Та часть, которую ассимилировали (потребили) консументы (А), частично тратится на дыхание (R), частично выделяется с экскрементами (NA), а остальное накапливается в виде вторичной продукции (Р).
Вторичная продукция на каждом последующем трофическом уровне консументов (Р2, Р3 и т.д.) составляет около 10 % предыдущей (хотя на уровне хищников может быть выше — около 20 %). В результате, чем длиннее пищевая цепь, тем меньше остается к ее концу накопленной в органическом веществе энергии. Поэтому число трофических уровней никогда не бывает слишком большим.
Экологическая пирамида и ее типы. В экосистеме при передаче энергии с одного трофического уровня на другой большая часть энергии рассеивается в виде тепла (в соответствии со вторым законом термодинамики), и только около 10 % от первоначального количества передается по пищевой цепи.
В результате, пищевые цепи можно представить в виде экологических пирамид. Различают три основных типа экологических пирамид (рис. 12).
Пирамида чисел (пирамида Элтона) отражает уменьшение численности организмов от продуцентов к консументам.
Пирамида биомасс показывает изменение биомасс на каждом следующем трофическом уровне: для наземных экосистем пирамида биомасс сужается кверху, для экосистемы океана — имеет перевернутый характер, что связано с быстрым потреблением фитопланктона консументами.
Пирамида энергии (продукции) имеет универсальный характер и отражает уменьшение количества энергии, содержащейся в продукции, создаваемой на каждом следующем трофическом уровне.
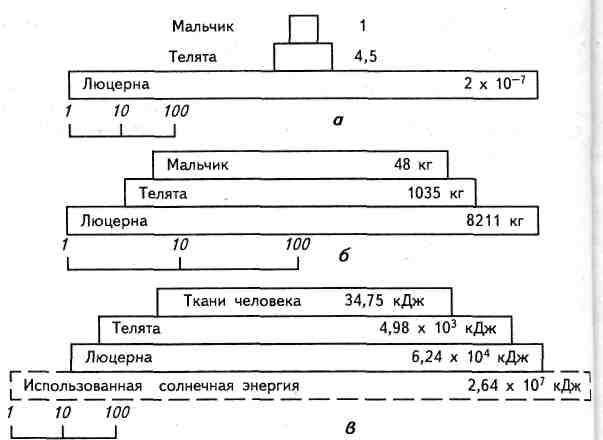
Рис. 7 Пирамиды чисел (а), биомасс (б) и энергии (в), представляющие упрощенную экосистему: люцерна — телята — мальчик 12 лет (по Ю. Одуму, 1959)
Пирамида чисел (а) показывает, что если бы мальчик питался в течение одного года только телятиной, то для этого ему потребовалось бы 4,5 теленка, а для пропитания телят необходимо засеять поле в 4 га люцерной, что составит 2 х Ю-7 растений. В пирамиде биомасс (6) число особей заменено их биомассой. В пирамиде энергии (в) учтена солнечная энергия. Люцерна использует 0,24 % солнечной энергии. Для накопления продукции телятами в течение года используется 8 % энергии, аккумулированной люцерной. На развитие и рост ребенка в течение года используется 0,7 % энергии, аккумулированной телятами. В результате чуть более одной миллионной доли солнечной энергии, падающей на поле в 4 га, используется для пропитания ребенка в течение одного года.
Биологическая продукция (продуктивность) экосистемы — называется прирост биомассы в экосистеме, созданной за единицу времени. В данном контексте более правильно пользоваться понятием «продуктивность». Однако вместо этого чаще используют понятие «продукция», все равно учитывая при этом и фактор времени. Различают первичную и вторичную продукцию сообщества.
Первичная продукция — биомасса, созданная за единицу времени продуцентами. Она делится на валовую и чистую. Валовая первичная продукция (общая ассимиляция) — это общая биомасса, созданная растениями в ходе фотосинтеза. Часть ее расходуется на поддержание жизнедеятельности растений — траты на дыхание (40— 70 %). Оставшаяся часть составляет чистую первичную продукцию (чистая ассимиляция), которая в дальнейшем используется консументами и редуцентами, или накапливается в экосистеме.
Вторичная продукция — биомасса, созданная за единицу времени консументами. Она различна для каждого следующего трофического уровня.
Масса организмов определенной группы (продуцентов, консументов, редуцентов) или сообщества в целом называется биомассой. Самой высокой биомассой и продуктивностью обладают тропические дождевые леса, самой низкой — пустыни и тундры (табл. 8).
Если в экосистеме скорость прироста растений (образования первичной продукции) выше темпов переработки ее консументами и редуцентами, то это ведет к увеличению биомассы продуцентов. Если при этом присутствует недостаточная утилизация продуктов опада в цепях разложения, то происходит накопление мертвого органического вещества. Это ведет к эвторфовыванию болот,
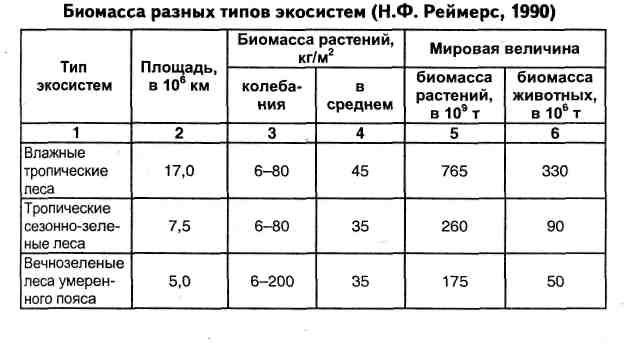
образованию мощной лесной подстилки и т.п. В стабильных экосистемах биомасса остается постоянной, так как практически вся продукция расходуется в цепях питания.
Энергетическая классификация экосистем
В зависимости от источника энергии и степени энергетических субсидий Ю. Одум (1986) разделил существующие экосистемы на 4 типа.
Природные экосистемы, движимые Солнцем и несубсидируемые (например, открытые океаны, глубокие озера, высокогорные леса). Они получают мало энергии и имеют низкую продуктивность, но при этом занимают основные площади биосферы.
2 Природные экосистемы, движимые Солнцем и субсидируемые другими естественными источниками (например, эстуарии в приливных морях, некоторые дождевые леса, речные экосистемы). Помимо солнечного света они получают дополнительную энергию в виде дождя, ветра, органических веществ, минеральных элементов и т.д.
Экосистемы, движимые Солнцем и субсидируемые человеком (например, агроэкосистемы, аквакультуры). Дополнительная энергия поставляется в них человеком в виде горючего, органических и минеральных удобрений, пестицидов, стимуляторов роста и т.п. Эти экосистемы производят продукты питания и другие материалы.
Индустриально-городские экосистемы, движимые топливом (например, города, пригороды, промышленные комплексы). Основным источником энергии служит не Солнце, а топливо. Эти экосистемы зависят от экосистем первых трех типов, паразитируют на них, получая продукты питания и топливо.
Информационная структура экосистем. Все природные экосистемы обладают двумя принципиальными свойствами – динамизмом и устойчивостью. Динамизм – это способность систем направленно изменятся, а устойчивость – свойство сохранять установившееся внутреннее динамическое равновесие. Под воздействием различных причин в экосистемах проходят закономерные изменения, проявляющиеся в вытеснении одних видов другими. Эти изменения происходят под воздействием как внешних, так и внутренних факторов. К внешним факторам, прежде всего, относятся климатические и геологические факторы, а также вмешательство человека. Внутренние изменения экосистем обусловлены активным воздействием живых организмов на среду обитания. Последовательность сукцессионых изменений направлены в сторону образования стабилизированной экосистемы, в которой максимально сбалансированные потоки вещества и энергии. Сбалансированность потоков вещества и энергии в зрелых экосистемах приводит к замыканию круговоротов основных биогенных элементов, к возрастанию видового разнообразия сообщества. Пищевые цепи пастбищного типа превращаются в детритные разветвления. Увеличиваются размеры особей, скорость обмена веществ между организмами и средой обитания снижается. Возрастает эффективность использования энергии и вещества системой. При загрязнении или иной форме отрицательного воздействия на экосистему вектор изменения признаков сукцессии становится противоположным: наблюдается обеднение видового состава, уменьшение общей биомассы, размыкание круговоротов и т.д. Зрелая экосистема, достигая гомеостатического состояния, проявляет свойство устойчивости.
Устойчивость экосистемы можно охарактеризовать, как способность сохранять ею свою структуру и характер функционирования при изменяющихся условиях среды. Устойчивость экосистемы выражается в поддержании фактически постоянным состава атмосферы и океанической воды, стабильности климата, сохранении постоянным соотношение численности особей и т.д.
Помимо потоков энергии и круговорота вещества, экосистемы характеризуются развитыми информационными сетями, и относятся к типу кибернетических или управляемых систем. Основными задачами управления в природных системах являются две – стабилизация, принимающая форму поддержания устойчивого состояния системы и оптимизация, как адаптация, приспособление системы к изменяющимся факторам окружающей среды.
Главным средством реализации управляющих функций является механизм обратной связи, который подразумевает влияние процессов, явлений и событий, протекающей на выходе системы, на входной сигнал.
В качестве примера действия обратной связи рассмотрим взаимозависимость следующих явлений.
1. Многие виды бобовых растений образуют особые вздутия – пузырьки, в которых поселяются клубеньковые бактерии, фиксирующие азот и снабжают растения нитратами, что в свою очередь повышает продуктивность растений и увеличивает количество углеводов, потребляемых клубеньковыми бактериями. А это в свою очередь способствует усилению фиксации азота.
2. При повышении вулканической деятельности в атмосферу поступает дополнительное количество углекислого газа, что приводит к увеличению его содержания в атмосфере. Это приводит к увеличению фотосинтетической активности зеленых растений (повышается потребление растениями углекислого газа) и, следовательно, способствует снижению его содержания в атмосфере.
В первом явлении обратная связь усиливает действие входного сигнала, (продуктивность растений увеличивается одновременно с ростом азотофиксирующей способности бактерий). Такую обратную связь принято называть положительной. Во втором примере взаимосвязь явлений «гасит» внешнее воздействие (дополнительные количества СО2 поступающего в атмосферу, поглощается зелеными растениями в процессе фотосинтеза). Это отрицательная обратная связь. На языке кибернетики обратную связь можно определить как возвращение части сигнала с выхода системы на ее вход.

Рис.8 - Графическая модель контура обратной связи.
В зависимости от способа реализации кибернетического механизма системы могут быть с внешним и с внутренним управлением (саморегулирующиеся системы).
Внутреннее управление осуществляется за счет взаимодействия подсистем данной системы и характерно для всех природных систем.
Системы, контролируемые внутренней обратной связью обладают определенной независимостью поведения, проявляющиеся в их способности к саморазвитию и устойчивости. Наиболее устойчивы крупные экосистемы и самая стабильная из них – биосфера, а наиболее неустойчивы – молодые экосистемы. Это объясняется тем, что в больших экосистемах создается саморегулирующийся гомеостаз за счет взаимодействия круговоротов веществ и потока энергии.
Среди форм отношений между элементами различных систем в живой природе и в человеческом обществе, одно из главных мест занимают парные взаимодействия, которые обобщенно могут быть обозначены как «ресурс-потребитель ресурса». Взаимодействия в каждой из таких пар можно представить в виде контура обратных связей. В качестве примера действия обратной связи рассмотрим взаимозависимость следующих явлений. Корни многих бобовых растений образуют симбиоз с клубеньковыми бактериями, которые снабжают растения нитратами. Это повышает продуктивность растения, увеличивают количество углеводов, потребляемых клубеньковыми бактериями, а это в свою очередь способствует усилению фиксации азота. При повышении вулканической деятельности в атмосферу Земли поступают дополнительные количества СО2, что может привести к росту его содержания. Однако это усиливает фотосинтетическую активность растений, что в свою очередь вызывает повышенное потребление растениями СО2, а это способствует снижению его содержания в атмосфере.
В кибернетике обратную связь определяют как возвращение части сигнала с выхода системы на ее вход. В первом случае обратная связь усиливает действие входного сигнала, (продуктивность растения увеличивается вместе с ростом азотфиксирующей способности бактерий). Такую обратную связь называют положительной. Такие связи не только не способствуют регуляции, а наоборот, генерируют дестабилизацию систем, приводя их либо к угнетению и гибели, либо к ускоряющемуся росту, за которым, как правило, также следует срыв и разрушение системы. Во втором случае взаимосвязь явлений «гасит», нейтрализует внешнее воздействие. Это отрицательная обратная связь.
На принципе отрицательной обратной связи построены все механизмы регуляции физиологических функций в любом организме и поддержание постоянства внутренней среды, т.е. гомеостаза любой авторегуляторной системы. Все экологические системы включают контуры отрицательных обратных связей. Системы, контролируемые внутренней обратной связью, обладают определенной независимостью поведения, проявляющиеся в их способности к саморазвитию и устойчивости.
Выделяют два типа простейших контуров обратной связи: усилитель сигнала и регулятор сигнала. Регулятор сигнала - контур с отрицательной обратной связью. Этот механизм обеспечивает стабилизацию состояния системы, возмущаемой внешним воздействием. Примером действия отрицательной обратной связи в биоценозах может служить взаимодействие в системе «хищник-жертва». Рост численности популяции жертвы, обусловленный благоприятными внешними условиями вызывает рост численности популяции хищников, что в свою очередь, снижает численность жертвы, (контур работает по принципу «чем больше, тем меньше»).
В целом такой контур имеет отрицательный знак. Это значит, что система авторегуляторна и может сама себя поддержать в рамках определенного уровня соотношения численностей. Усилитель сигнала - контур с положительной обратной связью. Этот механизм обеспечивает экстенсивное развитие системы, т.к. приводит к еще большему отклонению состояния, чем – то, которое вызвало бы внешнее воздействие (входной сигнал) при отсутствии обратной связи.
Действие положительной обратной связи можно рассматривать как процесс автостимуляции (самоускорения) системы, когда она на всякий внешний толчок реагирует возрастающим удалением от исходного состояния. Остановить ее в этом движении может только истощение ресурсов, необходимых для экстенсивного развития.
Особенностью механизма «усилитель сигнала» является то, что при внешнем воздействии достаточной интенсивности вектор движения системы меняется на обратный. В сложных системах всегда сочетаются конторы обоих знаков. В природных системах скорость распространения возмущающего воздействия характеризуется широким спектром значений. Запаздывание сигнала в контуре отрицательной обратной связи вызывает колебания - периодические изменения характеристик взаимосвязанных процессов.
Лекция 12 Основные круговороты веществ в биосфере
Вещественная структура экосистемы – это совокупность потоков вещества. В экосистеме происходит постоянный круговорот элементов питания с участием биотического и абиотического компонентов. Движущей силой круговоротов служит солнечная энергия, которую используют непосредственно фотосинтезирующие организмы и затем передают ее другим представителям биотического компонента. Перемещение элементов и неорганических соединений, необходимых для жизни с помощью биогеохимических циклов, называют круговоротом элементов питания.
Биотическую и абиотическую части биосферы связывают непрерывные потоки вещества, которые делятся на два типа: большой (геологический) и малый (биотический) круговороты. Большой круговорот наиболее ярко проявляется в круговороте воды и циркуляции атмосферы, а малый, развивающийся на основе большого и состоящий в непрерывном, циклическом, но неравномерном во времени и пространстве, и сопровождающийся более или менее значительными потерями закономерного перераспределения вещества энергии и информации в пределах экосистем различного уровня организации.
Оба круговорота взаимно связаны и представляют как бы единый процесс. Подсчитано, что весь кислород, содержащийся в атмосфере оборачивается через организмы (связывается при дыхании и высвобождается при фотосинтезе) за 2000 лет, СО2 совершает круговорот в обратном направлении за 300 лет, а все воды на Земле разлагаются и воссоздаются путем фотосинтеза и дыхания за 2000000 лет.
Взаимодействие абиотических факторов и живых организмов экосистемы сопровождается непрерывным круговоротом вещества между биотопом и биоценозом в виде чередующихся то органических, то минеральных соединений. Существование подобных круговоротов создает возможность для саморегуляции (гомеостаза), системы, что придает экосистеме устойчивость: удивительное постоянство процентного содержания различных элементов. Здесь действует принцип функционирования экосистемы: получение ресурсов и избавление от отходов в рамках круговорота всех элементов. Основу биотического круговорота составляет взаимодействие организмов трех экологических типов: продуцентов, консументов и редуцентов, представленных в биоценозе любой экосистемы.
В настоящее время сильное влияние на них оказывает хозяйственная деятельность человека, что ведет к нарушению биосферы и может иметь тяжелые последствия для будущих поколений землян. Рассмотрим круговороты основных биогенов - углерода, кислорода, азота, воды.
Круговорот углерода. Это один из самых важнейших биосферных круговоротов, поскольку углерод составляет основу органических веществ. В круговороте особенно велика роль диоксида углерода (рис. 1).
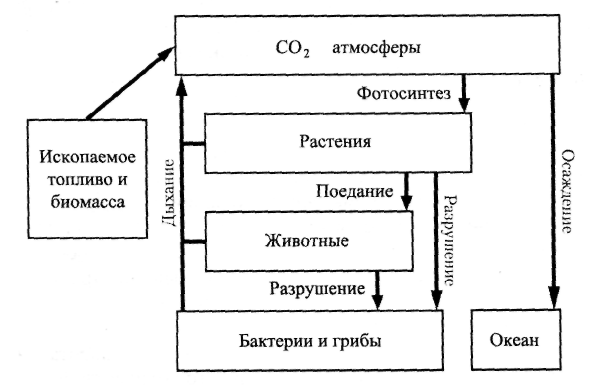
Рис. 1. Круговорот углерода в биосфере
Запасы «живого» углерода в составе организмов суши и океана составляют, по разным данным, 550—750 Гт (1 Гт = 1 млрд т), причем 99,5% его сосредоточено на суше, остальное - в океане. Кроме того, в океане содержится до 700 Гт углерода в составе растворенного органического вещества.
Запасы неорганического углерода значительно больше. Над каждым квадратным метром суши и океана находится 1 кг углерода атмосферы и под каждым квадратным метром океана на глубине 4 км - 100 кг углерода в форме карбонатов и бикарбонатов. Еще больше запасов углерода в осадочных породах — в известняках содержатся карбонаты, в сланцах - керогены и т.д.
Примерно 1/3 «живого» углерода (около 200 Гт) циркулирует, т. е. ежегодно усваивается организмами в процессе фото синтеза и возвращается обратно в атмосферу, причем вклад океана и суши в этот процесс примерно сходный. Несмотря на то, что биомасса океана много меньше биомассы суши, его биологическая продукция создается множеством поколений краткоживущих водорослей (соотношение биомассы и биологической продукции в океане примерно такое же, как в пресноводной экосистеме.
До 50% (по некоторым данным — до 90%) углерода в форме диоксида возвращают в атмосферу микроорганизмы-редуценты почвы. В этот процесс равный вклад вносят бактерии и грибы. Возврат диоксида углерода при дыхании всех прочих организмов, таким образом, меньше, чем при деятельности редуцентов.
Некоторые бактерии кроме диоксида углерода образуют метан. Выделение метана из почвы возрастает при переувлажнении, когда создаются анаэробные условия, благоприятные для деятельности метанообразующих бактерий. По этой причине резко увеличивается выделение метана лесной почвой, если древостой вырублен и вследствие уменьшения транспирации происходит ее заболачивание. Много метана выделяют рисовые поля и домашний скот.
В настоящее время отмечается нарушение круговорота углерода в связи со сжиганием значительного количества ископаемых углеродистых энергоносителей, а также дегумификацией пахотных почв и осушением болот. В целом содержание диоксида углерода в атмосфере ежегодно увеличивается на 0,6%. Еще быстрее возрастает содержание метана - на 1-2%. Эти газы являются главными виновниками усиления парникового эффекта, который на 50% зависит от диоксида углерода и на 33% - от метана.
Последствия усиления парникового эффекта для биосферы неясны, наиболее вероятный прогноз - потепление климата. Однако поскольку «машинами» климата являются морские течения, то вследствие их изменения при таянии ледников в ряде районов возможно существенное похолодание (в том числе в Европе в результате изменения течения Гольфстрим). Под влиянием изменения концентрации диоксида углерода значительно учащаются крупные стихийные бедствия (наводнения, засухи и т.д.)
Приведенные данные характеризуют биогенный круговорот углерода. В круговороте участвуют и геохимические процессы, при которых происходит обмен атмосферного углерода и углерода, содержащегося в горных породах. Однако данных о скорости этих процессов нет. Полагают лишь, что их интенсивность менялась в истории планеты и парниковый эффект, который наблюдается сегодня, многократно проявлялся в прошлом при усилении геохимических процессов с выделением диоксида углерода и при ослаблении процессов, которые «оттягивали» его из атмосферы.
Для того чтобы вернуть круговороту углерода равновесие, необходимо увеличить площадь лесов и сократить выброс газов при сжигании углеродистых энергоносителей.
Круговорот воды. Вода испаряется не только с поверхности водоемов и почв, но и живых организмов, ткани которых на 70 % состоят из воды (рис. 2). Большое количество воды (около 1/3 всей воды осадков) испаряется растениями, особенно деревьями: на созидание 1 кг органического вещества в разных районах они расходуют от 200 до 700 л воды.
Различные фракции воды гидросферы участвуют в круговороте по-разному и с разной скоростью. Так, полное обновление воды в составе ледников происходит за 8 тыс. лет, подземных вод - за 5 тыс. лет, океана - за 3 тыс. лет, почвы - за 1 год. Пары атмосферы и речные воды полностью обновляются за 10-12 суток.
До развития цивилизации круговорот воды был равновесным, однако в последние десятилетия вмешательство человека нарушает этот цикл. В частности, уменьшается испарение воды лесами ввиду сокращения их площади и, напротив, увеличивается испарение с поверхности почвы при орошении сельскохозяйственных культур. Испарение воды с поверхности океана уменьшается вследствие появления на ее значительной части пленки нефти.
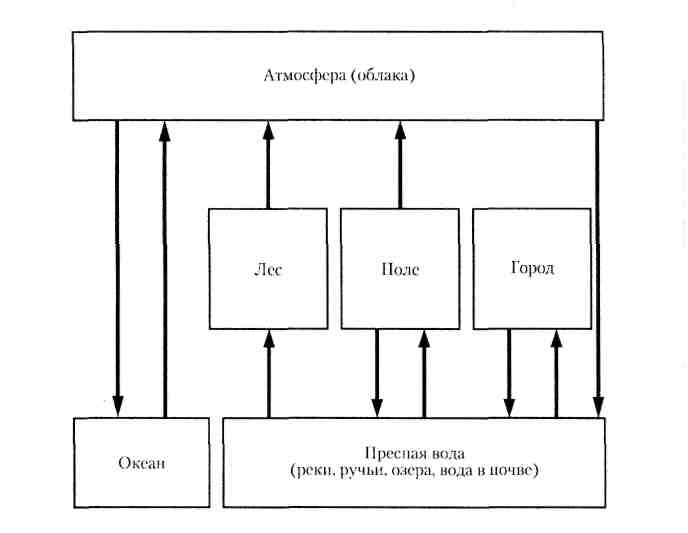
Рис. 2. Круговорот воды в биосфере
Влияет на круговорот воды потепление климата, вызываемое парниковым эффектом. При усилении этих тенденций могут произойти существенные изменения круговорота, опасные для биосферы.
Важную роль в годовом водном балансе биосферы играет океан. Испарение с его поверхности примерно в 2 раза больше, чем с поверхности суши.
Круговорот азота. Циркуляция азота в биосфере протекает по следующей схеме (рис. 3):
перевод инертного азота атмосферы в доступные для растений формы (биологическая азотфиксация, образование аммиака при грозовых разрядах, производство азотных удобрений на заводах);
усвоение азота растениями;
переход части азота из растений в ткани животных;
накопление азота в детрите;
- разложение детрита микроорганизмами-редуцентами вплоть до восстановления молекулярного азота, который возвращается в атмосферу.
В морских экосистемах азотфиксаторами являются циано-бактерии, связывающие азот в аммиак, который усваивается фитопланктоном.
В настоящее время вследствие уменьшения доли естественных экосистем биологическая азотфиксация стала меньше промышленной фиксации азота (соответственно 90-130 и 140 млн т в год), причем к 2020 г. ожидается увеличение промышленной азотфиксации на 60%. До половины азота, вносимого на поля, вымывается в грунтовые воды, озера, реки и вызывает эвтрофикацию водоемов.
Значительное количество азота в форме оксидов азота поступает в атмосферу, а затем в почву и водоемы в результате ее загрязнения промышленностью и транспортом (кислотные дожди). Этот азот был изъят из атмосферы экосистемами геологического прошлого и длительное время находился «на депоненте» в угле, газе, нефти, при сжигании которых он возвращается в круговорот. Например, в США азота с атмосферными осадками в год выпадает 20—50 кг/га, а в отдельных районах эмиссия достигает 115 кг/га.
Экологически безопасной считается величина эмиссии азота 10—30 кг/га в год. При более высоких нагрузках происходят значительные изменения в экосистемах: почвы подкисляются, происходит выщелачивание питательных элементов в глубокие горизонты, возможно усыхание древостоев и массовое развитие заносных видов-нитрофилов. Кроме того, высокое содержание азота в растениях, выросших на загрязненных азотом почвах, повышает их поедаемость, что может привести к выпадению из растительных сообществ даже доминантных видов. Так, в некоторых пустошах Западной Европы после того, как в вереске повысилось содержание азота, массово размножился вересковый жук (его количество достигало 2000 экземпляров на 1 м2). Жук практически полностью выел этот кустарник из сообществ. Те же изменения в составе загрязняемых промышленным азотом сообществ отмечены и в Калифорнии.
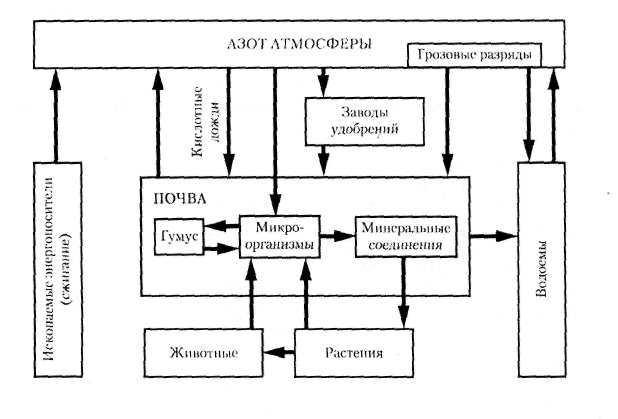
Рис. 3. Круговорот азота в биосфере
Однако не всегда кислотные дожди оказывают пагубное влияние на экосистемы. Экосистемы степной зоны, где почвы имеют слабощелочную реакцию, от выпадения кислотных дождей не только не страдают, но даже увеличивают свою продуктивность за счет дополнительного азота.
Восстановление естественного круговорота азота возможно за счет уменьшения производства азотных удобрений, резкого сокращения промышленных выбросов оксидов азота в атмосферу и расширения площади посевов бобовых, которые симбиотически связаны с бактериями-азотфиксаторами.
Круговорот кислорода. Кислород атмосферы имеет биогенное происхождение и его циркуляция в биосфере осуществляется путем пополнения запасов в атмосфере в результате фотосинтеза растений и поглощения при дыхании организмов и сжигании топлива в хозяйстве человека (рис. 4). Кроме того, некоторое количество кислорода образуется в верхних слоях атмосферы при диссоциации воды и разрушении озона под действием ультрафиолетового излучения; часть кислорода расходуется на окислительные процессы в земной коре, при вулканических извержениях и др.
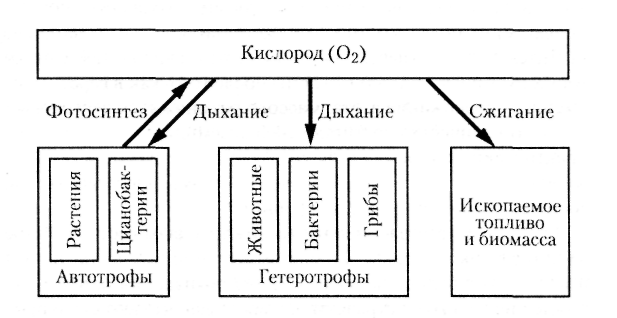
Рис.4. Круговорот кислорода в биосфере
Этот круговорот очень сложный, так как кислород вступает в разнообразные реакции и входит в состав очень большого числа органических и неорганических соединений, и замедленный. Для полного обновления всего кислорода атмосферы требуется около 2 тысяч лет (для сравнения: ежегодно обновляется около 1/3 диоксида углерода атмосферы).
В настоящее время поддерживается равновесный круговорот кислорода, хотя в крупных густонаселенных городах с большим количеством транспорта и промышленных предприятий возникают локальные нарушения.
Однако отмечается ухудшение состояния озонового слоя и образование «озоновых дыр» (областей с пониженным содержанием озона) над полюсами Земли, что представляет экологическую опасность. Временные «дыры» возникают также над обширными районами вне полюсов (в том числе и над континентальными районами России). Причиной этих явлений является попадание в озоновый слой хлора и оксидов азота, которые образуются в почве из минеральных удобрений при их разрушении микроорганизмами, а также содержатся в выхлопных газах автомобилей. Эти вещества разрушают озон с более высокой скоростью, чем он может образовываться из кислорода под влиянием ультрафиолетовых лучей. Сохранение озонового слоя — одна из глобальных задач мирового сообщества. Для прекращения разрушения озонового слоя и его восстановления необходимо отказаться от использования хлорсодержащих веществ - хлорфторуглеродов (фреонов), используемых в аэрозольных упаковках и холодильных установках. Необходимо также уменьшить количество выхлопных газов двигателей внутреннего сгорания и дозы азотных минеральных удобрений в сельском хозяйстве.
Так как озон является фотооксидантом, образующимся из оксида азота и углеводородов под влиянием ультрафиолетовых лучей, то возможно увеличение содержания азона в приземном слое атмосферы. В этом случае он становится опасным загрязнителем, вызывающим раздражение дыхательных путей человека. Однако отрицательно сказывается на здоровье человека и чрезмерно низкое содержание озона в атмосфере.
Круговорот фосфора. О круговороте фосфора за обозримое время можно говорить лишь условно. Будучи гораздо тяжелее углерода, кислорода и азота, фосфор почти не образует летучих соединений — он стекает с суши в океан, а возвращается в основном при подъеме суши в ходе геологических преобразований. По этой причине круговорот фосфора называют «открытым» (рис. 6). Фосфор содержится в горных породах, откуда выщелачивается в почву и усваивается растениями, а затем по пищевым цепям переходит к животным. После разложения мертвых тел растений и животных не весь фосфор вовлекается в круговорот, часть его вымывается из почвы в водоемы (реки, озера, моря). Там фосфор оседает на дно и почти не возвращается на сушу, лишь небольшое количество его возвращается с выловленной человеком рыбой или с экскрементами птиц, питающихся рыбой. Скопления экскрементов морских птиц служили в недалеком прошлом источником ценнейшего органического удобрения - гуано, но в настоящее время ресурсы гуано практически исчерпаны.
Рис. 6. Круговорот фосфора в биосфере
Отток фосфора с суши в океан усиливается вследствие возрастания поверхностного стока воды при уничтожении лесов, распашке почв и внесении фосфорных удобрений. Поскольку запасы фосфора на суше ограничены, а его возврат из океана проблематичен (хотя в настоящее время активно исследуются возможности его добычи со дна океана), в будущем в земледелии возможен острый дефицит фосфора, что вызовет снижение урожаев (в первую очередь зерна). Поэтому необходима экономия ресурсов фосфора.
Лекция 13 - Продуктивность и динамика экосистем
Экологическая пирамида и ее типы. В экосистеме при передаче энергии с одного трофического уровня на другой большая часть энергии рассеивается в виде тепла (в соответствии со вторым законом термодинамики), и только около 10 % от первоначального количества передается по пищевой цепи. В основе существования пищевых цепей лежат, помимо второго закона термодинамики, также размерные закономерности поедаемых организмов. Как правило, размеры пищи, которую могут добывать определенные животные, имеют довольно четкие верхний и нижний пределы. Соотношения размеров в пищевой цепи с хищниками обычно противоположны таковым в цепях с паразитами: во втором случае организмы-консументы в каждом последующем звене не увеличиваются, а мельчают. Так, на корнях сельскохозяйственных культур паразитируют нематоды, паразитами которых, в свою очередь, являются микроскопические организмы. В трофический оборот экосистемы включается в среднем 1 % поступающей в нее солнечной энергии. На каждом следующем трофическом уровне сохраняется лишь около 10 % энергии, усвоенной организмами предыдущего трофического уровня, а 80—90 % ее рассеивается в экосистеме в виде тепла. Поэтому чем выше трофический уровень консумента, тем меньше суммарная биомасса его особей.
Рис. 1. Пирамида чисел для луга, поросшего злаками:
Пр— продуценты (растения), БТ — беспозвоночные травоядные, БП — беспозвоночные плотоядные, П — позвоночные. Цифры — число особей.
На практике, однако, ограничиваются схематическим изображением пищевой сети и включают лишь ее главные звенья: первичные продуценты, т. е. зеленые растения; первичные консументы, т. е. травоядные животные; вторичные консументы, или плотоядные животные; деструкторы. Эти звенья изображают условно в виде прямоугольников, длина или площадь которых соответствует, например, численным значениям указанных звеньев. Располагая эти прямоугольники в соподчиненной последовательности, получают экологическую пирамиду, отражающую соотношение численности особей в пищевой цепи (рис. 1). Такой тип экологической пирамиды назван пирамидой чисел, основной принцип построения которой впервые сформулировал американский эколог Ч. Элтон. По этому же принципу составляют пирамиду биомасс. В этом случае размер прямоугольников пропорционален массе живого вещества каждого звена — продуцентов, первичных и вторичных консументов и деструкторов. Термин «пирамида биомасс» возник в связи с тем, что в огромном большинстве экосистем масса первичных консументов, живущих за счет продуцентов, намного меньше массы продуцентов, а масса вторичных консументов в свою очередь значительно меньше массы первичных консументов. Пирамида биомасс, таким образом, складывается из трех прямоугольников, уменьшающихся в размерах снизу вверх. Что касается биомассы деструкторов, то ее помещают иногда отдельно. Основное значение пирамиды состоит в том, что она показывает количество живого вещества на каждом трофическом уровне.
Скорость создания органического вещества не определяет его суммарные запасы, т. е. общую биомассу всех организмов каждого трофического уровня. Наличная масса продуцентов или консументов в конкретных экосистемах зависит от того, как соотносятся между собой темпы накопления органического вещества на определенном трофическом уровне и передачи его на вышестоящий, т. е. насколько сильно выедание образовавшихся запасов. Немаловажную роль при этом играет скорость оборота генераций основных продуцентов и консументов.
Отношение первичной продукции растений к фитомассе определяет те масштабы выедания растительности, которые возможны в сообществе без подрыва его продуктивности. Относительная доля потребляемой животными первичной продукции в травянистых сообществах.
В противоположность пирамидам чисел и биомасс, отражающим статику системы (количество организмов в данный момент), пирамида энергии отражает картину скоростей прохождения массы пищи через пищевую цепь. На форму этой пирамиды не влияют изменения размеров и интенсивности метаболизма особей, и если учтены все источники энергии, то пирамида всегда будет иметь типичный вид, как это диктуется вторым законом термодинамики.
Экологические пирамиды позволяют иллюстрировать количественные отношения в отдельных, представляющих особый интерес случаях, например в звеньях жертва — хищник или хозяин — паразит.
Пирамиды чисел и биомассы могут быть обратными, т.е. основа может быть меньшей, чем один или несколько верхних уровней. Так бывает когда средние размеры продуцентов меньше чем размеры консументов. Пирамида энергии всегда сужается к верху при условии, что будут учтены все источники питания в системе.
В результате, пищевые цепи можно представить в виде экологических пирамид. Различают три основных типа экологических пирамид (рис. 3).
Пирамида чисел (пирамида Элтона) отражает уменьшение численности организмов от продуцентов к консументам.
Пирамида биомасс показывает изменение биомасс на каждом следующем трофическом уровне: для наземных экосистем пирамида биомасс сужается кверху, для экосистемы океана — имеет перевернутый характер, что связано с быстрым потреблением фитопланктона консументами.
Пирамида энергии (продукции) имеет универсальный характер и отражает уменьшение количества энергии, содержащейся в продукции, создаваемой на каждом следующем трофическом уровне.
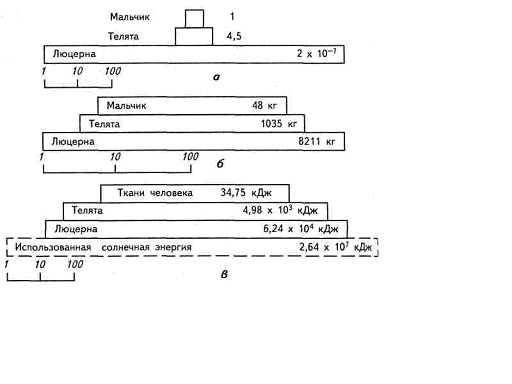
Рис. 2. Пирамиды чисел (а), биомасс (б) и энергии (в), представляющие упрощенную экосистему: люцерна — телята — мальчик 12 лет (по Ю. Одуму, 1959)
Пирамида чисел (о) показывает, что если бы мальчик питался в течение одного года только телятиной, то для этого ему потребовалось бы 4,5 теленка, а для пропитания телят необходимо засеять поле в 4 га люцерной, что составит 2 х Ю-7 растений. В пирамиде биомасс (6) число особей заменено их биомассой. В пирамиде энергии (в) учтена солнечная энергия. Люцерна использует 0,24 % солнечной энергии. Для накопления продукции телятами в течение года используется 8 % энергии, аккумулированной люцерной. На развитие и рост ребенка в течение года используется 0,7 % энергии, аккумулированной телятами. В результате чуть более одной миллионной доли солнечной энергии, падающей на поле в 4 га, используется для пропитания ребенка в течение одного года.
Биологическая продукция (продуктивность) экосистемы — прирост биомассы в экосистеме, созданной за единицу времени. В данном контексте более правильно пользоваться понятием «продуктивность». Однако вместо этого чаще используют понятие «продукция», все равно учитывая при этом и фактор времени. Различают первичную и вторичную продукцию сообщества.
Первичная продукция — биомасса, созданная за единицу времени продуцентами. Она делится на валовую и чистую. Валовая первичная продукция (общая ассимиляция) — это общая биомасса, созданная растениями в ходе фотосинтеза. Часть ее расходуется на поддержание жизнедеятельности растений — траты на дыхание (40— 70 %). Оставшаяся часть составляет чистую первичную продукцию (чистая ассимиляция), которая в дальнейшем используется консументами и редуцентами, или накапливается в экосистеме.
Вторичная продукция — биомасса, созданная за единицу времени консументами. Она различна для каждого следующего трофического уровня.
Масса организмов определенной группы (продуцентов, консументов, редуцентов) или сообщества в целом называется биомассой. Самой высокой биомассой и продуктивностью обладают тропические дождевые леса, самой низкой — пустыни и тундры (табл. 1).
Если в экосистеме скорость прироста растений (образования первичной продукции) выше темпов переработки ее консументами и редуцентами, то это ведет к увеличению биомассы продуцентов. Если при этом присутствует недостаточная утилизация продуктов опада в цепях разложения, то происходит накопление мертвого органического вещества. Это ведет к эвторфовыванию болот, образованию мощной лесной подстилки и т.п. В стабильных экосистемах биомасса остается постоянной, так как практически вся продукция расходуется в цепях питания.
Циклические изменения в экосистеме. Изменения в сообществах могут быть циклическими и поступательными.
Циклические изменения — периодические изменения в биоценозе (суточные, сезонные, многолетние), при которых биоценоз возвращается к исходному состоянию.
Суточные циклы связаны с изменением освещенности, температуры, влажности и других экологических факторов в течение суток и наиболее резко выражены в условиях континентального климата. Суточные ритмы проявляется в изменении состояния и активности живых организмов.
Сезонная цикличность связана с изменением экологических факторов в течение года и наиболее сильно выражена в высоких широтах, где велик контраст зимы и лета. Сезонная изменчивость проявляется не только в изменении состояния и активности, но и количественного соотношения отдельных видов. На определенный период многие виды выключаются из жизни сообщества, впадая в спячку, оцепенение, перекочевывая или улетая в другие районы.
Многолетняя изменчивость связана с флуктуациями климата или другими внешними факторами (степень разлива рек), либо с внутренними причинами (особенности жизненного цикла растений-эдификаторов, повторения массового размножения животных).
Сукцессии. Изменения в сообществах могут быть циклическими и поступательными. Поступательные изменения — изменения в биоценозе, в конечном счете, приводящие к смене этого сообщества другим. Сукцессия — последовательная смена биоценозов (экосистем), выраженная в изменении видового состава и структуры сообщества (рис. 4). Последовательный ряд сменяющих друг друга в сукцессии сообществ называется сукцессионной серией. К сукцессиям относятся опустынивание степей, зарастание озер и образование болот и др.
В зависимости от причин вызвавших смену биоценоза, сукцессии делят на природные и антропогенные, аутогенные и аллогенные.
Природные сукцессии происходят под действием естественных причин, не связанных с деятельностью человека. Антропогенные сукцессии обусловлены деятельностью человека.
Рис. 3. Сукцессия сибирского темнохвойного леса (пихтово-кедровой тайги) после опустошительного лесного пожара (обобщенная схема). Числа в прямоугольниках — колебания в длительности прохождения фаз сукцессии (в скобках указан срок их окончания). Биомасса и биологическая продуктивность показаны в произвольном масштабе. (Кривые отражают качественную и количественную стороны процесса.) (Н.Ф. Реймерс, 1990.)
Аутогенные сукцессии (самопорождающиеся) возникают вследствие внутренних причин (изменения среды под действием сообщества). Аллогенные сукцессии (порожденные извне) вызваны внешними причинами (например, изменение климата).
В зависимости от первоначального состояния субстрата, на котором развивается сукцессия, различают первичные и вторичные сукцессии. Первичные сукцессии развиваются на субстрате, не занятом живыми организмами (на скалах, обрывах, сыпучих песках, в новых водоемах и т.п.). Вторичные сукцессии происходят на месте уже существующих биоценозов после их нарушения (в результате вырубки, пожара, вспашки, извержения вулкана и т.п.).
В своем развитии экосистема стремится к устойчивому состоянию. Сукцессионные изменения происходят до тех пор, пока не сформируется стабильная экосистема, производящая максимальную биомассу на единицу энергетического потока. Сообщество, находящееся в равновесии с окружающей средой, называется климаксным.
Продуктивность экосистемы тесно связанна с потоком энергии, проходящим через нее. В каждой экосистеме только часть поступающей энергии накопляется в виде органических соединений. Скорость ассимиляции энергии называется продукцией, а величина продукции, отношение к единице площади экосистемы называется продуктивностью. Первичная продуктивность (Р) экосистемы определяется как скорость, с которой лучистая энергия усваивается продуцентами в процессе фото- и хемосинтеза накапливаясь в виде органических веществ, количество ее выражают в сырой или сухой фазе растений или энергетических единицах (ккал, Дж). Первичная продукция определяется общим потоком энергии через биотический компонент экосистем, а следовательно и биомасса живых организмов, которые могут существовать в биосистеме Величина создания первичных биологических продукций определяется возможностями фотосинтетического аппарата растений. Из общего количества лучевой энергии 44% составляет ФАР. – фотосинтез активная радиация т.е. свет по длине волны пригоден для фотосинтеза. Максимальная КПД фотосинтеза 10-12% ФАР, что является приблизительной половиной от теор возможного. По земному шару усвоение растениями солнечной энергии не превышает 0,1% из-за влияния на фотосинтез роста растений различных факторов: климатических, физических, химических.
В процессе производства органического вещества выделяют 4 последовательных уровня:
1 валовая первичная продуктивность – это общая В фотосинтеза с учетом органических веществ, которые за время измерений были израсходованы на дыхание (Р) .
2 чистая первичная продуктивность сообщества (Рчист) это - накапливание органического вещества в растительных тканях за вычетом органического вещества, которая израсходовалась на дыхание растений.
3 чистая продуктивность сообщества – это В накапливание органического вещества не потребленного гетеротрофами т.е. разность между чистой первичной продукцией и количеством органического вещества, потребленного гетеротрофами.
4 Вторичная продуктивность – накопление энергии на уровне консументов т.к. консументы используют ранее созданные питательные вещества часть из них расходуется на дыхание, а остальная часть на формирование тканей и органов (вторичную продукцию вычисляют отдельно для каждого проживающего уровня, т. к прирост массы для каждого из них происходит за счет энергии, поступающие предыдущем.
Постепенное увеличение общей биомассы продукции, происходит в том случае. Если В расхода первичной продукции в цепях питания отстает от темпов прироста растений. Биопродуктивность отличается от промышленной, т.к. процесс образования органического вещества в биологических сообществах непрерывен во времени. Поэтому продуктивность экосистем выражаем продукцией в граммах сухого вещества на 1м2 в сутки или год (гр м2/(сут) (год)). т. к. в экосистеме только растения ассимилируют солнечную энергиею превращая ее в органические вещества, а остальные только потребляют ее, то продукция экосистем в целом зависит от соотношения расхода энергии растениями на фотосинтез и дыхание. В неблагоприятных условиях расходы на дыхание организмов увеличивается, продукция экосистемы снижается. Мировое распределение первичной биологической продуктивности неравномерно.
Классификация сообществ по продуктивности. Уиттекер в 1979 году предложил классифицировать сообщества по продуктивности на 4 класса:
сообщества высшей продуктивности 3000 - 2000 г/м2 в год к таким сообществам относятся тропические леса, посевы риса и сок тростника. Запас биомассы в этом классе сообществ превышает 50 кг/м2 в местных сообществах равно продуктивности однолетних сельскохозяйственных культур.
Сообщество высокой продуктивности 200 – 1000 г/м2 гол листопадные леса умеренной зоны, луга при применении удобрений, посевы кукурузы, так биологически приближаются к биомассе 1 класса min. Биомасса равна чистой биологической продуктивности однолетних культур.
3 класс сообщества, сообщество умеренной продуктивности. От 1000 до 250 г м2/год к таким сообществам относятся основная масса сельхоз культур, кустарники, степи. Биомасса степей изменяется от 0,2 до 5 кг/м2.
последний класс сообщества с низкой производительностью (менее 250 г м2/год) пустыни, полупустыни, тундры. Обще годовая продуктивность сухого органического вещества на земле составляет 150-200 млн. т. из низ - 2/3 производится на суше и 1/3 приходится на океан. Практически вся первичная продукция служит для поддержания жизни гетеротрофных организмов. Питание людей обеспечивает в основном сельскохозяйственные культуры, которые занимают 10% всей площади суши. Годовой прирост культурных растений равен 16% от всей продукции суши. Половина урожая идет на питание людей, остальное на корм домашних животных и отходы. Всего человечество потребляет 0,2 % первичной продукции. Ресурсы, имеющиеся на земле, включая продукцию животноводства и рез промысла, на суше и на море могут обеспечить только 50 % потребности современного населения земли.
Биомасса — это выраженное в единицах массы или энергии количество живого вещества организмов (популяций, видов, группы видов, отдельных живых экологических компонентов, биотических сообществ в целом), приходящееся на единицу площади или объема. Для сопоставимости параметров биомассы разных организмов ее выражают в единицах сухого вещества. Совокупная масса организмов, присутствующих в различных экосистемах Земли (наземных и водных),
Еще одним важнейшим показателем автотрофного звена экосистем является первичная продукция, представляющая собой биомассу подземных и надземных органов, а также энергию и биогенные летучие вещества, произведенные автотрофной растительностью на единицу площади за единицу времени. Выражается обычно в граммах биомассы на 1 м2 за год.
Первичной валовой продукцией называют суммарную продукцию фотосинтеза (суммарную ассимиляцию), включающую, следовательно, и вещество, сжигаемое при дыхании за период наблюдения.
Чистая первичная продукция, или видимый фотосинтез, — это скорость накопления создаваемого органического вещества сверх того, которое затрачено на дыхание; чистая продукция — вещество, которое можно взвесить при уборке урожая.
При полном насыщении влагой и минеральным питанием дальнейший рост первичной продукции растений ограничивается количеством поступающей солнечной энергии. Повышение этого показателя возможно лишь при искусственном увеличении количества света. В условиях светокультуры, когда снабжение лучистой энергией превышает естественное освещение, продуктивность растений заметно возрастает.
Вторичная продукция экосистем в упрощенном виде представляет собой массу или заключенную в ней энергию органического вещества, образованного консументами — гетеротрофными организмами разных рангов и пищевых уровней при поедании ими первичных продуцентов (зеленых растений и водорослей).
Нередко употребляют термин общая продукция (общая продуктивность), обозначающий общее количество создаваемого в биоценозе живого вещества, т. е. сумму первичной и вторичной продукции. Однако следует иметь в виду, что вторичная продукция создается консументами за счет уничтожения части первичной продукции, как бы вычитаясь из нее. Вот почему общая продуктивность биоценоза может оцениваться только объемом первичной продукции, произведенной зелеными растениями. Можно сказать, что вторичная продукция — это скорость накопления массы или энергии в организмах на разных уровнях потребителей. Консументы лишь используют ранее созданные питательные вещества, часть из них расходуют на дыхание, а остальные превращают в биомассу собственного организма.
Общий поток энергии на гетеротрофном уровне, аналогичный валовой продукции на автотрофном уровне, называют не продукцией, а ассимиляцией. Поток энергии, проходящий через трофический уровень, представляет собой суммарную ассимиляцию на этом уровне, или сумму продуцируемой биомассы веществ, затраченных в процессе жизнедеятельности на дыхание. На рис.6 схематически показан поток энергии через три уровня простой пищевой цепи. При этом в соответствии со вторым законом термодинамики поток энергии с каждой ступенью уменьшается, так как при превращении одной формы энергии в другую часть ее теряется в виде тепла.
Вторичная продукция на каждом последующем трофическом уровне консументов составляет около 10 % предыдущей, очень редко достигая на уровне хищников 15—20 %. Обычно растительноядные животные потребляют около 10 % энергии, накопленной растениями, а плотоядные первого порядка — около 10 % энергии, содержащейся в биомассе растительноядных. Это значит, что эффективность плотоядных первого порядка по отношению к энергии, содержащейся в растениях, составляет 1 %, а по отношению к солнечной энергии — всего 0,001 %.
В целом же вторичная продуктивность во много раз меньше первичной и эффективность ее колеблется от 1 до 10 % в зависимости как от особенностей поедаемого корма, так и от самих животных. Между морскими и наземными экосистемами существует различие, состоящее в том, что во первых биомасса животных-потребителей больше биомассы микроскопических продуцентов, тогда как во вторых наблюдается обратное соотношение.
Это объясняется тем, что скорость репродукции автотрофного звена водной среды очень высока, благодаря чему обеспечивается постоянный приток биомассы первичных продуцентов в пищевую цепь. Несмотря на кажущуюся небольшую величину вторичной продукции, человечество обеспечивает себя необходимыми белками в сбалансированном пищевом рационе преимущественно за счет органических веществ вторичных продуцентов.
Лекция 14 Разнообразие экосистем биосферы
Екосистеми, що є в сучасній біосфері, належать до двох основних категорій. По-перше, це природні екосистеми, що виникають та існують незалежно від людини, по-друге, штучні антропогенні екосистеми, які створюються людиною (наприклад, посіви). Усі природні екосистеми тією чи іншою мірою змінені внаслідок господарської діяльності людини.
Природні екосистеми досить
різноманітні. Але за спільністю основних
структурних ознак, особливостями
функціонування, характером потоку
енергії та кругообігу речовин їх можна
об'єднати в декілька основних типів.
Практично при виділенні типів екосистем
перш за все беруть за основу ознаки
біоценозів. У цьому разі все різноманіття
екосистем світу можна звести до 15
основних типів (табл.
1, рис.2). Ці типи часто
називають біомами. Біоми — це великі
регіональні одиниці поділу біосфери,
які мають специфічну фауну та флору,
пристосувалися до певних ґрунтово-кліматичних
умов та сумісного життя. Найбільш
важливими для біосфери є біоми лісів
та різних типів злакової рослинності.
На лісові біоми припадає 27% території
суходолу (40,5 млн. км2).
Таблиця 1 - Класифікація основних природних екосистем світу
A |
Екосистеми суходолу |
|
1. Тундра |
|
2. Шпилькові ліси помірної зони |
|
3. Листяні ліси помірної зони |
|
4. Степи |
|
5. Тропічна та субтропічна злакова рослинність |
|
6. Савана |
|
7. Пустеля |
|
8. Вічнозелений тропічний дощовий ліс |
|
9. Болота |
|
10. Луки |
Б |
Екосистеми водойм |
|
11. Стоячі водойми: озера та ставки |
|
12. Екосистеми текучої води: ріки та ручаї |
|
13. Екосистеми відкритого океану |
|
14. Екосистеми континентального шельфу |
|
15. Естуарії |
На території України представлені зони змішаних лісів, лісостепи, степи, а також гірські біоми Карпат і Криму (рис. 2).
Р. Уиттекер (1975) залежно від обсягу біопродукції, що створюється в екосистемах, поділяв їх на чотири основні класи:
1. Екосистеми найвищої продуктивності, в межах 2000-3000 г/м2 на рік. До них належать екосистеми тропічних вологих лісів.
2. Екосистеми високої продуктивності - у межах 1000- 2000 г/м2 на рік. До них належать листяні ліси помірної зони та лук
3. Екосистеми помірної продуктивності - у межах 250-1000 г/м2 на рік. До них належать степи та чагарники.
4. Екосистеми низької продуктивності - менше 250 г/м2 на рік. До них належать пустелі та напівпустелі.
Різноманіття екосистем на нашій планеті є важливим фактором загальної стійкості біосфери.
Рис. 2. Схема фізико-географічного районування України.
Межі: 1 - країни, 2 - зони, 3 - підзони, 4 - провінції, 5 - підпровінції, 6 - області; МЛ - зона мішаних лісів; ЛС - лісостепова зона; С - степова зона; УК - Українські Карпати; КГ - Кримські гори (на схемі цифрами позначені області).
Південний захід Східно - Європейської рівнини. Зона мішаних лісів. Поліська провінція: 1 - Волинське Полісся; 2 - Мале Полісся; 3 - Житомирське Полісся; 4 - Київське Полісся; 5 - Чернігівське Полісся; 6 - Новгород-Сіверське Полісся. Лісостепова зона. Західно-Українська лісостепова провінція: 7 - Волинське лісостепове Опілля; 8 - Розточчя і Опілля; 9 - Захід-но-Подільський лісостеп; 10 - Північно-Подільський лісостеп; 11 - Прут-Дністровський лісостеп. Дністровсько-Дніпровська лісостепова провінція: 12 - Північно-Придніпровський лісостеп; 13 - Київський підвищений лісостеп; 14 - Придністровсько-Подільський лісостеп; 15 - Подільсько-Побузь-кий лісостеп; 16 - Центральнопридніпровський підвищений лісостеп; 17 -Південно-Подільський підвищений лісостеп; 18 - Південно-Придніпровсь-кий підвищений лісостеп. Лівобережно-Дніпровська лісостепова провінція: 19 - Північно-Дніпровський терасно-рівнинний лісостеп; 20 - Південно-Дніпровський терасно-рівнинний лісостеп; 21 - Північний Полтавський лісостеп; 22 - Південний Полтавський лісостеп. Середньоруська підвищена лісостепова провінція: 23 - Сумський підвищений лісостеп; 24 - Харківський підвищений лісостеп. Степова зона. Північностепова підзона. Дністровсько-Дніпровська північностепова провінція: 25 - степові відроги Центрально-молдавської височини; 26 - степові відроги Подільської височини; 27 –степові відроги Придніпровської височини. Лівобережно-Дніпровсько-При-азовська північностепова провінція: 28 - Орельсько-Конкський низовинний степ; 29 - Приазовський підвищений степ; 30 - Приазовський низовинний степ. Донецька північностепова провінція: 31 - степові західні відроги Донецької височини; 32 - Донецький підвищений степ. Задонецько-Донська північностепова провінція: 33 - Старобільський степ. Середньостепова підзона. Причорноморська середньостепова провінція: 34 - Задністровський низовинний степ; 35 - Дністровсько-Бузький низовинний степ; 36 - Бузько-Дніпровський низовинний степ; 37 - Дніпровсько-Молочанський низовинний степ; 38 - степові південно-західні схили Приазовської височини. Сухо-степова підзона. Причорноморсько-Приазовська сухостепова провінція: 39 - Приморський низовинний степ; 40 - Нижньодніпровський терасно-дель-товий степ; 41 - Присивасько-Приазовський низовинний степ. Кримська степова провінція: 42 - Кримсько-Присиваський низовинний степ; 43 - Тар-ханкутський підвищений степ; 44 - Центральнокримський рівнинний степ; 45 - Керченський горбкуватий степ.
Українські Карпати: 46 - Передкарпаття; 47 - Зовнішні Карпати; 48 - Вододільно-Верховинські Карпати; 49 - Полонинсько-Чорногірські Капрати; 50 - Рахівсько-Чивчинські Карпати; 51 - Вулканічні Карпати; 52 - Закарпатська низовина. Кримські гори: 53 - Передгірний лісостеп; 54 - Головна гірсько-лугово-лісова гряда; 55 - Кримське південнобережне субсередземномор'я.
ТУНДРИ. Екосистеми тундр розміщуються переважно у Північній півкулі, на Євро-Азіатському та Північно-Американському континентах у районах, що межують із Північним Льодовитим океаном. Загальна площа, яку займають екосистеми тундр та лісотундри у світі, дорівнює 7 млн. км2 (4,7% площі суходолу).
Клімат тундр дуже суворий. Середня добова температура вище 0°С спостерігається протягом 55-118 діб на рік. Вегетаційний період починається в червні та закінчується у вересні. Кількість опадів незначна - 200-400 мм на рік, але вологість ґрунту влітку досить висока внаслідок низького випаровування (на нього витрачається тільки 30% загальної кількості опадів) та наявності вічної мерзлоти. Суворість клімату тундри посилюють постійні сильні вітри, у зимовий період під їх впливом відмирають усі частини рослин, що розташовуються над поверхнею снігу. Тундри Євразії та Північної Америки мають багато спільного. Ареали багатьох видів рослин та тварин тундр охоплюють обидва континенти. Аналогічні кліматичні умови складаються у високогір'ї, де формуються гірські тундрові екосистеми, схожі на зональні.
Рельєф поверхні тундри в основному рівний. Ґрунти слабо розвинуті, торфові та болотисті. Вони погано прогріваються, і тому процеси гуміфікації та мінералізації відбуваються в них повільно. Ґрунти тундри завжди кислі, і вміст гумусу в них не перевищує 1-2%.
Тундра безліса. У рослинному покриві переважають низькорослі чагарники - карликова берізка, приземисті види верби, чорниця, лохина та водяниця. Місцями ростуть осоки та пухівка. Основний фон рослинного покриву складають кущисті лишайники та мохи. Значно поширені види лишайників з родів Cladonia та Cetraria. До них належить відомий оленячий мох -ягель. Вищі рослини тундр звичайно представлені багаторічними видами, що мають потовщені підземні частини з запасами поживних речовин, завдяки яким забезпечується раннє весняне відростання та швидке цвітіння. Ця особливість тундрових рослин дуже важлива в умовах короткого літа.
Тваринний світ екосистем тундр бідний. Його формування обмежує злиденний запас рослинної їжі та суворість клімату. На зиму більшість мешканців тундри мігрує в лісову зону, птахи відлітають на південь. Постійно зимує невелика кількість видів: лемінги, деякі ховрахи, песці, полярна сова. У багатьох місцях ходи-нори лемінгів займають до 20% площі. БЛ. Тихомиров (1959) показав, що вони сприяють накопиченню на поверхні ґрунту рослинних залишків та надають поверхні ґрунту дрібнобугристого вигляду. Улітку життя тундр оживляє маса водоплавних птахів (гуси, казарки, качки, кулики), але вони включаються в тундрові трофічні ланцюги тільки частково, оскільки харчуються в основному на прибережних водоймах. Однак гуси та качки використовуть в їжу до 50-80% рослинності тундри в місцях свого гніздування. Найбільше вони пошкоджують квіткові бруньки.
Постійними мешканцями тундри є північні олені. Але це кочові тварини. Вони залишають тундру на зимові місяці, коли з-під твердого снігового покриву вони не можуть здобувати ягель та мох - основну свою їжу. Середину літа північні олені проводять на узбережжі, де достатньо кормів та вітер відганяє кровососних комах. Такий характер має кочівля й свійських північних оленів. У результаті перевипасу оленями мохолишайникові тундри перетворюються в лугові з переважанням на них щучника та тонконога.
Хижаки тундри представлені совами, песцями, частково білим ведмедем. У цілому вони нечисленні. Чисельність хижаків у різні роки помітно змінюється, повторюючи відповідні зміни чисельності травоїдних тварин, особливо лемінгів. В ентомофауні тундр переважають справжні комарі та мошки, їхні личинки живляться відмерлими рослинними залишками, а дорослі форми - кровососи.
Ґрунтові тварини заселяють тільки поверхневі частини ґрунту. У деструкції органічної речовини в екосистемах тундри перше місце посідають не бактерії, а гриби.
У цілому вплив тварин на рослинний покрив тундр досить великий. На прикладі тундр добре помітна взаємозумовленість існування цих двох груп живих організмів. Трофічна мережа тундрових екосистем наведена на рис. 5.2.
Вона досить сильно спрощена та включає тільки основні форми, на прикладі яких можна простежити основні трофічні ланцюги, властиві цій екосистемі.
Первинна продуктивність екосистем тундр невелика і становить у середньому 140 г сухої органічної речовини на 1 м2 за рік. Валова біологічна продуктивність у тундрах не перевищує 340 г/м2 за рік. Запаси фітомаси коливаються від 0,1 до 100 тонн на гектар.
Тундрові екосистеми характеризуються нестійкістю та вразливістю. Порушення цих екосистем зберігаються досить довго, відновні процеси йдуть поволі.
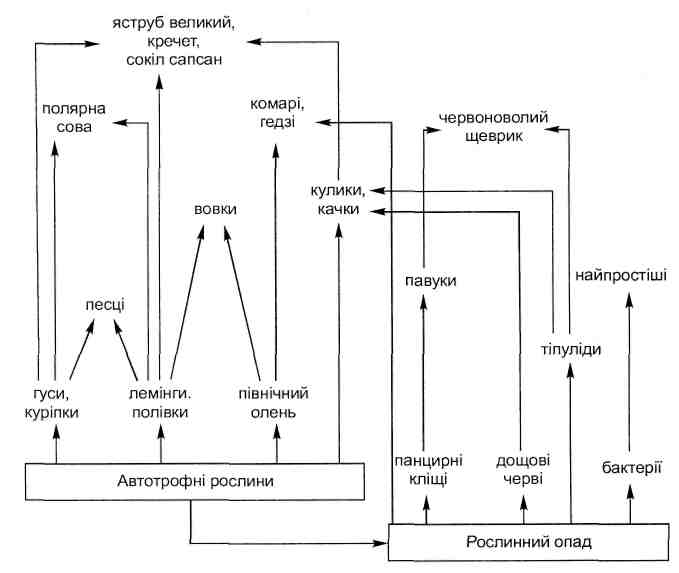
Рис. 3. Спрощена трофічна мережа екосистеми тундри
ЛІСОВІ ЕКОСИСТЕМИ ПОМІРНОГО ПОЯСУ. За визначенням Ю.Р. Шеляг-Сосонка (2001), ліс - це сукуність більш чи менш зімкнутих деревних угруповань на певній території або тип рослинності, в якому провідну роль відіграють більш чи менш зімкнуті деревостої. Загальна площа лісів у світі приблизно 4,2 млрд. га, що становить 37% поверхні суші. У них зосереджено 1,7 1012 сухої органічної речовини. Це майже 90% усієї біомаси, яка є на континентах. Ліс - це свого роду каркас біосфери, її екологічний остов. Екологічний вплив лісів має багатобічну дію. Так, наприклад, соснові ліси поглинають 9-15 т/рік вуглецю у формі вуглекислого газу, відповідно збагачуючи повітря киснем. Одночасно вони утримують на своєму листі до 38 т пилу, знижуючи запиленість повітря.
Лісові екосистеми займають на земній кулі великі площі. В їхньому рослинному покриві переважають дерева. Залежно від ґрунтово-кліматичних умов та географічного положення лісові екосистеми поділяються на тайгу, змішані та листяні ліси.
Тайга. Тайгою називають шпилькові ліси, які широкою смугою простягаються на Євро-Азіатському та Північно-Американському континентах південніше лісотундри. Екосистеми тайги займають 13,4 млн. км2, що становить 10% поверхні суходолу або 1/3 всієї лісовкритої території Землі.
Для екосистем тайги характерна холодна зима, хоча літо досить тепле і тривале. Сума активних температур у тайзі становить 1200-2200°С. Зимові морози сягають 30° - 40°С нижче нуля. Кількість опадів коливається від 300 до 1600 мм на рік. Ґрунтові процеси внаслідок тривалої зими йдуть малоактивно,гуміфікація сповільнена. Ґрунти переважно підзолисті.
Деревостій у тайзі представлений ялиною сибірською або європейською, ялицею, соснами та модриною. У Північній Америці їх замінюють тсуга та псевдотсуга. Усі ці види в цілому маловибагливі щодо родючості ґрунту. Модрина переважає на сході Євразії, сосна тяжіє до сухих або заболочених ґрунтів. Для нашої планети екологічно важливими є соснові ліси, вони займають друге місце після вологих тропічних лісів за обсягом продукування газоподібного кисню в розрахунку на одиницю поверхні ґрунту.
Кореневі системи дерев, як правило, поверхневі (окрім сосни). Це робить тайгу нестійкою до посух та схильною до буреломів. Зімкнутість крон дерев висока, і на землю проникає мало світла. Тому в тайзі слабо розвинуті підлісок та трав'яний покрив. Ґрунт вкритий зеленими мохами, а в більш вологих місцях - сфагнумом. На моховому покриві часто ростуть дрібні чагарнички - брусниця, чорниця, лохина та мучниця.
Після вирубок чи пожеж на місці шпилькового деревостою спочатку оселяються дрібнолисті деревні породи - береза чи осика. Пізніше під їхнім покривом з'являються відновлені шпилькові породи, які з часом переростають та витісняють менш довговічні дрібнолисті види дерев. Такий тип відновлення шпилькового деревостою називають відновленням через заміну порід.
Фауна екосистем тайги багатша, ніж тундри. Тут трапляється багато видів травоїдних ссавців та рослиноїдних птахів. Типовими для тайги є бобри, лосі, білки, заєць-біляк, бурундуки, глухарі, тетеруки, рябчики. Важливим видом кормів є насіння шпилькових порід дерев, яким харчується велика кількість видів гризунів та птахів. Врожаї насіння шпилькових порід досить сильно змінюються в різні роки, тому спостерігаються кочівлі тварин, коли в пошуках корму білки, шишкарі, горіхівки та інші тварини, що харчуються насінням, переміщуються на великі віддалі. Важливим джерелом корму в тайзі є бруньки дерев та чагарників, їх широко використовують в їжу снігурі, смеречни-ки, омелюхи, вівсянки, тетеруки, глухарі та рябчики. Значну чисельність мають тут і комахи-фітофаги, зокрема ті, що живляться деревиною. На деревах селиться велика кількість видів паразитичних та сапрофітних грибів. Є тут також тварини-ризофаги, які живляться корінням. Найбільш характерні в цій групі дротяники - личинки жуків-коваликів.
У підстилці та поверхневому шарі ґрунту немало фітофагів, з яких починається детритний ланцюг живлення. Це кліщі орибатіди, ногохвістки, нематоди, дощові черви.
Фауна хижаків представлена колонком, риссю, соболем, росомахою, вовком, бурим ведмедем, лисицею. Чимало тут комахоїдних птахів - дятли, повзики, дрозди, синиці та ін. Є тут також земноводні та плазуни.
У тайзі зустрічається багато кровососних комах (кліщі, комарі, мошки). У цілому трофічна мережа тайги багатша та складніша, ніж у тундрі. Трофічні ланцюги довші й мають паралельні ділянки (рис. 4). Тому екосистеми тайги порівняно з тундровими більш стійкі до різних порушень.
Біомаса екосистем тайги складає 350-400 тонн/га, а річна продукція - 8-10 тонн/га. В умовах більш континентального клімату, де переважає модрина, а деревостої розріджені, біомаса знижується до 50-200 тонн/га, а річна продукція - до 4-6 тонн/га. Екосистеми тайги можуть також формуватися в гірських масивах і є там одним із гірських поясів. Такі типи екосистем можна спостерігати в Карпатах. Тут вони утворені ялиною європейською, білою ялицею та сосною. Деревостої частіше одноярусні, підлісок відсутній. Як зауважував МЛ. Голубець (1971), ялиця в Українських Карпатах не утворює однодомінантних угруповань. Для фауни характерна присутність ізольованих популяцій таких видів, як глухар та довгохвоста сова. В.Б. Сочава (1956) оцінював карпатські шпилькові ліси не як тайгові, а як неморально-темношпилькову зональну формацію.
На користь цієї точки зору свідчить помітний генетичний зв'язок цих формацій з буковими лісами.
Середньоєвропейські соснові ліси в Україні поширені на крайньому заході, у Розточчі.
Серед загальних характеристик усіх формацій тайги можна назвати:
а) монодомінантність (деревостій формується лише з однієї породи дерев);
б) однорідність та одноманітність на великому просторі умов, що створюються під покривом лісу;
в) проста вертикальна та горизонтальна будова рослинного покриву;
г) переважання мезофітів;
д) некомпенсованість кругообігу мінеральних речовин.
За А.І. Толмачовим (1954), екосистеми тайги історично молоді, вони сформувалися в другій половині третичного періоду на місці субтропічних лісів. Таке походження проявляється у великій кількості вічнозелених видів рослин.
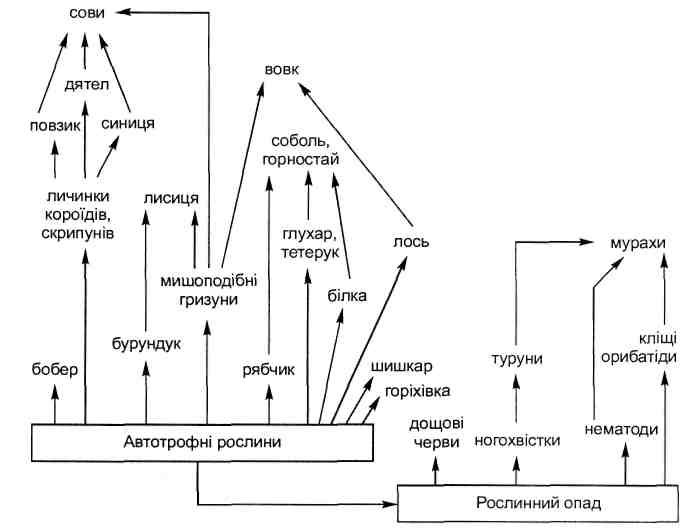
Рис. 5 - Спрощена схема трофічної мережі екосистеми тайги
Мішані та листяні ліси помірної зони. Екосистеми цього виду поширені на південь від зони тайги. Вони охоплюють майже всю Європу, простягаються більш чи менш широкою смугою в Євразії, добре виражені в Китаї. Є ліси такого типу й в Америці.
Кліматичні умови в зоні листяних лісів м'якші, ніж у зоні тайги. Зимовий період триває не більше 4-6 місяців, літо тепле. На рік випадає 700-1500 мм опадів. Ґрунти підзолисті.
Листовий опад сягає 2-10 тонн/га на рік. Він активно залучається до гуміфікації та мінералізації. Тому ґрунти листяних лісів більш багаті на гумус та мінеральні речовини, ніж ґрунти в тайзі. Запас гумусу досягає 10-20 тонн/га. У фауні ґрунтових фітофагів переважають дощові черв'яки, але є чимало нематод, ківсяків, багатоніжок, кліщів-орибатід.
Для екосистем, утворених листопадними породами, характерний різко контрастний режим освітленості. Узимку та напровесні, коли дерева та чагарники стоять без листя, освітленість на рівні ґрунту висока; улітку, навпаки, досить високе затінення. Це призводить до появи в листяних лісах особливої синузії весняних ефемероїдів. Період їх активної життєдіяльності припадає на ранню весну, коли температура вже підвищилася, але дерева ще не встигли одягнутися в листя.
Різноманіття видів дерев та чагарників в зоні листяних лісів дуже велике. В Європі можна виділити три основні зони за переважаючими лісоутворюючими породами. У Західній Європі переважають ліси дуба звичайного з домішками сосни, берези, вільхи, осики та клену. У Центральній Європі ліси утворені буком європейським, грабом та липою. На західному узбережжі Європи, у Франції та Великій Британії більш поширені дуб звичайний та дуб скелястий, граб.
Ярусна структура листяних та мішаних лісів більш складна, ніж тайги. Верхній ярус утворюють великі високорослі дерева, часто є й другий ярус деревостою, добре розвинутий також ярус чагарників. У ньому найчастіше зустрічається ліщина, вовче лико, жимолость, калина та горобина. При утворенні розрідженого верхнього ярусу сильно розростається травостій із конвалії, яглиці, зеленчуку, копитняку та осоки. Моховий покрив звичайно розвинутий досить слабо внаслідок пригнічення його росту великим листовим опадом. Найбільш складну будову мають ліси Білорусії та Правобережної України.
Тваринне населення екосистем листяних лісів досить різноманітне. Характерна наявність великих рослиноїдних видів ссавців - благородного оленя, лося, бізона. Типовим для листяних лісів є кабан. У мішаних та листяних лісах живе велика кількість видів птахів. Це сойки, дубоніс, дятли та сови. Частина їх належить до фітофагів, інші - до хижаків. Велика кількість видів харчується насінням, плодами та бруньками. Група комахоїдних птахів представлена дроздами, мухоловками, кропив'янками, вівчариками та синицями. З хижаків тут є бурий ведмідь, рись, вовк, росомаха, лисиця. Деякі птахи та ссавці мігрують на зиму в більш теплі місця, де менше снігу та легше добути їжу.
У групі лісових комах багато ксилофагів, що харчуються живою та мертвою деревиною. Є комахи, що пристосувалися поїдати листки (наприклад, дубова листовійка). Часто зустрічаються богомоли та цикади. У ґрунті є багато видів ризофагів: дротяники - личинки жуків-коваликів та личинки пластинчастовусих жуків і особливо хрущів. Серед комах чимало хижих форм: туруни, хижі кліщі та багатоніжки, оси-наїзники та ін. У цілому для екосистем змішаних та листяних лісів характерні складні трофічні мережі (рис. 5.4).
Запаси біомаси в листяних лісах становлять 400-500 тонн/га при річній продуктивності 10-50 тонн/га. На зоомасу припадає до 1 тонни/га, що перевищує цей показник в усіх інших біомасах суходолу
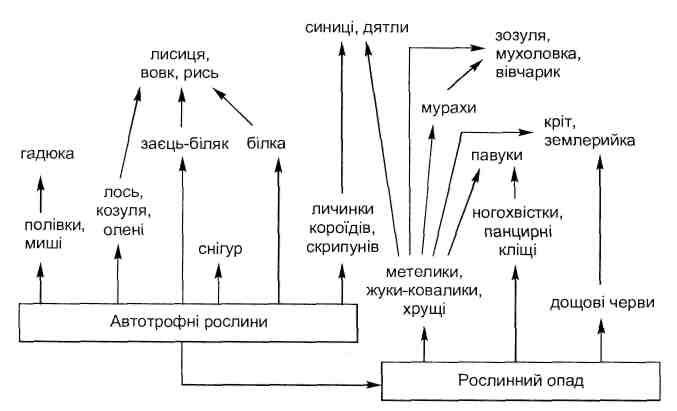
Рис. 6 - Спрощена трофічна мережа екосистеми змішаного лісу
Південний кордон поширення листяних лісів визначає дефіцит вологи та засолення ґрунту. Тут ліси поступово переходять у лісостеп, а далі - у степи.
Екосистеми листяних та мішаних лісів розташовані в найбільш сприятливому кліматі, де здавна селилися люди. Це привело до того, що величезні масиви таких лісів були вирублені. У середньому вже втрачено більше 3/4 площі мішаних та листяних лісів помірної зони. У США 3/4 таких лісів вирубано, у Китаї - 90% . У лісах, що збереглися, спостерігається збідніння флори та фауни. У Північній Америці повністю знищений мандрівний голуб, майже зник бізон. У той же час збільшення площ галявин та зріджування деревостою стало причиною проникнення до лісів степових видів тварин: зайців, хом'яків, куріпок, польового жайворонка, ховраха та куниці.
В Україні ліси займають 15,0% усієї території. Тут розпізнають 28 лісових формацій. Відповідно до лісових деревних порід вони розподілені таким чином: соснові ліси складають 33,6% загальної території, ялинові - 9,8%, ялицеві -1,4%, дубові бори - 26,1%, букові ліси - 9,8%, вільхові - 4,3%, березові -5,6% . В Україні налічується більше 200 видів дерев та чагарників. Сучасна Україна бідна на ліси. Але в V ст. до н.е. давньогрецький історик Геродот після своїх подорожей у регіонах, що лежать зараз на півдні України, дав їм назву «Галлія», що означає «лісова країна».
наш час ліси зосереджені переважно у північній та північно-західній частинах країни, включаючи гірські райони Карпат та Криму. Лісам Карпат та Криму характерна вертикальна поясність.
У рівнинних лісах України за характером рослинного покриву та представниками тваринного світу можна виділити шість великих регіонів.
Західне Полісся займає північну частину Волинської та Рівненської областей. Це найбільш лісисті регіони, де на дерново-підзолистих ґрунтах ростуть соснові, березові та вільхові (чорна вільха) ліси. Центральне Полісся займає північну та середню частину Житомирської області. Лісоутворюючими породами тут є сосна, береза, чорна вільха та дуб. Східне Полісся охоплює північну частину Київської, Чернігівської та Сумської областей. Ліси тут в основному складаються з сосни та дуба. Мале Полісся розташоване на території Львівської, Рівненської, Тернопільської та Хмельницької областей із прилеглими передгір'ями Карпат. У лісах цього регіону ростуть сосна, дуб, береза, бук, чорна вільха.
Гірський лісовий регіон Карпат відрізняється досить різноманітними лісовими екосистемами. Тут ліси займають до 1 млн. га площі. Головними лісовими породами цього району є ялина європейська та біла ялиця. У передгір'ях Карпат значні площі зайняті дубовими лісами.
Гірському лісовому регіону Криму також властиве велике різноманіття лісів, які займають до 223,6 тис. га площі. Ліси розташовані здебільшого на північних схилах. В їх формуванні бере участь понад 100 видів дерев та чагарників. Є тут і ендеміки, як, наприклад, кримський бук. На південних схилах ліси утворені фісташкою. За даними Я.П. Дідуха (1992), у Криму є 14 формацій лісів та 2 формації рідколісся. У цілому серед інших регіонів України найбільш багате лісовими екосистемами Полісся
ВІЧНОЗЕЛЕНИЙ ТРОПІЧНИЙ ДОЩОВИЙ ЛІС. Тропічні дощові ліси, що інколи називають гілеєю або джунглями, формуються в умовах досить вологого та жаркого клімату. Сезонність тут не виражена, і пори року розпізнаються за дощовим та відносно сухим періодами. Середня місячна температура цілорічно утримується на рівні +24°-26°С та не опускається нижче +18°С. Опадів випадає в межах 1800-2000 мм на рік. Відносна вологість повітря звичайно перевищує 90% . Тропічні дощові ліси займають площу, рівну 10 млн. км2. У фітомасі тропічного лісу утримується 40% усього зв'язаного вуглецю планети. Основні масиви таких лісів розміщені в басейні ріки Амазонки (Південна Америка), у басейні ріки Конго (Африка) та на південному сході Азії.
Ґрунти слаборозвинуті, бідні на поживні речовини внаслідок швидких процесів гуміфікації та мінералізації. Усі поживні речовини швидко перехоплюються коренями або вимиваються з ґрунту зливами. Корені рослин проникають у ґрунти не глибше 50 см. Такі ґрунти після вирубки лісу та сільськогосподарського використання швидко піддаються ерозії та втрачають родючість.
Загальною особливістю тропічного дощового лісу є надзвичайно велике розмаїття видів рослин та тварин. Тут представлено майже 50% світового генофонду рослин і 2/3 видів тварин планети. Так, у південноамериканських гілеях нараховується тільки дерев понад 3 тисячі видів. У середньому, за Ф. Фіткау (1988), на 1 га тропічного дощового лісу припадає 90 тисяч видів живих організмів. Майже всі рослини тут вічнозелені. Тривалість життя окремого листка складає 12-14 років, опадають вони поступово, і рослини завжди стоять вкриті листям. Частина листків має пристосування до всмоктування дощової води, інші, навпаки, пристосовані до швидкого стікання води з їхньої поверхні вздовж спеціальних жолобків та виростів.
Набір різних життєвих форм рослин дуже різноманітний. Тут представлені як карликові (2-3 м заввишки), так і гігантські (30-45 м і навіть до 70 м) форми. Трапляється багато різновидів ліан, стебла яких завдовжки іноді сягають 240 м. Широко представлені епіфіти, що оселяються на стовбурах та гілках дерев. На епіфіти особливо багата Америка. Цвітіння рослин тропічного дощового лісу спостерігається протягом усього року.
Тропічні дощові ліси мають складну багатоярусну структуру рослинного компонента, яка впливає на ярусне розміщення тварин. На поверхню ґрунту потрапляє мало світла, воно перехоплюється верхніми ярусами, і тому трав'яний покрив зріджений або зовсім відсутній. Усе життя такого лісу зосереджене на верхніх ярусах.
Фауна тропічного дощового лісу також різноманітна. Тут багато видів комах, плазунів та птахів. Ссавці представлені значно менше, великих видів мало. Серед них трапляються дрібні олені, антилопи, тапіри. Переважають гризуни, що живляться рослинною їжею. У тропічному дощовому лісі є багато видів мавп. Особливого колориту екосистемам тропічного дощового лісу надають великі види метеликів, терміти, мурахи. Тут є багато видів кліщів, тарганів, п'явок, ногохвісток, скорпіонів, павуків.
Трофічна мережа дощового лісу відрізняється великою складністю (рис. 7). Жива біомаса тропічного лісу складає 350-700 і навіть 1000 тонн/га, опаду — до 100 тонн. Тваринна біомаса сягає 200-300 кг/га, або 0,02% усієї біомаси. На частку хребетних припадає 13-20 кг/га. У цілому тут найвища щільність біомаси на планеті. Екосистеми тропічного дощового лісу є важливими постачальниками газоподібного кисню: вони продукують його в кількості 30 тонн/га на рік. Поглинання вуглецю та виділення кисню в тропічних лісах зрівноважені.
Екосистеми тропічного дощового лісу за всієї їх складності досить нестійкі і легко можуть бути зруйновані під впливом господарської діяльності людини. Найбільших збитків завдають вирубки цінних порід деревини, наприклад, червоного дерева, махогонії, білого кедра, бальси, ебенового дерева. Для лісозаготівлі та потреб перелогової системи землекористування щороку вирубається майже 7,1 млн. га лісу. Зокрема, в Африці початкова площа тропічного дощового лісу скоротилася більш як на 60%. Відновлення ж екосистем тропічного лісу відбувається поволі та вимагає десятків, якщо не сотень років.
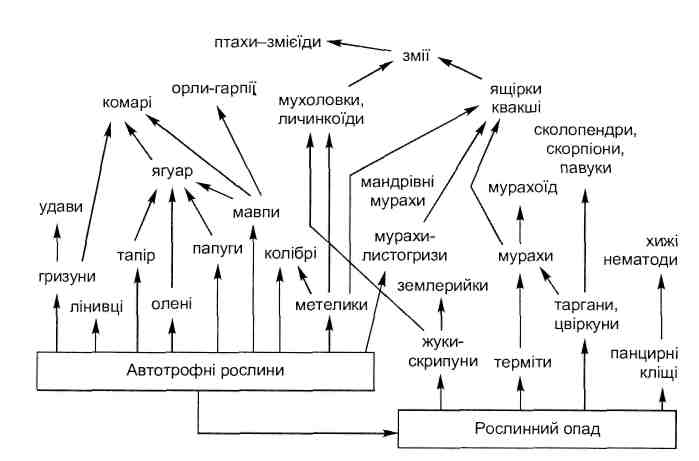
Рис. 7. Спрощена трофічна мережа тропічного дощового лісу
СТЕПИ. Степові екосистеми формуються в помірному поясі в умовах посушливого клімату і тому мають внутрішньоконтинентальне розташування. Середньорічна температура дорівнює +3-7,5°С. Опадів випадає за рік від 250 до 750 мм. Зволоження є головним фактором, що визначає розвиток рослинного покриву.
У Північній півкулі степова зона розташовується на південь від лісової та широкою смугою тягнеться в центрі Євразії. У місцях з океанічним типом клімату степи виклинюються та заміщуються іншими екосистемами. Аналогічне розташування мають степи і на Північно-Американському континенті (тут їх називають преріями). У Південній півкулі аналогом степів є пампа та злакові рослинні угруповання.
Ґрунти степів - це потужні чорноземи (тільки в південній частині їх замінюють бідні чорноземи та каштанові ґрунти). Підстилка завжди незначна, вона швидко муміфікується. Але швидкість мінералізації тут низька. Це є причиною накопичення потужних шарів гумусу. Його тут у 5-10 разів більше, ніж у лісовій зоні. Корені рослин проникають у ґрунт на глибину до 2 м.
Рослинний покрив степів формується з багаторічних трав. Головним чином це злакові. Рослинному покриву степів характерна полідомінантність та багатоярусність травостою. Є в степах і чагарники та чагарнички (степовий мигдаль, таволга, терен, степова вишня), але суцільного ярусу вони не утворюють. Усі рослини степів несуть на собі ознаки пристосованості до недостатності вологозабезпечення. У них є опушення, восковий покрив на листках, глибокі кореневі системи. Своєрідною є життєва форма перекотиполе. Такі рослини (їх більше 10 видів) з настанням осені відламуються на рівні ґрунту та завдяки кулеподібній формі переганяються вітром на значні відстані, постійно розсіюючи насіння. Степам характерне почергове цвітіння різних видів рослин, що проявляється в послідовній зміні аспектів. Протягом вегетаційного періоду їх буває 8-10. Видове різноманіття в степах досить значне, на 1 м2 реєструється до 80 видів квіткових рослин
У північних частинах степів переважають мезофітні крихко-дернові та кореневищні злаки, у південних їх замінюють дерно-винні. Північні степи іноді називають луговими, або ковилково-різнотравними. Тут звичайні ковила та типчаки, до складу різнотрав'я входять жовтецеві, свербіжниця, підмаренник, перстач та шавлія. Південний степ завжди має переважну більшість злаків, які представлені різнотравно-типчаково-ковилковими, типчако-во-ковилковими та полинно-злаковими формаціями. Південним степам характерна синузія ефемероїдів, що проходять основний життєвий цикл навесні, тобто в період найбільшої зволоженості. П. Халтенорт (1988) поділяє степові екосистеми на дві головні категорії: а) високотравні степи (розташовуються в місцях із річною кількістю опадів понад 300 мм та мають товстий горизонт гумусу); б) низькотравні степи (формуються в основному на каштанових ґрунтах та мають вміст гумусу 2-3%). Тваринний світ сучасних степів сильно збіднений та фрагментований.
У нижніх ярусах степів травоїдні тварини представлені гризунами, що живляться насінням, це ховрахи та байбаки. Тут також поширені гризуни-ризофаги, що поїдають корені, - кроти-сліпаки. Вони переміщують величезні об'єми ґрунту в пошуках корму і створюють у степах особливий мікрорельєф. Існує в степах кілька видів рослиноїдних птахів з роду куроподібних (луговий тетерук, сіра куріпка, перепілка), а також жайворонки. Є тут і всеїдні птахи, наприклад, дрофа та хохітва. Хижі птахи представлені степовим орлом. Звичайними для степу є гадюки, ящірки, степова черепаха. Степові птахи ряду видів віддають перевагу не польоту, а бігу. Так вони менш помітні для хижаків. У травостої проходить активне життя комах-фітофагів та хижаків: сарани, цикад, пінявок, клопів, трипсів, метеликів. Степи України зберігали багату фауну до 30-х років XX століття, але після колективізації та ліквідації меж суцільна оранка знищила місця існування багатьох видів тварин. Початкова цілина мала високе видове різноманіття тварин. Тут мешкали численні стада великих ссавців: дикі коні, антилопи, зубри, тури.
О.М. Формозов (1950) підкреслював особливу корисність для степових екосистем комахоїдних птахів. Вони запобігають нищівним спалахам чисельності саранових та інших видів комах. Хижаки контролюють чисельність гризунів. Сучасне скорочення кількості хижих птахів у степах привело до перетворення такого звичайного степового фітофага, як малий ховрах, на небезпечного шкідника сільського господарства.
Через чітко виражену зміну літнього та зимового сезонів у ряду тварин спостерігається зимова сплячка. Спрощена трофічна мережа степової екосистеми наведена на рис. 8.
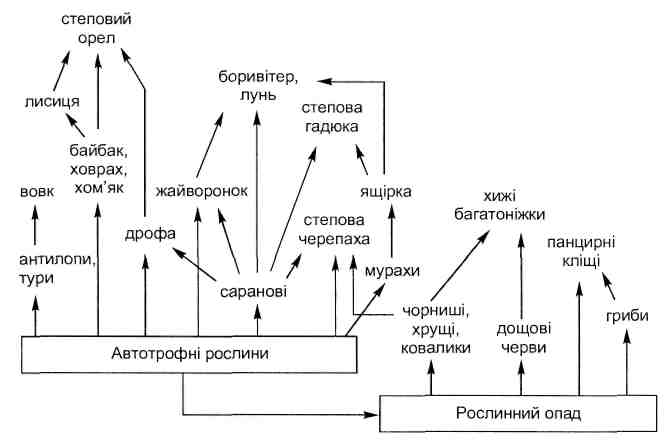
Рис. 8. Спрощена схема трофічної мережі степової екосистеми
Для північної частини степів, де вони переходять у лісостеп, характерне своєрідне спільне мешкання степових та типово лісових тварин. Тут поруч можна побачити білу куріпку та тушканчика, рись та дрофу, білку та ховраха. У лісостепу тварини звичайно полюбляють селитися та жити в лісі, а здобич шукати в степу. Така взаємопроникність степових та лісових екосистем є додатковою ілюстрацією континуальності біосфери. На Україні Ю.Р. Шеляг-Сосонко та ін. (1991) виділили 39 основних степових формацій рослинності. Основними є лугові, різнотравно типчаково-ковильні та пустельні полинно-злакові степи. У степах України багато видів ковилів, вони переважають на цілинних ділянках степу. Справжніх степових ділянок збереглося небагато. Це Михайлівська цілина, яка є луговим степом, Хомутов-ський степ з типовими приазовськими формаціями, Кам'яні Могили, залишок кам'янистого різнотравно-типчаково-ковило-вого степу, Асканія-Нова, де збереглося 11 тисяч га типчаково-ковилового степу та деякі інші.
Біомаса степових екосистем помірної зони вимірюється в межах 10-150 тонн/га, у середньому - 50 тонн/га. Річна біопродук-ція дорівнює 5-30 тонн/га. На долю зоомаси припадає 10-50 кг/га. У різні роки рівень біопродукції змінюється від 36 до 72 ц/га.
Висока родючість ґрунту степів та сприятливий клімат створюють оптимальні умови для землеробства. Основна маса степових систем унаслідок цього нині є повністю розораною.
ПУСТЕЛІ. Пустелі формуються в умовах континентального клімату з різкою перевагою випаровування над опадами. Розташовані вони в основному в тропічному та субтропічному поясах. Типова пустеля - це спекотна, суха територія. Так, у пустелі Сахара зареєстрована температура повітря в затінку +58°С, у каліфорнійській пустелі Долина Смерті —1-56,5°С. У зв'язку з континентальним положенням для пустель характерні значні добові коливання температур, наприклад, уночі в Сахарі прохолодно і температура може знижуватися до +10-12°С. Кількість опадів не перевищує 250 мм на рік, а перевищення випаровування над опадами у 5— 6 разів робить клімат украй посушливим.
У світі пустельні екосистеми займають 48,8 млн. км2, що становить 32% суходолу. Якщо до них ще приєднати близькі екосистеми напівпустель, то в сумі це складатиме 43% суходолу. Найбільші пустелі світу - це Сахара (7 млн. км2) та Лівійська пустеля в Африці (2 млн. км2), Гобі в Азії (1 млн. км2), Великий Басейн в Америці (1 млн. км2).
За характером ґрунту пустелі поділяються на піщані, глинисті, кам'янисті, солончакові та ін. Ґрунтовий покрив малопотужний. Гуміфікація йде дуже повільно через нестачу рослинного матеріалу та низьку вологість.
Рослинний покрив пустель сильно зріджений. Біопродукція низька; запаси біомаси вимірюються до 2,5 тонн/га сухої органічної речовини. Життєві форми рослинних організмів відрізняються своєрідністю, яка виробилася в результаті тривалої еволюції до найефективнішого отримання вологи та її економної витрати. Більшість рослин пустель дрібнолисткові, часто замість листя вони мають луски або колючки. На поверхні самих рослин розвивається потужний прошарок кутикули або кори. Листки в багатьох випадках сильно опушені. Кореневі системи типових для пустель рослин проникають у ґрунт на глибину 3—10 м. В окремих бобових чагарників корені сягають на глибину до 30 м.
Багатьом пустелям характерна весняна синузія ефемерів та ефемероїдів, весь цикл активного росту та розмноження яких пов'язаний з весняним - коротким, але найбільш вологим -періодом року.
Особливим елементом екосистем пустель є оази, що знаходяться в дельтах рік та біля інших джерел води. В оазах рослинність утворена головним чином культурними формами, оскільки вони здавна були основним місцем поселення людини в пустелях. Уздовж річок у долинах ростуть переважно тополі, тамариск, лох, обліпиха.
Домінуючі види рослин пустель залежать від типу ґрунту та географічного положення самих пустель. В азіатських пустелях деревно-чагарникові форми представлені саксаулами, джузганом та дроком. Трави порівняно нечисленні. Це різні види аристиди та полину. Пустелям американського континенту притаманні різновиди кактусів та агав, які запасають воду в стеблах та листках. Злаки представлені бізоновою травою. У місцях з трохи сприятливішим водним режимом пустелі переходять у напівпустелі. Зовні це проявляється в більш щільному злаковому рослинному покриві. Річна біопродукція тут вища.
Тваринний світ екосистем пустель бідний. Він так само, як і рослинність, сформувався унаслідок дефіциту вологи. Тільки під час весняних та осінніх перельотів тут вирує життя: з'являються зграї качок, гусей, журавлів та інших птахів.
Корінні види тварин пустель тісно пов'язані з ґрунтом, де вони знаходять вологу та захист від спеки. Це плазуни, гризуни, терміти та земляні комахи. Багато видів тварин виробили здатність зберігати вологу у вигляді жирових депо (горб у верблюда, підшкірний жировий прошарок у гризунів, жирове тіло в основі хвоста в тушканчиків тощо). Цей жир при біохімічному окисленні виділяє вільну воду, яка необхідна для організму. Деякі види тварин залишають пустелю на період найспекотливіших місяців, перебираючись на більш вологі та прохолодні території.
Активність тварин пустель проявляється в нічні прохолодні години. Удень тварини ховаються або в ґрунті, або, навпаки, піднімаються на верхівки гілок чагарників і дерев, де повітря більш прохолодне порівняно з піском, розпеченим сонцем. У норах гризунів велика кількість птахів влаштовує свої гнізда.
Основні фітофаги пустель - верблюди, гризуни та черепахи. Листя активно поїдається різними видами саранових. Невелика кількість видів пристосувалася до поїдання коренів (ризофаги). Хижаки представлені шакалами, гієнами, у пустелі Африки заходять леви.
Детритний трофічний ланцюг екосистем пустель представлений термітами, чорнишами та скарабеями. Схема трофічної мережі пустель наведена па. рис. 9. Тварини пустель завжди тяжіють до водойм. Внаслідок їх незначної кількості тварини змушені багато пити про запас. Вода, що отримується з великими труднощами, використовується бережливо: з метою запобігання втрат води тіла тварин мають товстий хітиновий (комахи) або роговий (плазуни) покрив. Але в будь-якому випадку випаровування необхідне для охолодження тіла. Тому в багатьох видів спостерігається швидке дихання, роти та дзьоби утримуються відкритими. Саме з цієї причини вуха в тушканчиків та зайців великі. Деякі види взагалі не п'ють воду, задовольняючись тією, яку отримують з їжею (рослинною чи тваринною). Існують дуже цікаві способи забезпечення молодняка водою. Так, деякі види птахів, зокрема рябки, які гніздяться далеко від водойм, прилітають на водопій та рясно змочують своє пір'я. Після цього швидко повертаються до гнізда, де їхні пташенята смокчуть вологе пір'я і таким чином отримують необхідну воду Чиста біопродуктивність у пустелях не перевищує 0,2 кг/см2 на рік, при цьому 75% біомаси зосереджено в ґрунті.
Сучасні екосистеми пустель несуть значний відбиток діяльності людини. Інтенсивний випас худоби та інші види господарської діяльності сильно порушили рослинний покрив та збіднили тваринний світ. Великих збитків завдає ерозія ґрунтів.
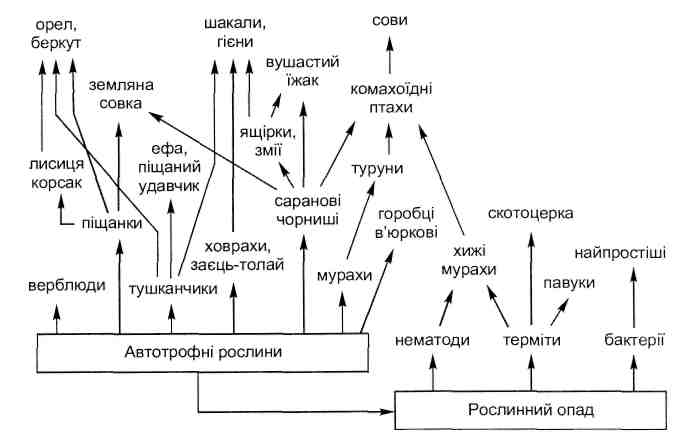
Рис. 9. Спрощена трофічна мережа екосистеми пустелі
ЕКОСИСТЕМИ ЛУКІВ. За думкою В.В. Альохіна (1925), луки належать до азональних природних екосистем. Це означає, що в їх формуванні провідну роль відіграє не клімат, а рельєф та ґрунт. Типові луки розвиваються в заплавах річок, і їх називають заплавними. Вигляд заплавних лук у багатьох випадках визначається характером річкових відкладів.
Заплавні луки розміщуються в різних кліматичних зонах. Уздовж течії північних річок вони сягають зон тайги та тундри, уздовж течії південних рік — степів та пустель. Залежно від положення в самій заплаві екосистеми луків можуть розміщуватися на дуже родючому або дуже бідному ґрунті. Відповідно такі біотопи можуть бути або сухими, або перезволоженими. Стосовно цього в заплавах розрізняють три основні частини: 1) прируслова частина з крупнопіщаних відкладів з низькою родючістю та швидким висиханням субстрату; 2) центральна частина заплави з родючими мулистими відкладами; 3) притерасна заплава з дрібномулистими відкладами, перезволожена або навіть заболочена.
Ґрунти луків дернові, лугові. У заплавах степових річок вони засолені. У таких випадках виникають солончакові луки з ячменем короткоостим, подорожником та ситником Жерара.
Неабияке значення для лугових екосистем має режим розливу рік. У річок, що мають витоки на рівнинах, повінь відбувається весною, а в ріках, що стікають з гір, - у середині літа, коли починається сходження снігу. На ріках України через масове вирубування лісів повені стали малими, а в деякі роки взагалі не спостерігаються
Рослинний покрив луків формується з багаторічних дводольних трав та злаків. У прирусловій заплаві ростуть довгокореневищні види (стоколос безостий, пирій, куничник наземний). У центральній частині заплави звичайно розвивається багатий травостій з дернових злаків, бобових та різнотрав'я. У притерасній заплаві переважають осоки, очерет та комиш. Згідно з Д.Я. Афанасьєвим (1965), на луках Дніпра травостій має такий склад: багаторічні трави - 69%, дворічні та багаторічні трави -1,3%, типові дворічні—6,7%, однорічні та дворічні види - 3,6%, однорічні трави - 19%. Залежно від кліматичної зони, в якій розташовані луки, до їх складу можуть входити тундрові, лісові або степові види.
Фауна луків не відрізняється специфічністю, їх у багатьох випадках складають окремі зональні фауністичні комплекси. На луках часто оселяються кроти, полівки.
Значна частина екосистем луків є результатом діяльності людини, яка вирубує ліс, сприяє формуванню луків. В Україні суходільні луки не займають великих площ. Усього в Україні налічується 6,8 млн. га природних луків. Лугові екосистеми відрізняються одна від одної залежно від зонального положення. На території України чітко виділяються луки заплав у лісовій зоні, луки заплав середньої течії Дніпра та Сіверського Дінця, луки заплав лісостепових річок, луки заплав плавнів Дніпра, Дунаю та Дністра.
За даними Ю.Р. Шеляг-Сосонка, в Україні налічується 54 формації природних луків. Існує 6 основних класів формацій луків:
Справжні мезофітні луки.
Остепнені ксеромезофітні луки.
Пустошні луки.
Болотяні луки з гідромезофітним рослинним покривом. Торф'янисті луки.
Галофітні луки
Разом з тим лукові екосистеми диференційовані від одної залежно від зонального положення.
Луки скрізь використовуються у сільському господарстві й дають у середньому 12-14 ц/га сіна або 6-8 ц/га пасовищного корму на рік. До складу флори лук України входить більш ніж 500 видів судинних рослин.
Заплавні луки здавна використовують як сіножаті і для випасу худоби, їх природний характер із цієї причини значною мірою втрачений.
БОЛОТА. Болотні екосистеми, як і лугові, є азональними. Вони виникають у місцях сильного перезволоження ґрунту. У таких випадках детритний трофічний ланцюг вкорочується та не завершується утворенням гумусу. Рослинний опад накопичується з року в рік у напіврозкладеному стані та утворює торф.
Загальна площа боліт Землі становить 350 млн. га. Торфова маса погано прогрівається, є бідною на мінеральні речовини, і тому рослинний покрив боліт у цілому досить убогий. Болотному ґрунтові характерна так звана фізіологічна сухість. При загальній високій вологості корені рослин ледь отримують з нього воду. Перешкодою є низька температура торфової маси та насиченість води гуміновими кислотами. Відповідно до основних характеристик боліт вони поділяються на три види: а) низинні; б) перехідні; в) верхові.
Низинні болота виникають у місцях виходу ґрунтових вод або на місці озер. Рослинний покрив таких боліт формується з осоки, очерету, рогозу та комишу. Вони і є основними торфоутворювачами. Часто такі болота мають розріджений деревостій з вільхи та верб. У низинних болотах мінералізація досить виражена, а болота такого типу визначаються як ефтотрофні.
Верхові болота утворюються головним чином на водотривких гірських породах з атмосферних опадів, але можуть також виникати й на місці низинних боліт. Основу рослинного покриву верхових боліт складають сфагнові мохи. Після відмирання формується торф, потужність залягання якого може сягати 5 м. Сфагновий торф погано
піддається гуміфікації та мінералізації, тому ґрунти таких боліт дуже бідні, а болота називаються оліго-трофними.
На сфагнових болотах може розміщатися розріджений сосновий деревостій. Ростуть тут також чагарники та чагарнички андромеда, касандра, баглиця та чорниця. Часто зустрічається журавлина. Видове розмаїття вкрай низьке. На 1 м2 тут налічується лише 2-5 видів рослин.
Перехідні болота є стадією переходу від низинних до верхових боліт. Часто вони розміщуються навколо верхових боліт. За вмістом поживних речовин вони також займають проміжне положення та називаються мезотрофними. У рослинному покриві переважає осока.
Болотні екосистеми небагаті на тварин. Найбільш поширені птахи.
У розміщенні боліт простежується загальна закономірність. У зоні лісотундри представлені головним чином бугристі болота. У тайговій зоні переважають аапа-болота, що являють собою грядово-мочарні комплекси з увігнутою поверхнею. У таких комплексах чергуються евтрофні, мезотрофні та оліготрофні гряди. Дерева тут не ростуть. На крайньому півдні тайгової зони з'являються опуклі грядово-мочарні болота. У лісостепу та степу болота евтрофні, осокові та очеретяні. На півдні степової зони та в пустелях розвиваються зволожені трав'яні болота.
В Україні болота можна спостерігати в усіх трьох природно-кліматичних зонах. В Українському Поліссі найбільш поширені оліготрофні сосново-сфагнові та евтрофні трав'янисті болота. У лісостепу України частіше зустрічаються евтрофні осокові та очеретяні болота. У степу їм на зміну приходять прісноводні чи засолені трав'янисті болота. У цілому в Україні можна виділити три основні райони поширення боліт:
а) поліські сфагнові болота;
б) поліські та лісостепові трав'янисто-гіпнові болота;
в) поліські лісові болота.
Усього болотних формацій в Україні налічується 53.
Запаси біомаси в болотних екосистемах вимірюються в межах 90-1770 ц/га. Болотні екосистеми відіграють у біосфері виняткову роль. Вони є накопичувачами прісної води та, займаючи всього 2% площі суходолу, утримують у зв'язаному вигляді (у формі торфу) 14% вуглецю. З боліт починається більшість річок. Особливо важлива роль боліт як своєрідних фільтрів або ж очисних систем, що затримують у шарі торфу різноманітні ксенобіотики та нітрати, які потрапляють туди разом зі стічними водами та атмосферними опадами.
На болотах збирають ягоди. Врожаї журавлини досягають 100-1000 кг/га. Після осушення боліт на них заготовляють сіно або випасають худобу.
ПРІСНОВОДНІ ЕКОСИСТЕМИ. Прісноводні екосистеми широко представлені на всіх континентах. Ріки та озера Землі вміщують 116 м3 води. Основна частина цієї води прісна, хоча деякі внутрішні водойми мають солону воду (це характерно для жаркого та сухого клімату).
У прісноводних озерах завжди виділяють три частини, які можна розглядати як окремі екосистеми. Це прибережна частина - літораль, глибоководна частина - профундаль та основна товща води - пелагіаль. Найбільш заселена живими організмами - літораль. Прибережні зони будь-яких водойм є їхніми головними трофічними областями. Окрім напівзануре-них рослин, у водоймах живуть придонні організми, які складають бентос, та планктон, що плаває у товщі води. Продукція більшості водойм часто лімітується нестачею біогенних мінеральних речовин. Річ у тому, що життя зосереджене у верхніх шарах води, де є достатньо сонячного світла, а мінеральні речовини надходять з придонних шарів. Верхні та нижні прошарки води розділені між собою так званим термоклином, що особливо чітко проявляється у водоймах субтропічного та тропічного поясів. Термоклин перешкоджає вертикальному водообміну та призводить до дефіциту мінеральних речовин у поверхневих шарах води.
Літораль характеризується наявністю великої кількості прикріплених рослин - макофітів. Фауна представлена комахами та їх личинками. З молюсків часто зустрічаються ставковики та плоскі котушки. На дні живуть жабурниці. Багата й фауна хижаків. Тут зустрічаються п'явки, водяні клопи, плавунці. Численні личинки бабок, одноденок, комарів.
У прибережній частині озер звичайними є такі види риб, як плітка, краснопірка, лин, дикий короп, колюшка. Хижі риби представлені щукою, окунем та судаком. Прибережна частина озер є місцем проживання жаб, тритонів, вужів та гадюк.
Придонна частина озер майже не має рослин, вода мало рухома і зберігає протягом майже всього року температуру +4°С. Фауна таких місць збіднена. Вона представлена в основному личинками комарів-дзвонців та молюсками.
У пелагіалі рослини представлені плантктоном із синьозе-лених, діатомових та зелених водоростей, макрофітами, що плавають (елодея, рдести). Усі живі органами мають різноманітні пристосування, що допомагають їм утримуватися в товщі води. У рослин це парашутоподібні вирости, крапельки жиру в тілі, тварини утримуються за допомогою активних рухів. У пелагіалі водяться озерна форель, сигові риби. Тут багато хижих коловерток, веслоногих рачків та циклопів. Рослинний та тваринний світ озер у багатьох випадках визначається наявністю у воді поживних речовин. За цією ознакою озера поділяються на евтрофні - багаті на азот, фосфор, оліготрофні - бідні на азот і фосфор (нітратів менше 1 мг/ л) та проміжні - мезотрофні. Фауна риб у цих трьох типах озер суттєво відрізняється. Для оліготрофних озер характерні сиги, гольці, окуні, щука та плітка. В евтрофних озерах живуть види, які стійкі до частого тут дефіциту кисню - короп, лин, карась, плітка та лящ.
У розвитку річкових екосистем основну роль відіграють характер дна та берегів, температура води та швидкість течії. У прибережній частині струмків та річок ростуть звичайні для цих місць очерети, комиші, лепешняки та стрілолист. У товщі води плавають елодея, латаття. При підвищенні швидкості течії до 0,3-0,6 м/сек та більше товща води вже не зростає. Для річок планктон не характерний, оскільки зноситься течією.
Річкова ентомофауна досить різноманітна. Тут чимало водяних комах та їхніх личинок. Часто зустрічаються рачки-бо-коплави. Уздовж течії рік спостерігається своя закономірність у розподілі іхтіофауни. У витоках чистих рік із прозорою водою живе форель, у середній течії основними видами є харіус та вусач, тут звичайні лини та головань. У нижній частині рік, де течія сповільнюється, до складу іхтіофауни входять лящ, короп, щука та верховодка.
Трофічні ланцюги прісноводних екосистем та особливо річок є короткими через відсутність багатої кормової бази. Вони починаються з автотрофних рослин і закінчуються в пасовищних трофічних ланцюгах хижими рибами, а в детритних трофічних ланцюгах - мікроорганізмами.
Прісна вода річкових екосистем дуже важлива для забезпечення потреб людей. На планеті вона в дефіциті. У наш час 1 млрд. населення страждає від гострої нестачі прісної питної води, а 1,7 млрд. користується водою низької якості.
На території України зареєстровано 71 тис. річок, що мають загальну довжину 243 тис. км. Більшість рік належить до басейнів Чорного та Азовського морів. В Україні три тисячі озер із загальною площею водного дзеркала 2 тис. км2. Крім цього, країна має 23 тис. ставків та водосховищ, особливо їх багато в районі середнього та нижнього Дніпра Ріки та озера України вміщують у собі 195 видів водяних макрофітів (Дубина та ін., 1993), а також багато тисяч видів водоростей. В Україні є 57 водних рослинних формацій. Водні екосистеми є важливим національним багатством. Це й сховища прісної води, і джерела різноманітної продукції, і місця відпочинку населення. Для збереження запасів прісної води України Національний екологічний центр України провів громадську акцію з охорони малих рік «Жива вода - 1996». Ці ріки складають 60% водяного ресурсу держави.
ОКЕАНІЧНІ Й МОРСЬКІ ЕКОСИСТЕМИ. Океанічні й морські екосистеми світу найбільш протяжні, їхня сукупна площа становить більше 70% поверхні земної кулі. Загальний обсяг води у Світовому океані досягає 138 000 тис. км3. Це приблизно 96% загальної кількості води на планеті. Важливою особливістю цих екосистем є цілісність - усі океани й моря з'єднуються між собою, за винятком кількох ізольованих засолених водойм (Каспійське й Аральське моря), які явно є реліктовими залишками й колись були з'єднані зі Світовим океаном.
Глибина океанів і морів сильно варіює, складаючи в середньому близько 3 700 м. На окремих ділянках можуть спостерігатися більші чи менші глибини. Так, глибина Маріанської западини в Тихому океані досягає більше 7 000 м, а в найглибшій її частині - 11 022 м, Азовське море має глибину всього 14 м.
Залежно від топографічного положення й глибини в будь-якому океані чи морі розрізняють кілька частин:
а) літораль - прибережна частина, яка осушується під час відливів і заливається під час припливів;
б) континентальний шельф - зона, що йде вздовж берегів із глибинами в 200-400 м;
в) пелагіаль - основна товща води в океані чи в морі;
г) абісаль - придонна частина води.
Для Світового океану характерна солона вода, обумовлена наявністю катіонів натрію, магнію, кальцію й калію та аніонів -хлоридів, сульфатів і бікарбонатів. Середня солоність води - близько 35 г/л.
Поширення фотосинтезуючих рослин в океанах і морях обумовлене глибиною проникнення сонячної радіації та охоплює приблизно 50 м, а там, де вода особливо прозора, - верхні 150-200м.
При збагаченні цього шару води мінеральними речовинами, що спостерігається на мілководдях у дельтах рік, естуаріях, лиманах і на коралових рифах, біологічна продукція буває дуже високою. Досить висока вона й у зоні континентального шельфу, до якого звичайно прив'язані промисли риби та інших морських тварин (оселедець, тріска, хек, краби, лангусти й ін.). В інших частинах пелагіалю біопродукція автотрофних рослин і відповідно чисельність тварин багато нижча.
Розмаїтість видів і біологічна продукція у Світовому океані має чітку зональність, обумовлену температурою. У тропічних і субтропічних широтах вони найбільші. Але фітопланктон найбідніший у тропічних широтах, і його кількість збільшується в напрямку до півдня й півночі від екватора. Це пов'язано з характером циркуляції води: теплі води (у приекваторіальних широтах їхня температура складає 26°-27°С) знаходяться біля поверхні, а більш важкі холодні (їхня температура не вище 0°-4°С) -на глибині, тому вертикальне підняття мінеральних речовин із дна морських екосистем до поверхні, де тільки й можуть жити автотрофні зелені організми, сильно ослаблене.
Винятком є так звані зони апвелінгу, де під впливом океанічних течій глибинні холодні води, багаті мінеральними речовинами, піднімаються на поверхню. У зонах апвелінгу біологічна продукція дуже велика. Тут рясно розростаються різні водорості (у першу чергу діатомові), що створює чудові умови для харчування багатьох видів тварин. Основні місця промислу оселедця, анчоусів та інших знаходяться саме в зонах апвелінгу.
У Світовому океані живуть найдревніші види тварин і рослин, що підтверджує думку про нього як про «колиску життя» нашої планети. Але кількість видів живих організмів тут менша, ніж на суші, тому що в більш однорідному водному середовищі еволюція відбувалася значно повільніше.
Мережі живлення в океанах і морях досить складні. їх основу складають автотрофні рослини. Вони представлені у Світовому океані різними групами водоростей (зелені, діатомові, бурі, червоні). У товщі води живуть плаваючі водорості, що складають фітопланктон, а біля дна - прикріплені (фітобентос). Консументи першого порядку у Світовому океані представлені головним чином зоопланктоном. Це найпростіші, дрібні ракоподібні, медузи, сальпи. Великі морські тварини мають або змішаний тип харчування, споживаючи фіто- і зоопланктон, як, наприклад, кити, або є консументами другого порядку - хижаками, як акули. До 90% біомаси тваринних океанів складають безхребетні тварини - молюски, омари, креветки, медузи й ін.
Редуценти представлені в океанічних екосистемах бактеріями, що живуть на всіх глибинах. У прибережних частинах їх чисельність досягає 0,5-3 млн. бактерій на 1 л води, у пелагіалі їх від 5 до 20 тис. на 1 л. Тому в пелагіалі частина фітомаси продуцентів і консументів не мінерализується, опускається на дно океанів і таким чином вилучається з подальшого кругообігу речовин. Орієнтовні підрахунки, виконані П. Дювиньо і М. Тангом (1968), такі:
на частку фітопланктону в океанах припадає 27 млрд. т органічної речовини;
безхребетні тварини, у тому числі молюски, формують З млн. т біомаси;
риби мають біомасу в ЗО млн. т, з яких оселедця - 3,3 млн. т, тріски - 2,5 млн. т;
кити складають в океанах близько 1 млн. т біомаси, цих великих тварин (вага одного синього кита дорівнює вазі 25 слонів) через хижацький промисел залишилося приблизно 40 тис. особин.
Для біологічної продуктивності морів важлива чистота морської води. В останні десятиліття моря й океани зазнають усе більшого забруднення різними токсикантами, що пов'язано з діяльністю людини. Особливо великі забруднення створює промисел і транспортування нафтопродуктів і газу, а також континентальні забруднені стоки. Незважаючи на величезну продуктивність океанів і морів планети, їх ресурси використовуються людиною поки на рівні 1-5%. Україна є співвласником Чорного й Азовського морів. Це замкнені водяні системи, істотно віддалені від Світового океану, зі специфічним екологічним режимом. Солоність води в Чорному морі - близько 19 г/л. Унаслідок погіршеного водообміну в Чорному морі з глибини 150-300 м товща води сильно збагачена сірководнем і будь-яке життя тут відсутнє. Серйозною проблемою Азово-Чорноморського басейну є зниження річного стоку і скидання в нього великої кількості промислових, сільськогосподарських та побутових відходів.
Лекция 15 – Определение, структура, границы и эволюция биосферы
Термин «биосфера» в научной литературе появился в 1875 г. в одной из публикаций австрийского геолога Э. Зюсса.
Учение о биосфере разработал в своих трудах В.И. Вернадский. Биосферой он называл оболочку Земли, в формировании которой живые организмы играли и играют основную роль. Ученый выделял в биосфере три главных компонента: живые организмы, или так называемое живое вещество, минеральные компоненты, включенные живым веществом в биогенный круговорот, и продукты деятельности живого вещества, временно не участвующие в биогенном круговороте. По В.И. Вернадскому, биосфера включает все части земной коры, на которые воздействовали живые организмы в течение всей геологической истории.
Биосфера — сложная многокомпонентная система, включающая всю живую и неживую природу. Она охватывает часть атмосферы, гидросферу и верхнюю часть литосферы, взаимосвязанные биогеохимическими циклами миграции веществ и энергии. Границы биосферы составляют: в атмосфере — до озонового слоя (25-30 км), в гидросфере — до максимальных глубин (около 11 км), в литосфере — до 8-10 м, реже до 3 км (нефтеносные слои воды). В состав биосферы входят живые организмы (около 3 млн. видов), их остатки, биогенное вещество (продукты жизнедеятельности живых организмов — осадочные породы органического происхождения), биокосное вещество (продукты распада и переработки горных и осадочных пород живыми организмами) и косное вещество (горные породы магматического и неорганического происхождения, а также переработанные и видоизмененные живыми организмами вещества космического происхождения).
В основе учения В.И. Вернадского о биосфере лежат представления о планетарной геохимической роли живого вещества в образовании биосферы как продукта превращения веществ и энергии в ходе геологического развития Земли. В пределах биосферы встречается либо само живое вещество, либо следы его деятельности — газы атмосферы, природная вода, нефть, известняк, сланцы и др.
Живое вещество, по В.И. Вернадскому, — это совокупность существующих или существовавших живых организмов, являющихся мощным геологическим фактором. В отличие от живых организмов,
изучаемых на всех уровнях их организации, живое вещество как биогеохимический фактор характеризуется элементарным химическим составом, массой и энергией; оно трансформирует солнечную энергию и вовлекает неорганическую материю в непрерывный круговорот. Через живое вещество многократно прошли атомы почти всех химических элементов. В конечном счете, живое вещество определило состав атмосферы, почв и осадочных пород планеты.
По мнению В.И. Вернадского, живое вещество аккумулирует энергию космоса и трансформирует ее в энергию земных процессов (химическую, механическую, тепловую, электрическую и т.д.), обеспечивает образование нового живого вещества, которое не только замещает отмирающие его массы, но и привносит новые качества, определяя тем самым эволюцию органического мира.
Живое вещество биосферы химически и биологически активно. При его участии образуются органические осадочные породы и биокосные вещества: вода, почва, кора выветривания; оно контролирует все основные химические превращения в биосфере.
Различают пять главных функций живого вещества на планете — энергетическую, газовую, концентрационную, окислительно-восстановительную и деструкционную.
Энергетическая функция заключается в осуществлении связи биосферно-планетарных явлений с космическим излучением, преимущественно с солнечной радиацией. В ее основе лежит фотосинтетическая деятельность зеленых растений, благодаря которой происходит аккумуляция солнечной энергии и ее перераспределение между компонентами биосферы. За счет энергии Солнца, накопленной зелеными растениями, протекают все жизненные явления на Земле.
Газовая функция обеспечивает газовый состав биосферы, обусловливает миграцию и превращение газов. В процессе жизнедеятельности живого вещества образуются азот, кислород, углекислый газ, метан и другие газы.
Концентрационная функция проявляется в извлечении живыми организмами биогенных элементов из окружающей среды и их накоплении, так как состав живого вещества значительно отличается от среднего состава планеты. В нем преобладают легкие атомы водорода, кислорода, углерода, азота, кремния, серы, хлора, калия, кальция. Концентрация этих элементов в организмах намного выше, чем во внешней среде.
Окислительно-восстановительная функция заключается в химическом превращении атомов, главным образом с переменной валентностью (соединения железа, алюминия, марганца), что приводит к пре вращению большинства химических соединений. На поверхности Земли преобладают биогенные процессы окисления и восстановления.
Деструкционная функция обусловливает процессы, связанные с разложением организмов после их смерти. Происходит минерализация органического вещества или превращение живого вещества в косное, что приводит к образованию биогенного и биокосного вещества биосферы.
В.И. Вернадский (1967) писал: «Живые организмы являются функцией биосферы и теснейшим образом материально и энергетически с ней связаны, являются огромной геологической силой, ее определяющей. Живое вещество представлено совокупностью живых организмов, существующих в данный момент времени, и численно выражено в элементарном химическом составе, в весе, энергии. Оно связано с окружающей средой биогенным током атомов: своим дыханием, питанием и размножением». Главная особенность биосферы, по его мнению, — это биогенная миграция атомов химических элементов, вызываемая лучистой энергией Солнца и проявляющаяся в процессе обмена веществ, роста и размножения организмов.
Таким образом, по современным представлениям, биосфера — это своеобразная оболочка Земли, содержащая всю совокупность живых организмов и ту часть вещества планеты, которая находится в непрерывном обмене с ними.
Возникновение и эволюция биосферы. Возникновение биосферы теснейшим образом связано с появлением жизни на Земле. Некоторые исследователи связывают появление жизни на нашей планете со временем ее остывания и оценивают возраст Земли по этому показателю в 24 млн. лет. Эти вычисления в 1861 г. сделал английский физик В. Томсон. Значительно позже, после открытия явления радиоактивности и разработки на ее основе метода измерения геологического времени, оказалось, что расчеты В. Томсона ошибочны. По современным представлениям, возраст Земли оценивается в 4,55 млрд. лет, а сохранившиеся древнейшие участки земной коры — в 4 млрд. пет.
Авторы большинства гипотез о происхождении жизни на Земле допускали, что наша планета в течение огромного промежутка времени была безжизненной и на ее поверхности происходила химическая эволюция, которая предшествовала биологической. На поверхности планеты, по их мнению, происходил медленный абиогенный синтез органических соединений, который в конечном счете привел к появлению примитивных форм жизни. Однако имеются и другие представления, согласно которым за такой период невозможно образование столь огромного количества видов живых организмов, то есть продолжительность существования жизни на Земле не укладывается в такие временные рамки.
Анализ новых данных космохимии свидетельствует о раннем зарождении жизни в пределах Солнечной системы. Химическая эволюция вещества Земли и других планет, вероятно, предшествовала их образованию. Первичная атмосфера нашей планеты в основном была представлена углекислым газом. Но это химическое соединение не может самопроизвольно образовывать более сложные органические соединения. В настоящие время существует много подтверждений в пользу того, что зарождение жизни, возможно, произошло в космических условиях.
Анализ вещественного состава метеоритов показывает, что содержащееся в них органическое вещество имеет достаточно сложный состав. Впервые органическое вещество в метеоритах обнаружил шведский химик И. Берцелиус в 1834 г. при анализе углистого хондритового метеорита Алаис.
В органическом веществе метеоритов были обнаружены углеводороды, среди которых наиболее распространены соединения с 16 атомами углерода в молекуле, а также спирты, карбониловые соединения, аминокислоты и др. Следует отметить, что характерной особенностью органических соединений, обнаруженных в метеоритах, является отсутствие оптической плотности. Это свидетельствует об их происхождении за пределами Земли.
Теоретические и экспериментальные данные, полученные в последнее время, позволяют сделать вывод: синтез относительно сложных органических соединений, предшествующих появлению живого вещества, — закономерный этап химической эволюции Солнечной системы. Эти органические вещества, образовавшиеся в космических условиях, вошли в состав многих тел, но на Земле реализовались возможности дальнейшей эволюции, что обеспечило возникновение саморегулирующихся высокомолекулярных систем — непосредственных предков первых живых организмов.
Рассматриваются два варианта событий. Либо химическая эволюция, начавшаяся в космических условиях, продолжила свое развитие на Земле и в относительно короткое время привела к появлению
первых живых организмов, либо образование молекул ДНК произошло в космических условиях, а реализация ее возможностей — в первых водоемах планеты, которые содержали некоторое количество органического вещества.
Дальше в ходе геологической истории эволюция биосферы происходила по пути разрешения противоречия между безграничной способностью организмов к размножению и ограниченностью ресурсов, доступных в определенную геологическую эпоху. Данное противоречие разрешается путем овладения организмами новыми источниками вещества и энергии за счет приобретения ими новых качеств. В этом случае наследственная изменчивость является предпосылкой развития, а естественный отбор служит механизмом закрепления новых качеств.
Переломным этапом в эволюции древней биосферы был переход от гетеротрофного режима питания к автотрофному, основанному на фотосинтезе. С появлением фотосинтезирующих организмов началось образование свободного кислорода, что со временем стало предпосылкой для создания в атмосфере озонового экрана. Это произошло около 4 млрд лет тому назад. Увеличение содержания кислорода в атмосфере способствовало выходу органического мира на поверхность континентов. О времени появления живых организмов на суше точных палеонтологических данных нет.
Переход от прокариотной флоры к эукариотной в древних морях происходил медленно, и обе группы длительное время существовали совместно. Соотношение между этими группами организмов постоянно изменялось и к настоящему времени оно сложилось в пользу эукариотов. Эволюция растений длительное время происходила в водной среде. Далее по мере накопления кислорода создались предпосылки для появления озонового экрана, который защищает все живое от ультрафиолетового излучения. Это в конечном счете создало условия для выхода растений из водной среды на континенты. Считается, что первыми растениями, появившимся на суше, были псилофиты — споровые низкорослые растения, напоминающие плауны. Потом псилофиты уступили место папоротникообразным растениям, которые в свою очередь сменились хвойными.
Развитие растений, видимо, создало предпосылки для появления животных. Они так же, как и растения, произошли от одноклеточных организмов. В результате дифференциации функций отдельных клеток на определенном этапе эволюции образовались организмы, давшие затем начало многоклеточным.
На основании палеонтологических данных в ходе эволюции органического мира выделяют ряд закономерностей:
 необратимость
эволюции. Это
положение в 1893 г. сформулировал
бельгийский палеонтолог Л. Далло:
организм не может вернуться
хотя бы частично к предшествующему
состоянию, которое было в ряду его
предков;
необратимость
эволюции. Это
положение в 1893 г. сформулировал
бельгийский палеонтолог Л. Далло:
организм не может вернуться
хотя бы частично к предшествующему
состоянию, которое было в ряду его
предков;ускорение биологической эволюции в ходе геологического времени;
закономерность, впервые отмеченная В.О. Ковалевским в 1871 г.: «..каждая следующая большая эпоха Земли короче предыдущей, и в это короткое время успевало народиться и вымереть больше разнообразных форм, чем в предыдущую эпоху...» (цит. по Г.В. Войткевичу, В.А. Вронскому, 1996);
эволюция различных групп организмов протекала с разной скоростью;
существуют консервативные группы организмов, которые почти не изменились в ходе геологического времени (микроорганизмы, папоротниковые, плауны, голосеменные), однако они составляют небольшую часть из общего числа видов;
—- на фоне общей тенденции ускорения эволюции определенные эпохи отличались повышенным видообразованием. Вероятно, это было связано с радиоактивностью. С.Г. Неручев (1983) в геологической истории Земли выделил 30 эпох уранонакопления. Эти эпохи отличаются значительным усилением мутационных процессов, видообразования и сменой фауны и флоры;
— среди животных в ходе геологического времени происходит направленное изменение нервной системы.
На основании данных палеонтологии, геохимии и космохимии эволюцию биосферы Земли можно представить в виде трех последовательно сменяющихся этапов.
Первый этап — восстановительный. Он начался в космических условиях и завершился появлением на Земле первой гетеротрофной биосферы. На этом этапе протекали каталитические и радиохимические реакции синтеза сложных органических соединений, отсутствовал свободный кислород, основным источником живых организмов была радиация. Этот период, вероятно, был коротким по времени.
Второй этап — слабоокислительный — характеризовался появлением фотосинтеза. Он длился более 2 млрд. лет и закончился около 1,8 млрд. лет тому назад. Свободного кислорода образовывалось еще мало, и атмосфера состояла преимущественно из углекислого газа. Развитие организмов ограничивалось прокариотами.
Третий этап — окислительный — связан с появлением фотоавтотрофной биосферы. Он начался с медленного роста содержания кислорода в атмосфере и завершился значительным ускорением эволюции организмов. Увеличение продукции кислорода привело к появлению растительного покрова и животных на континентах, что резко увеличило продукцию фотосинтеза. Под воздействием живого вещества сформировался современный химический состав атмосферы и растворенного вещества гидросферы.
Биологическая эволюция, будучи необратимым процессом, предопределила необратимость эволюции биосферы в целом и создала предпосылки для ее перехода в качественно новое состояние — ноосферу, или сферу разума, когда все происходящие в биосфере изменения контролируются человеком.
Современная концепция биосферы. Биосфера как целое образует единую экосистему Земли. Она представляет собой особую геооболочку, включающую все живые организмы, взаимосвязанные с естественной средой обитания. Биосфера, возраст которой составляет не менее -3,5 млрд. лет, является результатом развития жизни как планетарного явления.
Термин "биосфера" (от греческого bios - жизнь и sphaira - шар) был предложен, по- видимому, Ж.-Б. Ламарком в начале XIX столетия для обозначения совокупности живых существ, населяющих Землю. Позднее, в 1875 году, австрийский геолог Э.Зюсс использовал этот термин в расширительном смысле, обозначив им поверхностную геооболочку ("лик Земли").
Дальнейшую глубокую и детальную проработку понятие биосферы получило только в первой половине XX века в трудах выдающегося русского геохимика академика В. И. Вернадского (1863-1945).
По Вернадскому, биосфера - сложно организованная саморегулирующаяся и саморазвивающаяся система, представляющая собой закономерное космическое явление; "она теснейшим образом связана со строением земной коры, входит в ее механизм и в этом механизме исполняет величайшей важности функции, без которых он не мог бы существовать". Эти функции биосферы, по мнению В.И.Вернадского, носят энергетический характер. Он считал, что" по существу биосфера может быть рассматриваема как область Земной коры, занятая трансформаторами, переводящими космические излучения в действенную земную энергию - электрическую, химическую, механическую, тепловую и т.д."
Вследствие этого, "вещество биосферы проникнуто энергией; оно становится активным, собирает и распределяет в биосфере полученную в форме излучений энергию, превращает ее в конце концов в энергию в земной среде свободную, способную производить работу". Бесконечный поток энергии Солнца вызывает в биосфере закономерный круговорот вещества, в результате чего "между ее косной безжизненной частью, ее косными природными телами и живыми веществами, ее населяющими, идет непрерывный материальный и энергетический обмен, материально выражающийся в движении атомов, вызванном живым веществом". Отсюда следует, что биосфера "в такой же, если не в большей степени, есть создание Солнца, как и выявление процессов Земли".
Тесная внутренняя взаимосвязь и взаимозависимость живых и неживых компонентов (живого и косного вещества, по Вернадскому) находит свое отражение я "резко обособленных размерах" биосферы, призванных обеспечивать оптимальное взаимодействие земной поверхности с космическим, главным образом солнечным, излучением. "Пределы биосферы обусловлены, прежде всего, полем существования жизни. Жизнь может проявляться только в определенной среде, в определенных физических и химических условиях. Это как раз та среда, которая отвечает биосфере". Понимаемая таким образом биосфера захватывает верхнюю часть литосферы (земной коры), всю гидросферу и нижние слои атмосферы. При этом верхней границей ей служит озоновый слой на высоте 20-25 км, выше которого ультрафиолетовая часть солнечного спектра исключает существование незащищенных живых организмов. Нижняя граница биосферы опускается на 2-3 км под материками и на 1-2 км ниже дна океана.
Живые организмы Земли распределены в основном в верхнем слое литосферы и гидросферы. Организмов, проводящих всю свою жизнь в воздухе, то есть связанных с атмосферой как с основной средой обитания, по-видимому, нет.
'Толщина "пленки жизни" обусловлена глубиной проникновения в биосферу солнечной радиации: чем лучше та или иная геооболочка пропускает солнечные лучи, тем глубже в нее проникает жизнь, а благодаря гравитации поток энергии в форме органического вещества распространяется в глубины Мирового океана.
Вторым источником энергии (хотя незначительно более слабым) для биосферы являются недра Земли. Открытие в рифтовых зонах Мирового океана экосистем на основе хемосинтеза - "оазисов жизни" - значительно расширило наши представления об адаптативных возможностях живых организмов.
Гидросфера заполнена жизнью по всей своей толще, хотя наибольшая плотность жизни наблюдается в поверхностном слое вод, прозрачном для солнечных лучей (эвфотической зоне).
В литосфере биомасса локализована в верхних горизонтах почвы мощностью от нескольких десятков сантиметров до нескольких метров. По трещинам земной коры, шахтам, скважинам живые организмы могут проникать значительно дальше, достигая глубин в 2,5-3 км. Так, например, установлено, что нефтяные воды, сопровождающие залежи нефти, глубоко, под землей заселены нефтсокисляющими бактериями.
Естественным барьером на пути распространения живых организмы в вглубь является высокая температура: на суше на глубине 1 км она составляет 20-40 °С, на глубине 3 км - 40-60 "С, 4 км - 70-90 °С, 5км -100-120 °С.
Проникновение живых организмов на суше ввысь ограничивается в основном недостатком жидкой воды и разреженностью атмосферного воздуха. Поэтому в горах зеленые растения не поднимаются выше 6200 м. Тем не менее, на больших высотах встречаются насекомые и микроорганизмы.
В соответствии со взглядами В.И.Вернадского, вещество биосферы подразделяют на несколько геологически взаимосвязанных типов, важнейшими из которых являются:
живое вещество - совокупность всех живых организмов Земли;
биогенное вещество - органические и органоминеральные продукты жизнедеятельности организмов (например, каменный уголь, нефть, известняки);
биокосное вещество - вещество, созданное одновременно живыми организмами и косными процессами (например, почва, океаническая вода );
косное (неживое) вещество - горные породы неорганического происхождения.
В пределах биосферы всюду присутствует либо живое вещество, либо следы его биохимической деятельности в форме биогенного и биокостного вещества.
По современным оценкам (A.M.Рябчиков, 1980), масса биосферы, включая все органическое вещество геосфер, занятых биосферой, составляет (2,5-3)* 1018 т, в том числе масса живого вещества Земли приблизительно 2*1012 т, масса компонентов биогенного вещества - каустобиолитов (нефти, каменного угля и др.) и известняков, соответственно, 3*10 и 5*10 т, масса биокосного вещества, заключенного в почвах, 2*1012 т (в пересчете на гумус).
В состав биосферы входят:
Геобиосфера — верхняя часть литосферы, населенная геобионтами; гидробиосфера — гидросфера без подземных вод, населенная гидробионтами; аэробиосфера — нижняя часть атмосферы, населенная аэробионтами.
Геобиосфера состоит из террабиосферы (с террабионтами) — поверхность суши, и литобиосферы (с литобионтами) — глубокие слои земной коры. Террабиосфера разделяется на фитосферу — пространство от поверхности земли до верхушек деревьев (0—150 м), и педосферу (с педобионтами) — почвенный покров (до 2—3 м), нередко сюда включают всю кору выветривания. Литобиосфера (до 2—3 максимум до 6 км) включает гипотеррабиосферу (подтеррабиосферу) — слой, где возможна жизнь аэробов (до 1 — 1,5 км), ее нижняя граница совпадают с нижней границей подземной тропосферы (почвенным и подпочвенным воздухом) и теллуробиосферу (глубинобиосферу) — слой, где возможно обитание анаэробов (до 2—3, максимум до 6 км). Живые организмы в толще литобиосферы обитают в основном в порах горных пород, заполненных подземными водами.
Гидробиосфера включает маринобиосферу или океанобиосферу (с маринобионтами) — моря и океаны и аквабиосферу (с аквабионтами) — континентальные, главным образом, пресные воды, которая в свою очередь разделяется на лиманоаквабиосферу — стоячие континентальные воды и реоаквабиосферу — проточные континентальные воды. Кроме того, гидробиосфера делится на слои связанные, главным образом, с интенсивностью света: фото(био)сферу — относительно ярко освещенный слой (до 150—200 м), дисфото (био)сфе-ру — всегда сумеречный слой — проникает до 1 % солнечной инсоляции (от 200 м до 1,5—2 км), афото(био)сферу — слой абсолютной темноты, где невозможен фотосинтез (глубже 1,5—2 км).
Аэробиосфера состоит из тропобиосферы (с тропобионтами) — слой от вершин деревьев до высоты наиболее частого расположения кучевых облаков (до 5—6 км), постоянно населенный живыми организмами,более тонкий,чем атмосферная тропосфера, и стратобиосферы (со стратобионтами), или альтобиосферы (с альтобионтами), — слой (от 5—6 до 6—7 км), где могут постоянно существовать микроорганизмы, главным образом в виде спор.
Лимитирующим фактором развития жизни в аэробиосфере служит наличие капель воды и положительных температур, а также твердых аэрозолей, поднимающихся с поверхности земли. На больших высотах в горах (около 6 км) расположена высотная часть террабиосферы — эоловая зона. Здесь уже невозможна жизнь высших растений и вообще организмов-продуцентов, но ветры приносят сюда с более низких вертикальных поясов органическое вещество и при отрицательных температурах воздуха еще достаточно тепла от прямой солнечной инсоляции для существования жизни. Это царство членистоногих и некоторых микроорганизмов — эолобионтов. Еще одним лимитирующим фактором проникновения жизни вверх является жесткое космическое излучение. На высоте 22—24 км от поверхности Земли наблюдается максимальная концентрация озона — озоновый экран. Озон образуется из кислорода воздуха под действием солнечной радиации (02 —» 03). Озоновый экран отражает губительные для живых организмов космические излучения и частично ультрафиолетовые лучи.
Выше аэробиосферы расположена парабиосфера — слой (между 6—7 и 60—80 км), куда жизнь проникает лишь случайно и не часто, где организмы могут временно существовать, но не могут нормально жить и размножаться. Еще выше расположена апобиосфера, или «надбиосфера» (выше 60—80 км), куда никогда даже случайно не поднимаются живые организмы, но в незначительном количестве заносятся биогенные вещества (ее верхняя граница трудноуловима).
Жизнь в океанах достигает их дна. Живые организмы встречаются даже на глубине более 11 км, где температура воды около 200°С, но из-за высокого давления вода не кипит. Ниже, в базальтах, жизнь едва ли возможна.
Проникновение жизни в глубь литосферы ограничено высокими температурами земных недр и наличием жидкой влаги. В глубинах литосферы есть два теоретических предела распространения жизни — изотерма 100°С, ниже которой при нормальном атмосферном давлении вода кипит, а белки свертываются, и изотерма 460°С, где при любом давлении вода превращается в пар и жизнь принципиально невозможна (глубина 25 км). Перегретая жидкая вода обнаружена в литосфере до глубин 10,5 км. Нижняя граница жизни по литосфере фактически не опускается глубже 3—4, максимум 6—7 км на суше, и не более 1—2 км ниже дна океана.
Ниже геобиосферы расположена гипобиосфера («подбиосфера» — аналог парабиосферы в атмосфере) — слой, куда жизнь проникает лишь случайно и может здесь временно существовать, но не жить и размножаться. Еще ниже залегает метабиосфера — слой биогенных (преобразованных жизнью) пород, в котором ныне живые организмы не присутствуют (до 10—15 км). Образно выражаясь, это «следы былых биосфер». В ее нижней части процессы метаморфизма горных пород стирают признаки жизни. Под метабиосферой расположена абиосфера («небиосфера») — слои литосферы, не испытывающие сейчас и никогда ранее не подвергавшиеся влиянию живых организмов (глубже 10—15 км).
Кроме того, разделяют такие понятия как эубиосфера, метабиосфера и панбиосфера. Эубиосфера (собственно биосфера) — слой между верхней границей гипобиосферы и нижней границей парабиосферы. Это область наиболее активной современной жизни. Мегабиосфера — эубиосфера вместе с гипобиосферой и метабиосферой вглубь, и до озонового экрана вверх, т.е. область нынешнего и прошлого воздействия жизни на природу Земли. Панбиосфера — мегабиосфера с артебиосферой (пространством человеческой экспансии в околоземной Космос). Это оболочка Земли, преобразованная нынешней и прошлой жизнью и человеческой деятельностью. Вертикальная мощность эубиосферы в океанической области Земли достигает более 17 км, в сухопутной — 12 км. Мощность мегабиосферы — 33-35 км.
В.И. Вернадский рассматривал биосферу как область жизни, включающую наряду с организмами и среду их обитания. Он выделил в биосфере семь разных, но геологически взаимосвязанных типов веществ.
Живое вещество — живые организмы, населяющие нашу планету.
Косное вещество — неживые тела, образующиеся в результате процессов, не связанных с деятельностью живых организмов (породы магматического и метаморфического происхождения, некоторые осадочные породы).
Биогенное вещество — неживые тела, образующиеся в результате жизнедеятельности живых организмов (некоторые осадочные породы: известняки, мел и др., а также нефть, газ, каменный уголь, кислород атмосферы и др.).
Биокосное вещество — биокосные тела, представляющие собой результат совместной деятельности живых организмов и геологических процессов (почвы, илы, кора выветривания и др.).
Радиоактивное вещество — атомы радиоактивных элементов (например, уран (238U, 235U), торий (232Th), радий (226Ra) и радон (222Rn, 220Rn), калий (40К), рубидий (87Rb), кальций (Са), цирконий (96Zr), тритий (3Н), бериллий (7Ве, 10Ве) и углерод (14С) и др.
Рассеянные атомы — отдельные атомы элементов, встречающиеся в природе в рассеянном состоянии (в таком состоянии часто существуют атомы микро- и ультромикроэлементов: Mn, Co, Zn, Си, Аи, Нд и др.)
Вещество космического происхождения — вещество, поступающее на поверхность Земли из космоса (метеориты, космическая пыль).
Живое вещество биосферы обладает уникальными особенностями, обусловливающими его крайне высокую преобразующую деятельность (по Н.А. Воронкову, 1997).
Способность быстро занимать (осваивать) все свободное пространство. В.И. Вернадский назвал это всюдностью жизни. Данное свойство дало основание В.И. Вернадскому сделать вывод, что для определенных геологических периодов количество живого вещества было примерно постоянным (константой). Способность быстрого освоения пространства связана как с интенсивным размножением (некоторые простейшие формы организмов могли бы освоить весь земной шар за несколько часов или дней, если бы не было факторов, сдерживающих их потенциальные возможности размножения), так и со способностью организмов интенсивно увеличивать поверхность своего тела или образуемых ими сообществ. Например, площадь листьев растений, произрастающих на 1 га, составляет 8—10 га и более. То же относится к корневым системам.
Движение не только пассивное, но и активное, т.е. не только под действием силы тяжести, гравитационных сил и т.п., но и против течения воды, силы тяжести, движения воздушных потоков и т.п.
3 Устойчивость при жизни и быстрое разложение после смерти (включение в круговороты веществ). Благодаря саморегуляции живые организмы способны поддерживать постоянный химический состав и условия внутренней среды, несмотря на значительные изменения условий внешней среды. После смерти эта способность утрачивается, а органические остатки очень быстро разрушаются. Образовавшиеся органические и неорганические вещества включаются в круговороты.
Высокая приспособительная способность (адаптация) к различным условиям и в связи с этим освоение не только всех сред жизни (водной, наземно-воздушной, почвенной, организменной), но и крайне трудных по физико-химическим параметрам условий. Например, некоторые организмы переносят температуры, близкие к значениям абсолютного нуля —273°С, микроорганизмы встречаются в термальных источниках с температурами до 140°С, в водах атомных реакторов, в бескислородной среде, в ледовых панцирях и т.п.
Феноменально высокая скорость протекания реакций. Она на несколько порядков значительнее, чем в неживом веществе. Об этом свойстве можно судить по скорости переработки вещества организмами в процессе жизнедеятельности. Например, гусеницы некоторых насекомых потребляют за день количество пищи, которое в 100— 200 раз больше веса их тела. Дождевые черви (масса их тел примерно в 10 раз больше биомассы всего человечества) за 150—200 лет пропускают через свои организмы весь однометровый слой почвы. По представлениям В.И. Вернадского, практически все осадочные породы, а это слой до 3 км, на 95—99 % переработаны живыми организмами.
Высокая скорость обновления живого вещества. Подсчитано, что в среднем для биосферы она составляет 8 лет, при этом для суши — 14 лет, а для океана, где преобладают организмы с коротким периодом жизни (например, планктон), — 33 дня. В результате высокой скорости обновления живого вещества за всю историю существования жизни общая масса живого вещества, прошедшего через биосферу, примерно в 12 раз превышает массу Земли. Только небольшая часть его (доли процента) законсервирована в виде органических остатков (по выражению В.И. Вернадского, ушла в геологию), остальная же включилась в процессы круговорота.
Все перечисленные и другие свойства живого вещества обусловливаются концентрацией в нем больших запасов энергии. По В.И. Вернадскому, по энергетической насыщенности с живым веществом может соперничать только лава, образующаяся при извержении вулканов.
Выделяют следующие основные геохимические функции живого вещества.
1. Энергетическая (биохимическая) — связывание и запасание солнечной энергии в органическом веществе и последующее рассеяние энергии при потреблении и минерализации органического вещества. Эта функция связана с питанием, дыханием, размножением и другими процессами жизнедеятельности организмов. Основной источник биогеохимической активности организмов — солнечная энергия, используемая в процессе фотосинтеза зелеными растениями и некоторыми микроорганизмами для создания органического вещества, обеспечивающего пищей и энергией все остальные организмы.
Газовая — способность изменять и поддерживать определенный газовый состав среды обитания и атмосферы в целом. В частности, включение углерода в процессы фотосинтеза, а затем в цепи питания обусловливало аккумуляцию его в биогенном веществе (органические остатки, известняки и т.п.). В результате этого шло постепенное уменьшение содержания углерода и его соединений, прежде всего двуокиси, в атмосфере с десятков процентов до современных 0,03 %. Это же относится к накоплению в атмосфере кислорода, образованию озона и другим процессам. С газовой функцией живого вещества связаны два переломных периода (точки) в развитии биосферы. Первая из них относится ко времени, когда содержание кислорода в атмосфере достигло примерно 1 % от современного уровня (первая точка Пастера). Это обусловило появление первых аэробных организмов (способных жить только в среде, содержащей кислород). С этого времени восстановительные процессы в биосфере стали дополняться окислительными. Это произошло примерно 1,2 млрд лет назад. Второй переломный период связывают со временем, когда концентрация кислорода достигла примерно 10 % от современной (вторая точка Пастера). Это создало условия для синтеза озона и образования озонового слоя в верхних слоях атмосферы, что обусловило возможность освоения организмами суши (до этого функцию защиты организмов от губительных ультрафиолетовых лучей выполняла вода, под слоем которой возможна была жизнь).
Концентрационная — «захват» из окружающей среды живыми организмами и накопление в них (в большей степени, чем в окружающей среде) атомов биогенных химических элементов. Питание, дыхание и размножение организмов и связанные с ними процессы создания, накопления и распада органического вещества обеспечивают постоянный круговорот вещества и энергии. С этим круговоротом связана миграция атомов химических элементов (прежде всего биогенных — С, Н, О, N, P, S, Fe, Mg, Ca, Na, К, Mo, Mn, Cu, Zn и др.). В ходе биогеохимических циклов атомы большинства химических элементов проходили через живое вещество бесчисленное число раз. Так, например, весь кислород атмосферы оборачивается через живое вещество за 2000 лет, углекислый газ — за 200 (300) лет, а вся вода биосферы — за 2 млн лет. Разные организмы в разной степени способны аккумулировать из среды обитания различные элементы, например, железобактерии накапливают железо; простейшие фораминиферы, а также многие моллюски и кишечнополостные — кальций; хвощи, диатомовые водоросли, радиолярии и др. — кремний; губки — йод; асцидии — ванадий, и т.д. Концентрационная способность живого вещества повышает содержание атомов химических элементов в организмах по сравнению с окружающей средой на несколько порядков. Содержание углерода в растениях в 200 раз, а азота в 30 раз превышает их уровень в земной коре. Содержание марганца в некоторых бактериях может быть в миллионы раз больше, чем в окружающей среде. Результат концентрационной деятельности живого вещества — образование залежей горючих ископаемых, известняков, рудных месторождений и т.п.
Окислительно-Восстановительная — окисление и восстановление различных веществ с помощью живых организмов. Под влиянием живых организмов происходит интенсивная миграция атомов элементов с переменной валентностью (Fe, Mn, Сг, S, P, N, W), создаются их новые соединения, происходит отложение сульфидов и минеральной серы, образование сероводорода и т.п.
Деструктивная — разрушение организмами и продуктами их жизнедеятельности, в том числе и после их смерти, как остатков органического вещества, так и косных веществ. Наиболее существенную роль в этом отношении выполняют редуценты (деструкторы) — сапротрофные грибы и бактерии.
Транспортная — перенос вещества и энергии в результате активной формы движения организмов. Такой перенос может осуществляться на огромные расстояния, например, при миграциях и кочевках животных. С транспортной функцией в значительной мере связана концентрационная роль сообществ организмов, например в местах их скопления (птичьи базары и другие колониальные поселения).
Средообразующая — преобразование физико-химических параметров среды. Эта функция является в значительной мере интегральной — представляет собой результат совместного действия других функций. Она имеет разные масштабы проявления. Результатом средообразующей функции является и вся биосфера, и почва как одна из сред обитания, и более локальные структуры. К средообразующим свойствам растительного покрова относятся: создание микроклимата, очистка воздуха и вод от загрязняющих веществ, усиление питания грунтовых вод, защита почв от эрозии и т.п.
Рассеивающая — функция, противоположная концентрационной — рассеивание веществ в окружающей среде. Она проявляется через трофическую и транспортную деятельность организмов. Например, рассеивание вещества при выделении организмами экскрементов, смене покровов и т.п. Железо гемоглобина крови рассеивается кровососущими насекомыми.
9 Информационная — накопление живыми организмами определенной информации, закрепление ее в наследственных структурах и передача последующим поколениям. Это одно из проявлений адаптационных механизмов.
10. Биогеохимическая деятельность человека — превращение и перемещение веществ биосферы в результате человеческой деятельности для хозяйственных и бытовых нужд человека. Например, использование концентраторов углерода — нефти, угля, газа и др.
Таким образом, биосферу можно также определить как сложную динамическую систему, осуществляющую улавливание, накопление и перенос энергии путем обмена веществ между живым веществом и окружающей средой.
Биосфера обладает рядом свойств.
Целостность и дискретность. Целостность биосферы обусловлена тесной взаимосвязью слагающих ее компонентов. Она достигается круговоротом вещества и энергии. Изменение одного компонента неизбежно приводит к изменению других и биосферы в целом. При этом биосфера — не механическая сумма компонентов, а качественно новое образование, обладающее своими особенностями и развивающееся как единое целое. Биосфера — система с прямыми и обратными (отрицательными и положительными) связями, которые, в конечном счете, обеспечивают механизмы ее функционирования и устойчивости. На понимании целостности биосферы основывается теория и практика рационального природопользования. Учет этой закономерности позволяет предвидеть возможные изменения в природе, дать прогноз результатам воздействия человека на природу.
Централизованность. Центральным звеном биосферы выступают живые организмы (живое вещество). Это свойство, к сожалению, часто недооценивается человеком и в центр биосферы ставится только один вид — человек (идеи антропоцентризма).
Устойчивость и саморегуляция. Биосфера способна возвращаться в исходное состояние, гасить возникающие возмущения, создаваемые внешними и внутренними воздействиями, включением определенных механизмов. Гомеостатические механизмы биосферы связаны в основном с живым веществом, его свойствами и функциями. Биосфера за свою историю пережила ряд таких возмущений, многие из которых были значительными по масштабам (извержения вулканов, встречи с астероидами, землетрясения и т.п.). Гомеостатические механизмы биосферы подчинены принципу Ле Шателье—Брауна: при действии на систему сил, выводящих ее из состояния устойчивого равновесия, последнее смещается в том направлении, при котором эффект этого воздействия ослабляется.
Ритмичность. Биосфера проявляет ритмичность развития — повторяемость во времени тех или иных явлений. В природе существуют ритмы разной продолжительности. Основные из них — суточный, годовой, внутривековые и сверхвековые. Суточный ритм проявляется в изменении температуры, давления и влажности воздуха, облачности, силы ветра, в явлениях приливов и отливов, циркуляции бризов, процессах фотосинтеза у растений, поведении животных. Годовая ритмика — это смена времен года, изменения в интенсивности почвообразования и разрушения горных пород, сезонность в хозяйственной деятельности человека. Суточная ритмика, как известно, обусловлена вращением Земли вокруг оси, годовая — движением Земли по орбите вокруг Солнца. Разные экосистемы обладают различной суточной и годовой ритмикой. Годовая ритмика лучше всего выражена в умеренном поясе и очень слабо — в экваториальном. Наблюдаются и более продолжительные ритмы (11, 22—23, 80—90 лет и др.). Ритмические явления не повторяют полностью в конце ритма того состояния природы, которое было в его начале. Именно этим и объясняется направленное развитие природных процессов.
Горизонтальная зональность и высотная поясность. Общебиосферной закономерностью является горизонтальная зональность - закономерное изменение природной среды по направлению от экватора к полюсам. Зональность обусловлена неодинаковым количеством поступающего на разные широты тепла в связи с шарообразной формой Земли. Зональный климат, воды суши и океана, процессы выветривания, некоторые формы рельефа, образующиеся под влиянием внешних сил (поверхностных вод, ветра, ледников), растительность, почвы, животный мир.
Наиболее крупные зональные подразделения — географические пояса. Они отличаются друг от друга температурными условиями, а также общими особенностями циркуляции атмосферы, почвенно-растительного покрова и животного мира. На суше выделяются следующие географические пояса: экваториальный и в каждом полушарии субэкваториальный, тропический, субтропический, умеренный, а также в Северном полушарии субарктический и арктический, а в Южном — субантарктический и антарктический. Аналогичные по названию пояса выявлены и в Мировом океане. Географические пояса протягиваются преимущественно в широтном направлении.
Внутри поясов по соотношению тепла и влаги выделяются природные зоны, названия которых определяются по преобладающему в них типу растительности. Так, например, в субарктическом поясе это зоны тундры и лесотундры, в умеренном поясе — зоны лесов, лесостепи, степи, полупустынь и пустынь, в тропическом поясе — зоны лесов, редколесий и саванн, полупустынь и пустынь. Как правило, они совпадают с основными и переходными типами природных экосистем (биомами и экотонами). В связи с неоднородностью земной поверхности, а, следовательно, и увлажнения в различных частях материков зоны не всегда имеют широтное простирание.
Зональность характерна и для Мирового океана. От экватора к полюсам изменяются свойства поверхностных вод (температура, соленость, плотность и прозрачность, интенсивность волнения и др.), а также состав растительности и животного мира.
Высотная поясность — закономерная смена природной среды с подъемом в горы от их подножия до вершин. Она обусловлена изменением климата с высотой: понижением температуры (на 0,6°С на каждые 100 м подъема) и до определенной высоты (до 2—3 км) увеличением осадков. Смена поясов в горах происходит в той же последовательности, как и на равнине при движении от экватора к полюсам. Отличием является присутствие в горах особого пояса субальпийских и альпийских лугов, которого нет на равнинах. Высотная поясность начинается в горах с аналога той горизонтальной зоны, в пределах которой расположены горы. Так, в горах находящихся в степной зоне, нижний пояс горно-степной, в лесной — горно-лесной и т.д. Количество высотных поясов зависит от высоты гор и их местоположения.
Большое разнообразие. Биосфера — система, характеризующаяся большим разнообразием. Это свойство обусловлено следующими причинами: разными средами жизни (водной, наземно-воздушной, почвенной, организменной); разнообразием природных зон, различающихся по климатическим, гидрологическим, почвенным, биотическим и другим свойствам; наличием регионов, различающихся по химическому составу (геохимические провинции); биологическим разнообразием живых организмов.
В настоящее время описано более 2 млн. видов. Однако реальное число видов на Земле в несколько раз больше, чем их описано. Не учтены многие насекомые и микроорганизмы, особенно в тропических лесах, глубинных частях океанов и в других малоосвоенных местообитаниях. Кроме этого, современный видовой состав — это лишь небольшая часть видового разнообразия, которое принимало участие в процессах биосферы за период ее существования. Каждый
вид имеет определенную продолжительность жизни (10—30 млн лет), поэтому число видов, принимавших участие в эволюции биосферы, исчисляется сотнями миллионов. Считается, что к настоящему времени арену биосферы оставили более 95 % видов.
Разнообразие обеспечивает возможность дублирования, подстраховки, замены одних звеньев другими, степень сложности и прочности пищевых и другие связей. Поэтому разнообразие рассматривают как основное условие устойчивости любой экосистемы и биосферы в целом.
К сожалению, практически вся без исключения деятельность человека подчинена упрощению экосистем любого ранга. Сюда следует отнести и уничтожение отдельных видов или резкое уменьшение их численности, и создание агроценозов на месте сложных природных систем. Например, полностью исчезли с лица земли степи как тип экосистем и ландшафтов, резко уменьшились площади лесов (до появления человека они занимали примерно 70 % суши, а сейчас — не более 20-23 %). Идет дальнейшее, невиданное по масштабам, уничтожение лесных экосистем, особенно наиболее ценных и сложных тропических, спрямление русел рек, создание промышленных районов и т.п.
Простые экосистемы с малым разнообразием удобны для эксплуатации, они позволяют в короткое время получить значительный объем нужной продукции (например, с сельскохозяйственных полей), но за это приходится рассчитываться снижением устойчивости экосистем, их распадом и деградацией среды.
Не случайно, что биологическое разнообразие отнесено Конференцией ООН по окружающей среде и развитию (1992 г.) к числу трех важнейших экологических проблем, по которым приняты специальные Заявления или Конвенции. Кроме сохранения разнообразия, такие конвенции приняты по сохранению лесов и по предотвращению изменений климата.
Круговорот веществ — многократное участие веществ в процессах, протекающих в атмосфере, гидросфере и литосфере, в том числе в тех слоях, которые входят в состав биосферы Земли. Круговорот веществ осуществляется при непрерывном поступлении (потоке) внешней энергии Солнца и внутренней энергии Земли.
В зависимости от движущей силы, с определенной долей условности, внутри круговорота веществ можно выделить геологический, биологический и антропогенный круговороты. До возникновения человека на Земле осуществлялись только первые два.
Круговорот веществ и энергозависимость. Биосфера — открытая система. Ее существование невозможно без поступления энергии извне. Основная доля приходится на энергию Солнца. В отличие от количества солнечной энергии, количество атомов вещества на Земле ограничено. Круговорот веществ обеспечивает неисчерпаемость отдельных атомов химических элементов. При отсутствии круговорота, например, за короткое время был бы исчерпан основной «строительный материал» живого — углерод.
Лекция 1 - Понятие о техногенном загрязнении биосферы. Типы загрязнений
По отношению к человечеству биосфера Земли выполняет две важнейшие защитные функции:
- обеспечивает биопродуктивные процессы – снабжает людей биологически доступной энергией в виде пищи;
- создает благоприятные условия среды – температурный, газовый, водный режимы, к которым адаптирован вид ГОМО САПИЕНС.
Способность биосферы успешно реализовывать и длительно поддерживать этот механизм определяется ее устойчивостью. Пригодная для жизни устойчивая среда в биосфере может поддерживаться только при строго сбалансированных внутренних потоках энергии, вещества и информации. Нарушение этих системообразующих связей может вызвать нарушение структуры биосферы и повлечь за собой деградацию природной среды. В последние годы слишком интенсивное, резкое воздействие на природные компоненты, вызывают в биосфере побочные отрицательные эффекты, которые выражаются :
1) в истощении природных ресурсов;
2) в загрязнении окружающей природной среды;
3) в нарушении природных биогеохимических циклов;
4) в разрушении природных экосистем и замене их менее продуктивными и менее устойчивыми антропогенными системами.
Загрязнением биосферы (природной среды) называется любое внесение в экосистему несвойственных ей биотических и абиотических компонентов или структурных изменений, прерывающих потоки вещества и энергии, в результате чего экосистема разрушается и снижается ее продуктивность. Источники загрязнения биосферы могут быть естественными и техногенными. К естественным источникам относят вулканизм, водную и ветряную эрозию почвы, природные пожары и др. Техногенные источники – это объекты хозяйственной деятельности (промышленные предприятия, транспорт, с/х, объекты энергетики).
Техногенное загрязнение по масштабам сопоставимо с естественным, а по токсичности – превышает его. В зависимости от масштабов загрязнение может быть глобальным, региональным и локальным, что в свою очередь определяется соотношением двух факторов – мощности источника загрязнения и свойств среды, в которую это загрязнение поступает. Техногенное загрязнение среды обуславливает значительную часть природоемкости техносферы и приводит к деградации экологических систем, глобальным климатическим и геохимическим изменениям, к региональным и локальным экологическим бедствиям, поражением людей. Среди слагаемых современного экологического кризиса наибольшее внимание привлекают четыре компонента: кислотные дожди, парниковый эффект, загрязнение планеты особо токсичными веществами, радиация и так называемые озоновые дыры. Все они связаны с техногенными эмиссиями и воздействиями. Современные глобальные экологические проблемы возникли на почве столкновения между техносферой и биосферой, столкновение, в котором техносфера играет активную, агрессивную роль, Т.е. это процесс конкурентного вытеснения биосферы техносферой. Поскольку техносфера и биосфера находятся в постоянном взаимодействии, их сумму можно представить как единую систему – экосферу. Но именно человечество, ресурсы и продукты его производства и потребления оказывают серьезное влияние на материал и процессы экосферы, вмешиваются в природный круговорот, изменяя его сбалансированность и гармоничность. В связи с тем, что все геооболочки взаимосвязаны вещественными потоками, загрязнение, как правило, не локализуется в какой-либо одной из них.
Рис. Графическая модель потоков техногенного вещества в биосфере
В зависимости от характера вызываемых в природной бреде изменений загрязнения делятся на:
- физические – изменение физических параметров среды;
- химические - изменение химического состава среды;
- биологические - изменение видовой структуры биоценозов.
К физическому относят тепловое, световое, шумовое, ультразвуковое и др. загрязнения.
Химическое загрязнение обусловлено миграциями химических элементов, большинство которых возвращается в природный круговорот в необычных, несвойственных биосфере формах. Особую опасность представляют искусственно созданные органические соединения (ксенобиотики).
По своему токсическому действию загрязняющие вещества делятся на:
Общесоматические (оксид углерода, соединения свинца, ртути, мышьяка, др.);
Раздражающие (сернистый ангидрид, аммиак, оксиды азота, озон, ацетон, др.);
Аллергеные (большинство красителей, лаки, и др.);
Канцерогенные (бенз(а)пирен, асбест, соединения никеля);
Мутагенные (соединения радия, урана, марганца, свинца).
Они могут находиться в газообразном, жидком и твердом состоянии, загрязняя атмосферу, почву, и гидросферу.
Под биологическим загрязнением понимают изменение видовой структуры, путем интродукции несвойственных для данного биоценоза видов (колорадский жук, моллюск рапана и др.).
Массовое размножение микроорганизмов на антропогенных субстратах или техногенно измененных средах носит название микробиологического загрязнения. Это загрязнение может не только явиться причиной заболеваний людей и домашних животных, но и вызвать усиленную коррозию металлов.
К особому классу загрязнений относят деструктивное загрязнение – непреднамеренное структурное изменение ландшафтов, вызванное хозяйственной деятельностью (добыча полезных ископаемых открытым способом, строительство дорог, лесозаготовки и др.).
Количество загрязняющих веществ в мире огромно, и число их по мере развития новых технологических процессов постоянно растет. В этом отношении «приоритет», как в локальном, так и в глобальном масштабе, учете отдают следующим, загрязняющим веществами;
- диоксиду серы (с учетом эффектов вымывания диоксида серы из атмосферы и попадания, образующихся серной кислоты и сульфатов на растительность, почвы и в водоемы);
- тяжелым металлам в первую очередь свинцы, кадмию и особенно ртути (с учетом цепочек ее миграции и превращения в высокотоксичную метилртуть);
- некоторым канцерогенным веществам, в частности бенз(а)пирену;
- нефти и нефтепродуктам в морях и океанах;
- хлорорганическим пестицидам (в сельских районах);
- оксиду углерода и оксидами азота (в городах).
Этот перечень должен быть дополнен радионуклидами и другими радиоактивными веществами, пагубные последствия которых для человеческой популяции и экосистем в полной мере проявились после атомной бомбардировки Хиросимы и Нагасаки и аварии на Чернобыльской АЭС. Следует упомянуть и диоксины – весьма опасное загрязняющее вещество из класса хлоруглеводородов.
Нейтрализацию возмущающих воздействий (загрязнений) в биосфере и ее компонентах за счет механизмов саморегулирования природных экосистем принято называть самоочищением биосферы. Сложный механизм самоочищения природной среды включает систему взаимосвязанных природных процессов:
Механические (перемешивание, разбавление, оседание, перемешивание и др.);
Химические (окисление, осаждение, нейтрализация, фотохимические процессы)
Биохимические (разложение органических веществ микроорганизмами-деструкторами).
Важное место в механизме самоочищения биосферы занимают процессы, происходящие в атмосфере и гидросфере. Это обусловлено тем, что эти две геооболочки представляют собой сплошные подвижные среды, которые обеспечивают:
Кислород для дыхания, углекислый газ для фотосинтеза, воду для жизнедеятельности.
Стабильность и благоприятность для жизни климатических условий земли.
Геохимическую миграцию химических веществ в круговороте.
Быстрое распространение загрязняющих веществ на большие расстояния.
Активную химическую и биологическую трансформацию загрязняющих веществ.
Количественно мерой устойчивости биосферы к техногенным нагрузкам может служить ассимиляционный потенциал – предельная количественная характеристика загрязнения, которое может быть нейтрализовано за счет механизмов саморегулирования экосистем на данной территории или в биосфере в целом. Если величина потока техногенного загрязнения не превышает ассимиляционный потенциал территории, то экосистемы легко справляется с переработкой загрязнения, и состояние природной среды не ухудшается. Если же поток техногенного загрязнения превосходит регуляторные возможности экосистем, то загрязнение накапливается в них, в результате чего на данной территории наблюдается прогрессирующее ухудшение состояния природной среды.
Вокруг источника загрязнения (например, промышленного узла) формируется сфера влияния, размер которой зависит от многих факторов: объема промышленных выбросов, их химического состава, времени функционирования, характера почвенно-растительного покрова, рельефа, климата и др. Проекция сферы влияния на поверхность земли определяет ареал воздействия (загрязнения), форма, которой в той или иной степени определяется рельефом и преимущественным направлением ветра в данной местности. При распространении воздействия на территорию с относительно однородными природными свойствами ареал воздействия должен иметь правильную эллиптическую форму.

РИС.2 Схема внутреннего строения ареала влияния источника загрязнения: 1-источник загрязнения, 2-зона геоматического воздействия, 3-зона биотического воздействия, 4-зона геохимического воздействия.
Сфера воздействия технической системы на природную среду имеет сложное внутреннее строение и состоит из нескольких зон, выявляемых по степени изменения отдельных природных компонентов или структуры ландшафта в целом. Наиболее удаленной от источника загрязнения является зона геохимического загрязнения. Она занимает, как правило, большую площадь (для крупных предприятий и городов – несколько тысяч квадратных километров) и характеризуется повышенным относительно фона содержанием загрязняющих веществ.
Однако, так как уровень общего загрязнения в этой зоне не превышает ассимиляционный потенциал ландшафта, природные комплексы обеспечивают процессы самоочищения.
Зона биотического воздействия выделяется по трансформации биотических компонентов ландшафта (снижению видового разнообразия за счет выпадения наиболее чувствительных к загрязнению видов).
Зона геоматического воздействия (или импактная зона, или техногенная пустошь) характеризуется сильным загрязнением и структурной перестройкой природных комплексов: растительный покров нарушен, активно протекают ветровая и водная эрозия почвы, заболачивание, карст, оврагообразование и др. разрушительные процессы. Эта зона окаймляет промузел. Она формируется либо при малой устойчивости ландшафта (горные районы, зоны вечной мерзлоты и т.п.), либо при большой силе воздействия (химические, горнометаллургические производства).
Лекция 2 - Техногенное воздействие на атмосферу. Последствия загрязнения
Атмосфера – газовая оболочка Земли. Она имеет следующее строение. Наиболее плотный слой воздуха, прилегающий к земной поверхности, носит название тропосферы. Толщина ее в средних широтах составляет 10-12 км над уровнем моря, на полюсах – 7-10, над экватором – 16-18 км. Температура воздуха в тропосфере по высоте уменьшается на 0.6˚ на каждые 100 м и колеблется от 40-50˚С. Выше тропосферы находится стратосфера. Между ними находится тропопауза. Стратосфера имеет протяженность около 40 км. Воздух в ней разряжен, влажность невысокая, температура воздуха от-50˚С до 10˚С. В стратосфере под воздействием космического излучения молекулы воздуха ионизируются, в результате образуется озон. Озоновый слой находится на высоте 25-40 км. Стратопауза отделяет стратосферу от лежащей выше мезосферы. В нем количество озона уменьшается, поэтому средняя температура там значительно ниже. В слое мезосферы наблюдаются серебристые облака. Выше мезосферы расположена ионосфера, между которой находится мезопауза. Для термосферы характерно непрерывное повышение температуры с увеличением высоты. Наиболее удалена от Земли экзосфера (от 800и до 1600 км). Атмосфера выполняет следующие функции:
- содержит кислород, необходимый для дыхания живых организмов;
- является источником углекислого газа для фотосинтеза растений:
- защищает живые организмы от космического излучения;
- сохраняет тепло Земли и регулирует климат;
- трансформирует газообразные продукты обмена веществ;
- переносит водяные пары по планете;
- является средой обитания летающих форм организмов;
- служит источником химического сырья и энергии;
- принимает и трансформирует газообразные и пылевые отходы.
Состав атмосферы находится в состоянии динамического равновесия, поддерживаемого такими климатическими факторами, как перемещения воздушных масс и атмосферные осадки, жизнедеятельность животного и растительного миров, особенно лесов и планктона мирового океана, а также в результате космических процессов, геохимических явлений и хозяйственной деятельности человека.
Атмосферный воздух необходим для дыхания живых организмов (существ), используется в технологических процессах горения и плавки как сырье для получения кислорода, азота, инертных газов, оксида углерода. Атмосфера является средой для размещения газообразных отходов производства, Под воздействием атмосферных осадков, солнечной радиации и в результате переноса воздушных масс атмосферный воздух избавляется от посторонних примесей. Этот процесс называется самоочищением атмосферы.
Примесь в атмосфере – это рассеянное в атмосфере вещество, не содержащееся в постоянном составе.
Загрязняющее воздух вещество – примесь в атмосфере, оказывающая неблагоприятное воздействие на окружающую среду и здоровье населения.
Первичная примесь – примесь сохранившееся за рассматриваемый интервал времени свои физические и химические свойства.
Вторичная примесь – это примесь в атмосфере, образовавшаяся в результате превращения первичных примесей.
Загрязнение атмосферы – поступление в нее химических соединений, которые прямо или косвенно оказывают или могут оказывать в будущем отрицательное воздействие на атмосферу. Техногенные источники загрязнения атмосферы нарушают сложившиеся природные равновесия и вызывают экологический стресс, т.к. техногенная эмиссия загрязняющих веществ осуществляется на 5% суши, где она в 5-20 раз превышает естественную. В насыщенных промышленными предприятиями регионах экосистемы уже не могут выдерживать стрессовые нагрузки («ассимиляционный потенциал территории»). Наиболее распространены (многотонажные) загрязнители сравнительно немногочисленны. Это различные твердые частицы (пыль, дым, сажа), окись углерода, диоксид серы, окислы азота, различные летучие углеводы, соединения фосфора, сероводород, аммиак, хлор, фтористый водород. Наиболее распространенный и значительный по массе загрязнитель – СО (угарный газ), содержание которого в атмосфере крупных городов колеблется от 1-250 млн-1 при среднем значении около 20-1. Очень токсичный газ, который блокирует дыхательную функцию гемоглобина и развитием в последствии кислородной недостаточности. Диоксид серы обладает общесоматическим токсическим действием. Оксиды азота образуются при высокотемпературном окислении азота воздуха кислородом. Во влажном среде в присутствии кислорода превращаются в азотную кислоту. Основным техногенным источником образования оксидом азота является сжигание органического топлива, в том числе автотранспортом. Поступившие из различных источников загрязняющие вещества переносятся в атмосфере упорядоченными воздушными потоками, выпадают на поверхность с атмосферными осадками и вступают в различные химические взаимодействия.
Время, в течение которого в среднем молекула загрязнителя находится в атмосфере, называется временем пребывания. По времени пребывания все загрязнители делят на кратковременные и долговременные. Кратковременные загрязняющие вещества (кислород, окислы азота, ртути и др.) обычно образуют локальные, реже региональные области загрязнения. Долговременные загрязнители (фреоны, оксид углерода и др.) прогрессивно накапливаются в атмосфере. Вертикальные перемещение загрязняющих веществ в атмосфере происходит посредством конвекции, вызванной градиентом температуры.
Горизонтальное распределение загрязняющего вещества зависит от его времени пребывания и от направления и скорости ветра. При средней скорости западных потоков, наблюдаемых в верхней и нижней стратосфере умеренных широт, загрязнители успевают за 10-12 суток обогнуть земной шар. Скорость движения воздуха в меридиональном направлении значительно меньше зональной скорости.
Под действием солнечной радиации находящиеся в атмосфере загрязнители вступают в фотохимические реакции. В этих превращениях участвуют кислород, озон, оксиды азота, серы, углерода. Продукты фотохимических трансформаций, претерпеваемых загрязняющими веществами в окислительной среде атмосферы под действием солнечной радиации, могут оказаться еще более опасными, чем исходные соединения. Особенно опасен фотохимический смог, образующийся в крупных городах вследствие скопления выхлопных газов автотранспорта. В условиях сильной солнечной радиации из них образуется вещества, значительно превосходящие исходные по своей токсичности: озон, пероксиацетилнитрат, пероксибензоилнитрат и др. Компоненты фотохимического смога раздражают слизистые глаз и дыхательных путей, приводят к спазмам в грудной клетке, а при высоких концентрациях вызывают сильный кашель и нейрофизиологические отклонения.
В мире численность автомобилей превышает 400 млн. единиц 80% из которых приходится на легковые, 15-17% - на грузовые и автобусы. По подсчетам, в автомобильных двигателях внутреннего сгорания ежегодно сгорает около 2 млрд. т. нефтяного топлива, а если учитывать, что коэффициент полезного действия двигателя не превышает 23%, то остаток топлива используется не только на обогрев, но и загрязнение окружающей среды. В местах с развитой промышленностью до 80% всех загрязнений дает автотранспорт.
Факторами загрязнения атмосферы автотранспортом являются:
- отработанные газы, выбрасываемые через выхлопную трубу;
- картерные газы;
- углеводородные газы с бака, карбюратора и трубопроводов вследствие испарения и протекания.
Выбросы автотранспорта содержат до 200 химических соединений, некоторые из них чрезвычайно токсичны (CO, CO2, свинец, бенз(а)пирен). Только один грузовой автомобиль, работающий на бензине, при сгорании 1т. топлива выбрасывает в воздух до 0.6т. CO (угарного газа), а за год это составляет 8-10 тонн. В двигателях внутреннего сгорания на сжигание 1т. бензина расходуется около 15т. воздуха или 2000л. О2 что в 2.5 раза больше суточного потребления О2 человеком. При сгорании 1000л. топлива карбюраторный двигатель выбрасывает в атмосферу: 200кг. СО2, 25кг. углеводородов, 20кг. оксидов азота, 1кг. оксидов серы, 1кг. сажи.
Для уменьшения негативного влияния транспорта на окружающую среду существует несколько путей:
- установление в городах скорости автомобильного транспорта 60км/час, при которой количество выхлопных газов наименьшее;
- проектирование объездных путей для транзитного транспорта;
- создание дорожных развязок на двух или трех уровнях с целью уменьшения количества остановок перед светофорами, когда резко увеличивается выброс газов;
- расширение сети электротранспорта и метрополитена;
- перевод автотранспорта на экологически чистое топливо или природный газ;
- замена антидетонационной добавки к бензину (этиловым спиртом);
- снижение затрат топлива и др.
Высокие концентрации примесей и их миграция в атмосферном воздухе приводят к образованию более токсичных веществ (смог, кислоты) или к таким явлениям как парниковый эффект и разрушение озонового слоя.