

2. Первичный транспорт
Первичный транспорт - это такой транспорт, когда энергия расходуется непосредственно на перенос частиц. Он включает, во-первых, перенос отдельных ионов вопреки концентрационному и электрическому градиентам с помощью специальных ионных насосов, во-вторых, эндоцитоз, экзоцитоз и трансцитоз (микровезикулярный транспорт).
Транспорт веществ с помощью насосов (помп). Насосы представляют собой белковые молекулы, обладающие свойствами переносчика и АТФазной активностью. Непосредственным источником энергии являются АТФ. Достаточно хорошо изучены Na/K-, Са- и Н- насосы. Рассмотрим основные характеристики насосов.
1. Специфичность насосов заключается в том, что они обычно переносят какой-то определенный ион или 2 иона. Например, Na/K-насос (объединенный насос для Na+ и К+) не способен переносить ион лития, хотя по своим свойствам он очень близок к натрию.
2. Характеристика отдельных насосов. Натрий-калиевый насос (Na/K-АТФаза) - это интегральный белок клеточной мембраны, обладающий, как и все другие насосы, свойствами фермента, т.е. сам переносчик обеспечивает расщепление АТФ и освобождение энергии, которую он же сам использует. Этот насос изучен наиболее хорошо, он имеется в мембранах всех клеток и создает характерный признак живого - градиент концентрации Na+ и К+ внутри и вне клетки, что обеспечивает формирование мембранного потенциала и вторичный транспорт веществ. Главными активаторами насоса являются гормоны (альдостерон, тироксин). Работа натриевого насоса после удаления К+ из среды сильно нарушается. Кальциевый насос локализуется в эндоплазматическом ретикулуме, он обеспечивает транспорт ионов Са2+. Насос строго контролирует содержание ионов Са в клетке, поскольку изменение уровня Са нарушает ее функцию. Насос переносит ионы Са либо во внеклеточную среду, либо в цистерны ретикулума и митохондрии (внутриклеточное депо ионов Са).
3. Постоянная работа насосов необходима для поддержания концентрационных градиентов ионов, связанного с ними электрического заряда клетки и движения воды и незаряженных частиц в клетку и из клетки вторично активно согласно законам диффузии и осмоса. Эти процессы обеспечивают жизнедеятельность нейрона, как и любой другой клетки. В результате разной проницаемости клеточной мембраны для различных ионов и постоянной работы ионных помп концентрация ионов внутри и снаружи клетки неодинакова. Ионы являются заряженными частицами, поэтому существует электрический заряд нейрона. Почти во всех изученных клетках внутреннее содержимое их заряжено отрицательно по отношению к внешней среде, т.е. внутри клетки преобладают отрицательные ионы, а снаружи - положительные.
Ионы К+ находятся преимущественно в клетке, а ионы Na+ и Сl - во внеклеточной жидкости. Внутри клетки расположены также крупномолекулярные (в основном белкового происхождения) анионы. Na/K-насос транспортирует не только ионы Na+ и К+, но и другие молекулы, например глюкозу, аминокислоты. Более трети энергии АТФ, потребляемой клеткой в состоянии покоя, расходуется на перенос только ионов Na+ и К+. Это обеспечивает сохранение клеточного объема (осморегуляция), поддержание электрической активности в нервных клетках, транспорт других веществ.
Таким образом, первичный транспорт ионов играет исключительно важную роль в жизнедеятельности клеток.
4. Механизм работы ионных насосов заключается в следующем. Na/K-насос - молекула интегрального белка, пронизывающая всю толщу клеточной мембраны, переносит за один цикл 3 иона Na+ из клетки и 2 иона К+ в клетку (антипорт - противотранспорт). Это осуществляется в результате конформации молекулы белка в форму E1 или Е2. Молекула имеет участок, который связывает либо ион Na+, либо ион К+, - это активный участок. При конформации E1 белковая молекула активной своей частью обращена внутрь клетки и обладает сродством к иону Na+, который присоединяется к белку, в результате чего активируется его АТФаза, обеспечивающая гидролиз АТФ и освобождение энергии. В результате освобождения энергии изменяется конформация молекулы белка: она превращается в форму Е2, в результате чего активный ее участок уже обращен наружу клеточной мембраны. Теперь белок теряет сродство к иону Na+, последний отщепляется от него, а белок-помпа приобретает сродство к иону К+ и соединяется с ним. Это ведет снова к изменению конформации переносчика: форма Е2 переходит в форму E1, активный участок белка снова обращен внутрь клетки. При этом он теряет сродство к иону К+ и последний отщепляется, а белок приобретает снова сродство к иону Na+ - цикл повторяется. Насос является электрогенным, поскольку за один цикл выводится из клетки 3 иона Na+, а возвращаются в клетку 2 иона К+. Энергия расходуется только на перенос ионов Na+. На обеспечение одного цикла работы Na/K-помпы расходуется одна молекула АТФ.
Подобным образом работают Са-АТФазы эндоплазматического ретикулума и клеточной мембраны, с той лишь разницей, что переносятся только ионы Са и в одном направлении - из гиалоплазмы в эндоплазматический ретикулум, а также наружу клетки.
Эндоцитоз и экзоцитоз (микровезикулярный транспорт).
Это еще два первичных (первично активных), близких по механизму транспорта, посредством которых различные материалы переносятся через мембрану либо в клетку (эндоцитоз), либо из клетки (экзоцитоз). С их помощью транспортируются крупномолекулярные вещества (белки, полисахариды, нуклеиновые кислоты), которые не могут транспортироваться по каналам или с помощью насосов.
1. При эндоцитозе клеточная мембрана образует впячивания, или выросты, внутрь клетки, которые, отшнуровываясь, превращаются в пузырьки. Последние затем обычно сливаются с первичными лизосомами, образуя вторичные лизосомы, в которых содержимое подвергается гидролизу - внутриклеточному перевариванию. Продукты гидролиза используются клеткой. Например, выделившийся медиатор нервным окончанием захватывается снова посредством эндоцитоза.
2. Экзоцитоз - процесс, обратный эндоцитозу, это механизм секреции нейрогормонов и нейромедиаторов. Экзоцитозные пузырьки образуются в аппарате Гольджи. В пузырьки упаковываются белки, образовавшиеся в рибосомах эндоплазматического ретикулума. Пузырьки транспортируются посредством сократительного аппарата клетки к клеточной мембране, сливаются с ней, а содержимое клетки выделяется во внеклеточную среду. Энергия АТФ расходуется на деятельность сократительного аппарата клетки. В процессе взаимодействия эндо- и экзоцитоза происходит самообновление клеточной мембраны (кругооборот, рециркуляция): в течение каждого часа в процессе эндоцитоза в разных клетках используется от 3 до 100% клеточной оболочки, но с такой же скоростью происходит ее возобновление в результате экзоцитоза.
3. Трансцитоз сочетает элементы эндо- и экзоцитоза. Это перенос частиц через клетку: например, перенос молекул белка в виде везикул через эндотелиальную клетку капилляров на другую сторону в интерстиций мозга. В данном случае эндоцитозные пузырьки не взаимодействуют с лизосомами, при этом пузырьки могут сливаться друг с другом, образуя каналы, пересекающие всю клетку.
25РАЗДРОЖИМОСТЬ И ВОЗБДИМОСТЬ.ВИДЫ РАЗДРАЖИТЕЛЕЙ.
Раздражимость – это способность клеток, тканей, организма в целом переходить под воздействием факторов внешней или внутренней среды из состояния физиологического покоя в состояние активности. Состояние активности проявляется изменением физиологических параметров клетки, ткани, организма, например изменением метаболизма.
Возбудимость – это способность живой ткани отвечать на раздражение активной специфической реакцией – возбуждением, т.е. генерацией нервного импульса, сокращением, секрецией. Т.е. возбудимость характеризует специализированные ткани – нервную, мышечные, железистые, которые называются возбудимыми. Возбуждение – это комплекс процессов реагирования возбудимой ткани на действие раздражителя, проявляющийся изменением мембранного потенциала, метаболизма и т.д. Возбудимые ткани обладают проводимостью. Это способность ткани проводить возбуждение. Наибольшей проводимостью обладают нервы и скелетные мышцы.
Раздражитель – это фактор внешней или внутренней среды действующий на живую ткань.
Процесс воздействия раздражителя на клетку, ткань, организм называется раздражением.
Все раздражители делятся на следующие группы:
1.По природе
а) физические (электричество, свет, звук, механические воздействия и т.д.)
б) химические (кислоты, щелочи, гормоны и т.д.)
в) физико-химические (осмотическое давление, парциальное давление газов и т.д.)
г) биологические (пища для животного, особь другого пола)
д) социальные (слово для человека).
2.По месту воздействия:
а) внешние (экзогенные)
б) внутренние (эндогенные)
3.По силе:
а) подпороговые (не вызывающие ответной реакции)
б) пороговые (раздражители минимальной силы, при которой возникает возбуждение)
в) сверхпороговые (силой выше пороговой)
4.По физиологическому характеру:
а) адекватные (физиологичные для данной клетки или рецептора, которые приспособились к нему в процессе эволюции, например, свет для фоторецепторов глаза).
б) неадекватные
Если реакция на раздражитель является рефлекторной, то выделяют также:
а) безусловно-рефлекторные раздражители
б) условно-рефлекторные
По биологическому значению все раздражители относят к адекватным инеадекватным.
Адекватным считается такой раздражитель, к восприятию которого данная биосистема специально приспособилась в процессе эволюции. Так, для органа зрения адекватно электромагнитное воздействие в определенном диапазоне длин волн; для слуха — упругие механические колебания среды и т.п.
К Неадекватным относят раздражители, не являющиеся в естественных условиях средством возбуждения данной биосистемы, но, тем не менее, способные при достаточной силе вызвать возбуждение. Все раздражители (адекватные и неадекватные) в зависимости от их силы подразделяют на пороговые, подпороговые, максимальные, субмаксимальные и супермаксимальные.
Минимальная сила раздражителя, необходимая для возникновения минимального по величине возбуждения, называется порогом возбуждения. Величина порога является мерой возбудимости ткани.
Раздражители, сила которых ниже порога возбуждения, рассматриваются какподпороговые.
Если сила раздражения превосходит порог возбуждения, величина ответной реакции ткани (возбуждения) возрастает вплоть до известного, определенного для каждого живого образования предела. Дальнейшее увеличение силы раздражителя уже не ведет к росту ответной реакции.
Минимальная сила раздражителя, вызывающая наибольший (максимальный) ответ ткани, называется максимальной силой раздражения.
Раздражители, сила которых меньше или больше максимальной, называются, соответственно, субмаксимальными и супермаксимальными.
________________________
26ПОРОГ РАЗДРАЖЕНИЯ КАК МЕРА ВОЗБУДИМОСТИ.
ПОРОГ РАЗДРАЖЕНИЯ, порог возбудимости, минимальная интенсивность раздражения, способная вызвать распространяю
щийся потенциал действия; мера возбудимости клетки или организма в целом. П. р. зависит от силы и качества раздражителя, длительности его воздействия и градиента нарастания силы, а также от свойств и физиол. состояния возбудимой ткани в момент раздражения. Сила раздражения меньше пороговой, т. е. не вызывающая возбуждения, наз. подпороговой, а больше пороговой — надпороговой. Чем ниже П. р., тем выше возбудимость.
Порог раздражителя - та минимальная сила, при действии которой возникает возбуждение. Законы раздражения Действие раздражителя описывается несколькими законами: 1. Закон силы раздражения: Чем больше сила раздражения, тем, до известных пределов, сильнее ответная реакция. Раздражители имеют нижний предел - подпороговое раздражение не вызывает ответной реакции. Возбудимые ткани работают только на пороговых и надпороговых раздражителях. Но есть сила раздражения для любого биологического раздражителя, которая способна вызывать max эффект - оптимальная сила (оптимум частоты и силы раздражения). Если сила больше, чем оптимальная, то ответная реакция ниже - пессимум частоты или силы раздражения. 2. Закон длительности раздражения: Чем длительнее раздражение, необходимое для возникновения возбуждения, тем сильнее, до известных пределов, ответная реакция живых систем. Есть зависимость между силой раздражения и временем, в течение которого этот раздражитель должен действовать, чтобы вызвать ответную реакцию. Зависимость выражается гиперболой, следовательно, даже сильные раздражители, действуя кратковременно, либо не способны вызвать ответную реакцию, либо - слабую ответную реакцию и наоборот. График "сила-время" Особенно чётко зависимость между силой и временем в прослеживается в диапазоне промежуточных величин. 3. Закон градиента силы: Величина ответной реакции и её характер зависят ещё и от интенсивности/крутизны/ нарастания действия силы. Более интенсивное нарастание силы раздражения вызывает больший ответ. При этом длительное действие раздражителей одной и той же по величине силы, приводит к развитию аккомодации - явления, которое выражается в понижении чувствительности ткани к раздражению, уменьшению возбудимости ткани. Механизм этого явления бкдет рассмотрен в следующей лекции. 4. Закон "всё или ничего": Если раздражитель меньше пороговой силы, он никогда не вызовет ПД (потенциал действия) - "ничего". Но какой бы силы ни был надпороговый раздражитель, он всегда будет вызывать max для данного состояния электрическую реакцию, т.е. max пик ПД - "всё". Ответная реакция, её характер зависят от скорости химических процессов обеспечивающих ответные реакции, так называемые скорости активационных и инактивационных /восстановительных/ процессов. Введенский назвал свойство клеток, тканей, связанное со скорость активационных и инактивационных процессов - лабильность (функциональная подвижность)-свойство клетки, ткани, отражающее их максимальные возможности. Мера лабильности - это максимальная частота, которую способна воспроизвести ткань или клетка. Характеризуется способностью ткани отвечать ПД на каждое раздражение. У каждой ткани лабильность различна: в синапсах - 40-50 раз в сек., в периферических нервах - до 20000 раз в сек. Если лабильность ткани превышена, то ткань отвечает либо снижением ответной реакции, либо, если Вы долго будете принуждать ткань работать в режиме большем, чем лабильность - гибелью (это своего рода защитная реакция). Вот почему раздражение выше - по силе или по частоте - чем то, которое вызывает максимальный ответ - вызывает снижение ответа - пессимум (то, о чем мы говорили чуть раньше - при разборе закона силы - вот почему сверхсильные раздражители не дают сверхсильной реакции - они дают в здоровом организме снижение эффекта - это своеобразная защитная реакция). Частота раздражения близкая или совпадающая с величиной лабильности вызывает максимальный ответ, т.е. является оптимальной/ оптимум частоты раздражения/ 27ПОТЕНЦИАЛ ПОКОЯ,МЕХАНИЗМЫ ВОЗНИКНОВНИЯ.
Между наружной поверхностью клетки и ее цитоплазмой в состоянии покоя существует разность потенциалов около 0,06-0,09 в, причем поверхность клетки заряжена электроположительно по отношению к цитоплазме. Эту разность потенциалов называютпотенциалом покоя или мембранным потенциалом. Точное измерение потенциала покоя возможно только с помощью микроэлектродов, предназначенных для внутриклеточного отведения токов, очень мощных усилителей и чувствительных регистрирующих приборов - осциллографов.
Микроэлектрод представляет собой тонкий стеклянный капилляр, кончик которого имеет диаметр около 1 мкм. Этот капилляр заполняют солевым раствором, погружают в него металлический электрод и соединяют с усилителем и осциллографом. Как только микроэлектрод прокалывает покрывающую клетку мембрану, луч осциллографа отклоняется вниз из своего исходного положения и устанавливается на новом уровне. Это свидетельствует о наличии разности потенциалов между наружной и внутренней поверхностью клеточной мембраны. Наиболее полно происхождение потенциала покоя объясняет так называемая мембранно-ионная теория. Согласно этой теории все клетки покрыты мембраной, имеющей неодинаковую проницаемость для различных ионов. В связи с этим внутри клетки в цитоплазме в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем на поверхности. В состоянии покоя клеточная мембрана более проницаема для ионов калия, чем для ионов натрия. Диффузия положительно заряженных ионов калия из цитоплазмы на поверхность клетки придает наружной поверхности мембраны положительный заряд.
Таким образом, поверхность клетки в покое несет на себе положительный заряд, тогда как внутренняя сторона мембраны оказывается заряженной отрицательно за счет ионов хлора, аминокислот и других крупных органических анионов, которые через мембрану практически не проникают Формирование потенциала покоя[править | править исходный текст]
ПП формируется в два этапа.
Первый этап: создание незначительной (-10 мВ) отрицательности внутри клетки за счёт неравного асимметричного обмена Na+ на K+ в соотношении 3 : 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии АТФ, обеспечивает его электрогенность.
Результаты деятельности мембранных ионных насосов-обменников на первом этапе формирования ПП таковы:
1. Дефицит ионов натрия (Na+) в клетке.
2. Избыток ионов калия (K+) в клетке.
3. Появление на мембране слабого электрического потенциала (-10 мВ).
Второй этап: создание значительной (-60 мВ) отрицательности внутри клетки за счёт утечки из неё через мембрану ионов K+. Ионы калия K+ покидают клетку и уносят с собой из неё положительные заряды, доводя отрицательность до -70 мВ.
Итак, мембранный потенциал покоя - это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.
28ПОТЕНЦИАЛ ДЕЙСТВИЯ,ЕГО ФАЗЫ,
Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембранеживой клетки в процессе передачи нервного сигнала. По сути своей представляет электрический разряд — быстрое кратковременное изменениепотенциала на небольшом участке мембраны возбудимой клетки (нейрона,мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса.
Потенциалы действия могут различаться по своим параметрам в зависимости от типа клетки и даже на различных участках мембраны одной и той же клетки. Наиболее характерный пример различий: потенциал действия сердечной мышцы и потенциал действия большинства нейронов. Тем не менее, в основе любого потенциала действия лежат одни и те же явления. нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания.
Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала, равного -90 мВ.
Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь аксона. Нормальное поляризованное состояние в -90 мВ немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении.
Этот процесс называют деполяризацией, В крупных нервных волокнах значительный избыток входящих внутрь положительных ионов натрия обычно приводит к тому, что мембранный потенциал «проскакивает» за пределы нулевого уровня, становясь слегка положительным. В некоторых более мелких волокнах, как и в большинстве нейронов центральной нервной системы, потенциал достигает нулевого уровня, не «перескакивая» его.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов. Электроупавляемые натриевые и калиевые каналы. Необходимым участником процессов деполяризации и реполяризации во время развития потенциала действия в мембране нервного волокна является электроуправляемый натриевый канал. Электроуправляемый калиевый канал также играет важную роль в увеличении скорости реполяризации мембраны. Оба типа электроуправляемых каналов существуют дополнительно к Na+/K+ -насосу и каналам К*/Na+-утечки. Электроуправляемый натриевый канал. В верхней части рисунка показан электроуправляемый натриевый канал в трех различных состояниях. Этот канал имеет двое ворот: одни вблизи наружной части канала, которые называют активационными воротами, другие — у внутренней части канала, которые называют инактивационными воротами. В верхней левой части рисунка изображено состояние этих ворот в покое, когда мембранный потенциал покоя равен -90 мВ. В этих условиях активационные ворота закрыты и препятствуют поступлению ионов натрия внутрь волокна. Активация натриевого канала. Когда мембранный потенциал покоя смещается в направлении менее отрицательных значений, поднимаясь от -90 мВ в сторону нуля, на определенном уровне (обычно между -70 и -50 мВ) происходит внезапное конформационное изменение актива-ционных ворот, в результате они переходят в полностью открытое состояние. Это состояние называют активированным состоянием канала, при котором ионы натрия могут свободно входить через него внутрь волокна; при этом натриевая проницаемость мембраны возрастает в диапазоне от 500 до 5000 раз. Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации. Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
Фазы потенциала действия
Фаза деполяризации. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциалчувствительных Na+-каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсиейзаряда мембраны.
Фаза быстрой и медленной реполяризации . В результате деполяризации мембраны происходит открытие потенциалчувствительных К+ -каналов. Положительно заряженные ионы К+ выходят из клетки по градиенту концентрации (калиевый ток), что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока снижается и реполяризация замедляется.
Фаза гиперполяризации развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+ / K+ помпы.
Овершут – период времени, в течение которого мембранный потенциал имеет положительное значение.
Пороговый потенциал – разность между мембранным потенциалом покоя и критическим уровнем деполяризации. Величина порогового потенциала определяет возбудимость клетки – чем больше пороговый потенциал, тем меньше возбудимость клетки.
29ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ ПРИ ВОЗНИКНОВЕНИИ ВОЗБУЖДЕНИЯ.
Возбудимость – способность клетки переходить из состояния покоя в состояние возбуждения при действии раздражителя. Разные клетки имеют различную возбудимость. Возбудимость одной и той же клетки меняется в зависимости от ее функционального состояния.
Изменение возбудимости клетки при развитии возбуждения
Если принять уровень возбудимости клетки в состоянии физиологического покоя за норму, то в ходе развития цикла возбуждения можно наблюдать ее колебания. В зависимости от уровня возбудимости выделяют следующие состояния клетки
• Супернормальная возбудимость (экзальтация) – состояние клетки, в котором ее возбудимость выше нормальной. Супернормальная возбудимость наблюдается во время начальной деполяризации и во время фазы медленной реполяризации. Повышение возбудимости клетки в эти фазы ПД обусловлено снижением порогового потенциала по сравнению с нормой.
• Абсолютная рефрактерность – состояние клетки, в котором ее возбудимость падает до нуля. Никакой, даже самый сильный, раздражитель не может вызвать дополнительного возбуждения клетки. Во время фазы деполяризации клетка невозбудима, поскольку все ее Na+ -каналы уже находятся в открытом состоянии.
• Относительная рефрактерность – состояние, в котором возбудимость клетки значительно ниже нормальной; только очень сильные раздражители могут вызвать возбуждение клетки. Во время фазы реполяризации каналы возвращаются в закрытое состояние и возбудимость клетки постепенно восстанавливается.
• Субнормальная возбудимость характеризуется незначительным снижением возбудимости клетки ниже нормального уровня. Это уменьшение возбудимости происходит вследствие возрастания порогового потенциала во время фазы гиперполяризации.
30ОПТИМУМ И ПЕССИМУМ ЧАСТОТЫ.
Повышение частоты и силы раздражения до известного предела вызывает увеличение высоты тетанического сокращения скелетной мышцы. Наиболее благоприятная частота нервных импульсов, поступающих в скелетную мышцу, вызывает наибольшую высоту тетануса. Эта частота называется оптимальной, или оптимумом частоты. Оптимуму частоты соответствует такая частота, при которой каждое последующее раздражение застает скелетную мышцу в состоянии наибольшей возбудимости, наблюдающейся в экзальтационной фазе. Поэтому высота каждого одиночного сокращения возрастает. Наоборот, если каждое последующее раздражение застает скелетную мышцу в фазе абсолютной рефрактерности, то тетаническое сокращение мышцы резко уменьшается или не наступает. Эта чрезмерно большая частота — наихудшая, пессимальная, или пессимум частоты. Оптимум частоты соответствует высокому уровню лабильности нерва и мышцы, а пессимум частоты — низкому уровню лабильности нерва, даже более низкому, чем лабильность мышцы
. 1. Закон оптимума
выражается в том, что любой экологический фактор имеет пределы положительного влияния на живые организмы.
Сила воздействия экологических факторов постоянно меняется. Лишь в определенных местах планеты значения некоторых из них более или менее постоянны (константны). Например: на дне океанов, в глубинах пещер сравнительно постоянны температурный и водный режимы, режим освещения.
Рассмотри действие закона оптимума на конкретном примере: животные и растения плохо переносят и сильную жару, и сильные морозы, оптимальными для них являются средниетемпературы - так называемая зона оптимума. Чем сильнее отклонения от оптимума, тем в большей степени данный экологический фактор угнетает жизнедеятельность организма. Эта зона носит название зоны пессимума. В ней имеются критические точки - "максимальное значение фактора" и "минимальное значение фактора"; за их пределами наступает гибель организмов.Расстояние между минимальным и максимальным значениями фактора называют экологической валентностью или толерантностью организма (рис. 3).

Рис. 3. Схема действия факторов среды на живые организмы.
Пример проявления данного
закона: яйца аскарид развиваются при 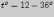 ,
а оптимальной для их развития является
,
а оптимальной для их развития является 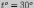 .
То есть экологическая толерантность
аскарид по температурному режиму
составляет от
.
То есть экологическая толерантность
аскарид по температурному режиму
составляет от 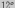 до
до 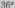 .
.
По характеру толерантности выделяют следующие виды:
эврибионтные - имеющие широкую экологическую валентность по отношению к абиотическим факторам среды; делятся на эвритермные (выносящие значительные колебания температур),эврибатные (выносящие широкий диапазон показателей давления), эвригалинные (выносящие разную степень засоленности среды).
стенобионтные - неспособные переносить значительные колебания фактора (например, стенотермными являются белые медведи, ластоногие млекопитающие, обитающие при низком температурном режиме).
Нейрофизиология 41-50:
41)Продолговатый мозг
(лат. Myelencephalon,
Medulla
oblongata) — отдел
головного
мозга. Встречается также
традиционное название
bulbus
( луковица, из-за формы
этого
отдела).Имеет вид
усеченного конуса.
Функции продолговатого
мозга
1. Защитные рефлексы
(например, кашель,
чихание).
2. Жизненно важные
рефлексы
(например, дыхание).
3. Регулирование
сосудистого
тонуса.
4. Регулирование
дыхательной
системы
Рефлекторные центры
продолговатого мозга:
1. пищеварение
2. сердечная деятельность
3. защитная (кашель,
чихание
и тому подобное)
4. центры регуляции тонуса
скелетных мышц для
поддержания позы
человека.
5. укорочение или
удлинение
времени спинального
рефлекса.
42)Через варолиев мост
проходит
пирамидный
(двигательный) путь,
пути от коры к мозжечку,
общечувствительный путь,
путь от
ядер слухового нерва. В
варолиевом мосту
находится
несколько ядер, в том числе
ядра
слухового, лицевого и
тройничного нервов.
Варолиев мост представляет
собой
расположенный на
основании
мозга массивный
волокнистый
тяж, ограниченный сзади
продолговатым мозгом,
спереди
ножками мозга; выполняет
проводниковую и
регуляторную
функции. Через мост
проходят все
чувствительные и
двигательные
пути ЦНС, регулирует
вегетативные
реакции (слезотечение,
слюнотечение, жевание,
глотание
и др.), участвует в
образовании
голоса.
43) Средний мозг
(лат. Mesencephalon) —
отдел
головного мозга, древний
зрительный центр. Включен
в
ствол головного мозга.В
глубине покрышки
среднего
мозга (под четверохолмием)
находятся ядра
глазодвигательных нервов,
красные ядра (лат. nuclei
rubri,
управление движением),
чёрное
вещество (лат. substantia
nigra,
инициация движений),
ретикулярная
формация.Функции
среднего мозга
Средний мозг выполняет
следующие функции:
1. центр ориентировочного
рефлекса
2. центр позы
3. центр обработки
первичной
информации (зрение, слух)
4. регулирующую в
продолжительности
актов жевания и глотания
5. тонус мышц.
Мне нравится
1
Поделиться
24 мая в 15:21
Скрыть комментарии

Юлия Черняк
44)С участием ствола мозга
осуществляются сложные
соматические рефлексы, в
каждом
из которых задействованы
ядра
нескольких черепных
нервов.
1. Глазодвигательные
рефлексы
имеют центры,
функционально
объединяющие
чувствительные
ядра тройничного,
преддверно-
улиткового нервов, бугорки
четверохолмия,
двигательные
ядра глазодвигательного,
блокового и отводящего
нервов.
Координация их
деятельности
осуществляется
ретикулярной
формацией ствола мозга, а
также
мозжечком и корой
больших
полушарий. В результате
этих
рефлексов происходит
содружественные движения
глаз в
различных направлениях.
2. Рефлекторный акт
жевания
обеспечивается мышцами,
вызывающими движения
нижней
челюсти и удерживающими
пищу
между зубными рядами.
Афферентная импульсация
возникает с различных
рецепторов слизистой
оболочки
рта и проприорецепторов
аппарата жевания и
распространяется в
основном по
сенсорным волокнам
тройничного
нерва. Центр жевания
(центральный генератор
ритма
жевания) находится в
ретикулярной формации
продолговатого мозга и
моста и
вызывает ритмическое
возбуждение мотонейронов
мышц, поднимающих и
опускающих нижнюю
челюсть.
Генератор ритма жевания
может
быть запущен и от
жевательной
области лобной коры, что
обеспечивает
произвольный
контроль жевания.
Эфферентное
влияние центра жевания
осуществляется через
двигательные ядра V, VII и XII
нервов.
3. Рефлекторный акт
глотания
обеспечивает поступление
пищи
из ротовой полости в
желудок. При
передвижении пищевого
комка из
полости рта в пищевод
происходит
последовательное
возбуждение
рецепторов корня языка,
мягкого
неба, глотки и пищевода.
Импульсация по
чувствительным
волокнам тройничного,
языкоглоточного и
блуждающего
нервов поступает в центр
глотания, который
расположен в
продолговатом мозге и
мосте. Этот
центр функционально
объединяет
нейроны примерно двух
десятков
ядер ствола, шейных и
грудных
сегментов спинного мозга. В
результате этого
обеспечивается
строго координированная
последовательность
сокращения
мышц, которые участвуют в
акте
глотания: мышц мягкого
неба,
глотки, гортани и
надгортанника,
пищевода. Центр глотания
функционально связан с
центром
дыхания, которое
прекращается в
течение каждого
глотательного
акта.
4. Рвотный рефлекс
является
защитной реакцией,
возникающей при
раздражении
рецепторов корня языка,
глотки,
желудка, кишечника,
брюшины,
вестибулярного аппарата.
Афферентная импульсация
по
волокнам языкоглоточного,
блуждающего или
преддверно-
улиткового нервов
поступает в
рвотный центр,
расположенный в
продолговатом мозге. Рвоту
можно
вызвать и
непосредственным
раздражением рвотного
центра
местным патологическим
процессом или химическими
веществами. Эфферентные
импульсы из рвотного
центра
поступают по блуждающему
нерву
к пищеводу, желудку,
кишечнику
и через спинальные
моторные
центры к диафрагме и
мышцам
брюшной стенки,
сокращение
которых приводит к
перемещению содержимого
желудка.
5. Рефлекс кашля является
защитным рефлексом,
возникающим при
раздражении
рецепторов гортани, трахеи
и
бронхов. Импульсация по
чувствительным волокнам
блуждающего нерва
возбуждает
кашлевой центр
продолговатого
мозга, имеющий
эфферентный
выход на спинальные
моторные
центры дыхательных мышц.
Центр
кашля запускает жестко
запрограммированную
последовательность
реакций, в
которой можно выделить
три
фазы:
1) глубокий вдох;
2) сокращение мышц
выдоха на
фоне закрытой голосовой
щели и
сужения бронхов, что
приводит к
резкому повышению
давления в
легких;
3) активный выдох на фоне
мгновенного раскрытия
голосовой
щели, создающий мощный
воздушный поток,
направляемый
за счет напряжения мягкого
неба
через рот.
6. Рефлекс чихания
возникает при
раздражении рецепторов
преимущественно
верхнечелюстной и
частично
глазничной ветви
тройничного
нерва в слизистой оболочке
полости носа, особенно
средней
носовой раковины и
перегородки.
Центр чихания,
расположенный в
продолговатом мозге,
организует
те же центральные системы,
что и
при кашле, но поток
воздуха при
форсированном выдохе на
фоне
быстрого открывания
голосовой
щели и опускания мягкого
неба
направляется
преимущественно
через нос.
Мне нравится
24 мая в 15:28|Ответить
Юлия Черняк
45)Мозжечок
представляет собой
мозговой
центр, который имеет в
высшей
степени важное значение
для
координации и регуляции
двигательной активности и
поддержания позы.
Мозжечок
работает главным образом
рефлекторно, поддерживая
равновесие тела и его
ориентацию в пространстве.
Также он играет важную
роль
(особенно у
млекопитающих) в
локомоции (перемещении в
пространстве).
Соответственно главными
функциями мозжечка
являются:
1. координация движений
2. регуляция равновесия
3. регуляция мышечного
тонуса
4. мышечная память
Мозжечок — это
функциональное
ответвление
главной оси «кора больших
полушарий — спинной
мозг». С
одной стороны, в нём
замыкается сенсорная
обратная
связь, то есть он получает
копию
афферентации
(информации,
передаваемой из спинного
мозга
в кору полушарий
головного
мозга), с другой стороны,
сюда
же поступает копия
эфферентации (информации
от
коры полушарий к
спинному
мозгу) от двигательных
центров.
Говоря техническим
языком,
первая сигнализирует о
текущем
состоянии регулируемой
переменной, а вторая даёт
представление о требуемом
конечном состоянии.
Сопоставляя первое и
второе,
кора мозжечка может
рассчитывать ошибку, о
которой
сообщает в двигательные
центры. Так мозжечок
непрерывно корректирует
и
преднамеренные, и
автоматические движения. У
низших позвоночных
информация в мозжечок
поступает также из
акустической
области, в которой
регистрируются ощущения,
относящиеся к равновесию,
поставляемые ухом и
боковой
линией, а у некоторых даже
от
органа обоняния.
Мне нравится
24 мая в 15:30|Ответить
Юлия Черняк
46) Функции
промежуточного мозга
1. Центр боли и
удовольствия
2. Центр нейрогуморальной
регуляции
3. Центр жажды, голода,
насыщения
4. Центр сна и
бодроствования
5. Центр терморегуляции
Промежуточный мозг
подразделяется на:
Таламический мозг
(лат. thalamencephalon)
Подталамическую область
или
гипоталамус (лат.
hypothalamus)
Третий желудочек, который
является полостью
промежуточного мозга
Зрительный бугор (Таламус)
Надталамическую область
(Эпиталамус)
Заталамическую область
(Метаталамус)
аламус
является подкорковым
центром
всех видов
чувствительности
(болевой, температурной,
тактильной,
проприоцептивной).
Таламус является местом
переключения всех
чувствительных
проводящих
путей, идущих от экстеро-,
проприо- и
интерорецепторов.
Эпиталамус или
надталамическая область
(лат. epithalamus)
располагается
в верхнезадней части
таламуса.
Эпиталамус образует
шишковидное тело
(эпифиз),
которое посредством
поводков
крепится к таламусу.
Шишковидное тело
представляет собой железу
внутренней секреции,
которая
отвечает за синхронизацию
биоритмов организма с
ритмами
окружающей среды.
Метаталамус или
заталамическая область
(лат. metathalamus)
образован
парными медиальным и
латеральным коленчатыми
телами, лежащими позади
таламуса. Медиальное
коленчатое тело находится
позади подушки таламуса.
Оно
является подкорковым
центром
слуха. Латеральное
коленчатое
тело расположено книзу от
подушки. Оно является
подкорковым центром
зрения. Гипоталамус или
подталамическая область
расположен под таламусом.
Гипоталамус включает в
себя
сосцевидные тела,
являющиеся
подкорковыми центрами
обоняния, гипофиз,
зрительный
перекрест, II пары
черепных
нервов, серый бугор,
представляющий собой
вегетативный центр обмена
веществ и терморегуляции.
В
гипоталамусе содержатся
ядра,
контролирующие
эндокринные
и вегетативные
процессы.Третий
желудочек
III желудочек (лат.
ventriculus
tertius) — полость
промежуточного мозга. Он
представляет собой узкое,
расположенное в
сагиттальной
плоскости щелевидное
пространство.
Мне нравится
24 мая в 15:33|Ответить
Юлия Черняк
47) Гипоталамус является
центральной структурой
лимбической системы мозга
и
выполняет многообразные
функции. Часть этих
функций
относится к гормональным
регуляциям, которые
осуществляются через
гипофиз.
Другие функции связаны с
регуляцией биологических
мотиваций. К ним относят
потребление пищи и
поддержание
массы тела, потребление
воды и
водно-солевой баланс в
организме, регуляцию
температуры в зависимости
от
температуры внешней
среды,
эмоциональных
переживаний,
мышечной работы и других
факторов, функцию
размножения.
Она включает у женщин
регулирование
менструального
цикла, вынашивание и
рождение
ребенка, кормление и
многое
другое. У мужчин —
сперматогенез,
половое поведение.
Гипоталамус играет также
центральную роль в
реакции
организма на стрессовые
воздействия.
Мне нравится
24 мая в 15:34|Ответить
Юлия Черняк
48)Функции черепных
нервов
Обонятельный нерв
( лат. nervi olfactorii) —
первый из
черепных нервов,
отвечающий
за обонятельную
чувствительность.
Зрительный
нерв (лат. Nervus
opticus) — вторая пара
черепных нервов, по
которым
зрительные раздражения,
воспринятые
чувствительными
клетками сетчатки,
передаются
в головной мозг.
Глазодвигательный нерв
(лат. nervus oculomotorius)
— III
пара черепных нервов,
отвечающий за движение
глазного яблока, поднятие
века,
реакцию зрачков на свет.
Блоковый нерв лат. nervus
trochlearis — IV пара
черепных
нервов, который
иннервирует
верхнюю косую мышцу
(лат. m.obliquus superior),
которая поворачивает
глазное
яблоко кнаружи и вниз.
Тройничный
нерв
V (тройничный) нерв
является
смешанным. По трем его
ветвям
(ramus ophthalmicus — V1,
ramus
maxillaris — V2, ramus
mandibularis — V3) через
Гасеров
узел (ganglion trigeminale)
идет
информация от верхней,
средней и нижней третей
лица
соответственно. Каждая
веточка
несет информацию от мышц,
кожных и болевых
рецепторов
каждой трети лица. В
Гасеровом
узле информация
сортируется по
типу, и уже информация от
мышц
всего лица идет в
чувствительное ядро
тройничного нерва,
расположенный большей
частью
в среднем мозге (частично
заходит в мост); кожная
информация от всего лица
идет в
«главное ядро» (nucleus
pontinus
nervi trigemini),
расположенное
в мосту; а болевая
чувствительность — в
nucleus
spinalis nervi trigemini,
идущий
от моста через
продолговатый
мозг в спинной.
Тройничному нерву
принадлежит также
двигательное ядро (лат.
nucleus
motorius nervi trigemini),
залегающее в мосте и
отвечающее за иннервацию
жевательных мышц.
Отводящий нерв (лат. nervus
abducens) — VI пара
черепных
нервов, который
иннервирует
латеральную прямую
мышцу
( лат. m. rectus lateralis) и
отвечает за отведение
глазного
яблока.
Лицевой нерв (лат. nervus
facialis), седьмой (VII) из
двенадцати черепных
нервов,
выходит из мозга между
варолиевым мостом и
продолговатым мозгом.
Лицевой
нерв иннервирует
мимические
мышцы лица. Также в
составе
лицевого нерва проходит
промежуточный нерв,
ответственный за
иннервацию
слёзной железы,
стременной
мышцы и вкусовой
чувствительности двух
передних
третей языка.
Преддверно-улитковый
нерв
(лат. nervus
vestibulocochlearis) — нерв
специальной
чувствительности
отвечающий за передачу
слуховых импульсов и
импульсов, исходящих из
вестибулярного отдела
внутреннего уха.
Языкоглоточный нерв
(лат. nervus
glossopharyngeus) —
IX пара черепных нервов.
Является смешанным.
Обеспечивает:
двигательную иннервацию
шилоглоточной мышцы
(лат. m.
stylopharyngeus),
поднимающую
глотку
иннервацию околоушной
железы (лат. glandula
parotidea)
обеспечивая её
секреторную
функцию
общую чувствительность
глотки, миндалин, мягкого
нёба,
евстахиевой трубы,
барабанной
полости
вкусовую чувствительность
задней трети языка.
Блуждающий нерв
(лат. n.vagus) — X пара
черепных
нервов. Является
смешанным.
Обеспечивает:
двигательную иннервацию
мышц мягкого нёба, глотки,
гортани, а также поперечно-
полосатых мышц пищевода
парасимпатическую
иннервацию гладких мышц
лёгких, пищевода, желудка
и
кишечника (до
селезёночного
изгиба ободочной кишки), а
также мышцы сердца. Также
влияет на секрецию желез
желудка и поджелудочной
железы
чувствительную
иннервацию
слизистой оболочки нижней
части глотки и гортани,
участка
кожи за ухом и части
наружного
слухового канала,
барабанной
перепонки и твёрдой
мозговой
оболочки задней черепной
ямки.
Дорсальное ядро
блуждающего
нерва nucleus dorsalis nervi
vagi,
располагается в
продолговатом
мозге латеральнее ядра
подъязычного нерва.
Добавочный нерв (лат.
nervus
accessorius) — XI пара
черепных
нервов. Содержит
двигательные
нервные волокна
иннервирующие мышцы
ответственные за повороты
головы, приподнимание
плеча и
приведение лопатки к
позвоночнику.
Подъязычный нерв (лат.
nervus
hypoglossus) — XII пара
черепных нервов. Отвечает
за
движение языка.
Мне нравится
24 мая в 15:36|Ответить
Юлия Черняк
49) Лимбическая система (от
лат. limbus — граница, край)
—
совокупность ряда структур
головного мозга. Участвует в
регуляции функций
внутренних
органов, обоняния,
инстинктивного поведения,
эмоций, памяти, сна,
бодрствования и др.
Функции лимбической
системы
Получая информацию о
внешней
и внутренней средах
организма,
лимбическая система
запускает
вегетативные и
соматические
реакции, обеспечивающие
адекватное приспособление
организма к внешней среде
и
сохранение гомеостаза.
Частные
функции лимбической
системы:
регуляция функции
внутренних органов (через
гипоталамус);
формирование мотиваций,
эмоций, поведенческих
реакций;
играет важную роль в
обучении;
обонятельная функция.
Включает в себя:
обонятельную луковицу
(Bulbus olfactorius)
обонятельный тракт (Tractus
olfactorius)
обонятельный треугольник
(Trigonum olfactorium)
переднее продырявленное
вещество (Substantia
perforata
anterior)
поясная извилина (Gyrus
Cinguli) (англ. Cingulate
gyrus):
автономные функции
регуляции
частоты сердцебиений и
кровяного давления;
парагиппокампальная
извилина (Gyrus
parahippocampalis)
зубчатая извилина (Gyrus
dentatus)
гиппокамп (Hippocampus):
требуемый для
формирования
долговременной памяти
миндалевидное тело (Corpus
amygdaloideum)
( англ. Amygdala): агрессия и
осторожность, страх
гипоталамус (Hypothalamus):
регулирует автономную
нервную систему через
гормоны,
голод, жажду, половое
влечение, цикл сна и
пробуждения
сосцевидное тело (Corpus
Mamillare) (англ. Mammilary
body): важен для
формирования
памяти
ретикулярную формацию
среднего мозга (Formatio
reticularis)
Мне нравится
24 мая в 15:37|Ответить
Юлия Черняк
50) Стриопаллидарная
система стала
обеспечивать фон
«предуготованности» к
совершению движения; на
этом
фоне осуществляются
контролируемые корой
головного
мозга быстрые, точные,
строго
дифференцированные
движения.
Для совершения движения
необходимо, чтобы одни
мышцы
сократились, а другие
расслабились, иначе говоря,
нужно
точное и согласованное
перераспределение
мышечного
тонуса. Такое
перераспределение
тонуса мышц как раз и
осуществляется
стриопаллидарной системой.
Эта
система обеспечивает
наиболее
экономное потребление
мышечной энергии в
процессе
выполнения движения.
Совершенствование
движения в
процессе обучения их
выполнению (например,
отработка до предела
отточенного
бега пальцев музыканта,
взмаха
руки косаря, точных
движений
водителя автомобиля)
приводит к
постепенной экономизации
и
автоматизации. Такая
возможность
обеспечивается
стриопаллидарной системой.
51) Новая кора (синонимы: неокортекс, изокортекс) (лат. neocortex) — новые области коры головного мозга, которые у низших млекопитающих только намечены, а у человека составляют основную часть коры. Новая кора располагается в верхнем слое полушарий мозга, имеет толщину 2-4 миллиметра и отвечает за высшие нервные функции — сенсорное восприятие, выполнение моторных команд, осознанное мышление и, у людей, речь. Неокортекс содержит два основных типа нейронов: пирамидальные нейроны (~80 % нейронов неокортекса) и вставочные нейроны (~20 % нейронов неокортекса).
Структура новой коры относительно однородна (отсюда альтернативное название: «изокортекс»). У человека она насчитывает шесть горизонтальных слоев нейронов, отличающихся по типу и характеру связей. Вертикально, нейроны объединены в так называемые колонки кортекса. У всех млекопитающих новая кора насчитывает 6 горизонтальных слоев нейронов.
52) Зрительный анализатор включает в себя глаз - орган зрения, воспринимающий световые раздражения,
Глаз, или глазное яблоко, имеет шаровидную форму и помещается в костной воронке - глазнице. Спереди он защищен веками. По свободному краю века растут ресницы, которые защищают глаз от попадания в него частиц пыли. У верхненаружного края глазницы расположена слезная железа, выделяющая слезную жидкость, омывающую глаз. Глазное яблоко имеет несколько оболочек, одна из которых - наружная - склера, или белочная оболочка (белого цвета). В передней части глазного яблока она переходит в прозрачную роговицу. Под белочной оболочкой расположена |
|
сосудистая оболочка, состоящая из большого количества сосудов. В переднем отделе глазного яблока сосудистая оболочка переходит в ресничное тело и радужную оболочку (радужку). В ней имеется круглое отверстие - зрачок. Здесь расположены мышцы, которые изменяют величину зрачка и, в зависимости от этого, в глаз попадает большее или меньшее количество света. Позади радужки в глазу располагается хрусталик, он имеет форму двояковыпуклой линзы. За хрусталиком полость глаза заполнена прозрачной желеобразной массой -стекловидным телом. Внутренняя поверхность глаза выстлана тонкой, сложной по строению, оболочкой - сетчаткой. Она содержит светочувствительные клетки, названные, по их форме, палочками и колбочками. Нервные волокна, отходящие от этих клеток, собираются вместе и образуют зрительный нерв. Роговица и хрусталик обладают светопреломляющей способностью Хрусталик может менять свою форму - становиться более или менее выпуклым и соответственно сильнее или слабее преломлять лучи света. Благодаря этому человек способен отчетливо видеть предметы, расположенные на разном расстоянии. |
|
зрительный нерв и зрительные центры, расположенные в коре головного мозга.
Компоненты глаза |
Особенности строения |
Роль |
Белочная оболочка (склера) |
Наружная, плотная, непрозрачная |
Защищает внутренние структуры глаза, поддерживает форму |
Роговица |
Тонкая, прозрачная |
Сильная «линза» глаза |
Конъюнктива |
Прозрачная, слизистая |
Покрывает переднюю часть глазного яблока до роговицы и внутреннюю поверхность века |
Сосудистая оболочка |
Средняя оболочка, черная, пронизана сетью кровеносных сосудов |
Питающая глаз, свет, проходя сквозь нее, не рассеивается |
Ресничное тело |
Гладкие мышцы |
Поддерживает хрусталик и изменяет его кривизну |
Радужная оболочка (радужка) |
Содержит пигмент меланин |
Светонепроницаема. Ограничивает количество света, попадающего в глаз на сетчатку. Определяет цвет глаз |
Зрачок |
Отверстие в радужной оболочке, окруженное радиальными и кольцевыми мышцами |
Регулирует количество света, попадающего на сетчатку |
Хрусталик |
Двояковыпуклая линза, прозрачное, эластичное образование |
За счет изменения кривизны фокусирует изображение |
Стекловидное тело |
Прозрачная желеобразная масса |
Заполняет внутреннюю часть глаза, поддерживает сетчатку |
Передняя камера |
Пространство между роговицей и радужкой, заполненное прозрачной жидкостью – водянистой влагой |
Участие в иммунной системе глаза |
Задняя камера |
Пространство внутри глазного яблока, ограниченное радужкой, хрусталиком и держащей его связкой, заполнено водянистой влагой |
Участие в иммунной системе глаза |
Сетчатая оболочка (сетчатка) |
Внутренняя оболочка глаза, тонкий слой клеток зрительных рецепторов: палочки (130 млн) колбочки (7 млн) |
Зрительные рецепторы формируют изображение; колбочки ответственны за цветопередачу |
Желтое пятно |
Скопление колбочек в центральной части сетчатки |
Область наибольшей остроты зрения |
Слепое пятно |
Место выхода зрительного нерва |
Месторасположение канала для передачи зрительной информации в мозг |
53) Слуховые рецепторы находятся в улитке внутреннего уха, которая расположена в пирамиде височной кости. Звуковые колебания передаются к ним через наружный слуховой проход, барабанную перепонку, слуховые косточки, жидкость лабиринта и основную перепонку улитки.
Внутри среднего канала улитки расположен звуковоспринимающий аппарат - кортиев орган, содержащий рецепторные волосковые клетки, которые трансформируют звуковые колебания в электрические изменения и возбуждение волокон слухового нерва.
При действии звуков происходит колебание эндолимфы и перилимфы, начинает колебаться основная мембрана и волоски рецепторных клеток, касаясь текториальной мембраны, деформируются. Это сопровождается электрическими изменениями в слуховом нерве.
При звуковой рецепции возникают три электрических явления: микрофонный потенциал улитки, суммарный потенциал и потенциал слухового нерва.
Если ввести в улитку электроды, соединить их с усилителем и громкоговорителем, и дать звуковое раздражение, то громкоговоритель точно воспроизводит звук. Это - микрофонный потенциал. Он генерируется поверхностью волосковой клетки, которая подобна пьезокристаллу. Для генерации микрофонного потенциала нужна энергия. Частота кохлеарных микрофонных потенциалов соответствует частоте звуковых колебаний, а амплитуда этих потенциалов в определенных границах пропорциональна интенсивности звука.
При отведении от слухового нерва регистрируются импульсы, частота которых различна в зависимости от высоты тона. Кроме того, при низких звуках импульсация наблюдается в большем числе, а при высоких - небольшом числе нервных волокон.
В улитке имеет место явление резонанса. Однако резонирующим субстратом является не определенное волокна основной мембраны, а столб жидкости определенной длины. Чем выше звук, тем меньше длина колеблющегося столба жидкости, и тем ближе к основанию улитки и овальному окошку расположено место максимальной
амплитуды колебания. При действии звуков низкой частоты длина столба растет.
Таким образом, при действии тонов разной высоты, т.е. при действии звуковых колебаний разной частоты, в улитке происходит пространственное кодирование звуковой информации. Каждой высоте тона соответствует определенная длина участка основной мембраны, охваченного колебательным процессом. Вследствие этого каждой высоте тона соответствует определенное количество рецепторов и определенная группа рецепторов, сила раздражения которых наибольшая. Интенсивность звука кодируется частотой импульсов.
54) В соматосенсорную систему включают систему кожной чувствительности и чувствительную систему скелетно-мышечного аппарата, главная роль в которой принадлежит проприорецепции.
Кожная рецепция. Кожные рецепторы. Рецепторная поверхность кожи огромна (1,4—2,1 м2). В коже сосредоточено множество рецепторов, чувствительных к прикосновению, давлению, вибрации, теплу и холоду, а также к болевым раздражениям. Их строение весьма различно (рис. 14.19). Они локализуются на разной глубине кожи и распределены неравномерно по ее поверхности. Больше всего таких рецепторов в коже пальцев рук, ладоней, подошв, губ и половых органов. У человека в коже с волосяным покровом (90 % всей кожной поверхности) основным типом рецепторов являются свободные окончания нервных волокон, идущих вдоль мелких сосудов, а также более глубоко локализованные разветвления тонких нервных волокон, оплетающих волосяную сумку. Эти окончания обеспечивают высокую чувствительность волос к прикосновению. Рецепторами прикосновения являются также осязательные мениски (диски Меркеля), образованные в нижней части эпидермиса контактом свободных нервных окончаний с модифицированными эпителиальными структурами. Их особенно много в коже пальцев рук. В коже, лишенной волосяного покрова, находят много осязательных телец (тельца Мейсснера). Они локализованы в сосочковом слое дермы пальцев рук и ног, ладонях, подошвах, губах, языке, половых органах и сосках молочных желез. Эти тельца имеют конусовидную форму, сложное внутреннее строение и покрыты капсулой. Другими инкапсулированными нервными окончаниями, но расположенными более глубоко, являются пластинчатые тельца, или тельца Фатера — Пачини (рецепторы давления и вибрации). Они есть также в сухожилиях, связках, брыжейке. В соединительнотканной основе слизистых оболочек, под эпидермисом и среди мышечных волокон языка находятся инкапсулированные нервные окончания луковиц (колбы Краузе).
55) Кожа – покровная система, представленная одним органом. Кожа покрывает все наше тело на протяжении всей жизни и подвергается множеству механических повреждений и различных факторов, поэтому главная ее функция – защитная.
Строение: -Эпидермис Состоит из наружного (роговой) и внутреннего (костистый) слоев. Внутренний слой. Клетки размножают и делятся. Наружный слой. Мертвый. Защищает и отшелушивается. Тут находится меланин, который поглащает солнечный свет и провитамин Д. -Дерма С лат. – кожа. Наука о коже – дерматология. Это сложноустроенная часть. Она образована соединительной тканью. В ней находятся потовые железы (клубочек, выводной проток), выполняющие выделительную функцию (моча, соли). Обильно снабжена кровеносными сосудами, которые выполняют транспортную и терморегуляторную функции. Здесь же находятся рецепторы и нервы, от их структур возникают импульсы, которые передаются в головной мозг по обе стороны от центральной борозды. Мы представители класса Млекопитающие, поэтому имеем сухую кожу и волосяной покров. В дерме находятся сальные железы, которые выполняют секреторную функцию (воски, они делают кожу и волосы эластичными) -Подкожно жировая клетчатка. В ней находится слой питательных веществ. Поэтому она выполняет запасающую, защитную и терморегуляторную функции.
Функции кожа: -защитная -терморегуляторная -выделительная -дыхательная -обменная -секреторная - иммунная
56) англ. olfactory system) - сенсорная система, осуществляющая анализ пахучих веществ, которые воздействуют на слизистую оболочку носовой полости. О. а. состоит из периферического отдела (обоня-гелъпые рецепторы), специфических проводящих нервных путей (обонятельный нерв и центральный обонятельный путь), подкорковых нервных структур (сосковидные тела) и коркового отдела (извилина морского коня). Периферическим отделом О. а. служат рецепторные поверхности, расположенные в слизистой оболочке верхней части носовой перегородки. Обонятельный рецептор состоит из т. н. биполярного нейрона размером 5-10 мкм и периферической части - палочкообразного Отростка (шириной ок. 1 мк и длиной 20-90 мкм), заканчивающегося обонятельным пузырьком, на котором симметрично расположены 9-16 ресничек (диаметром 0,1-0,2 мкм), непосредственно вступающих в контакт с пахучими веществами. Аксоны биполярных нейронов, собираясь в пучок обонятельного нерва, проникают через отверстия в продырявленной пластинке решетчатой кости в полость черепа и входят в обонятельную луковицу. Из митральных клеток обонятельной луковицы начинается центральный обонятельный путь, который заканчивается в височных базальных отделах мозга в области извилины морского коня. Подкорковыми нервными структурами являются т. н. сосковидные тела. Пахучие вещества проникают в слизистую оболочку носа при вдыхании через нос или через рот. Минимальная концентрация пахучего вещества, вызывающего обонятельное ощущение, называется абсолютным порогом чувствительности. О. а. человека имеет высокую чувствительность, в частности запах "искусственного мускуса" ощущается при наличии 5х1015 г его в 1 см3 воздуха. Некоторые пахучие вещества (ванилин, валерьяновая кислота) вызывают чисто обонятельные ощущения. Др. пахучие вещества вызывают наряду с обонятельными также и температурные, тактильные, болевые и вкусовые ощущения (напр., хлороформ - сладкий вкус, ментол и камфора - холод и др.). Важное свойство О. а. - адаптация (уменьшение чувствительности) к длительному раздражению пахучим веществом. Одновременное действие нескольких пахучих веществ приводит к их смешению. В некоторых случаях происходит подавление одного запаха др. Возможны нейтрализация запахов, когда смесь не вызывает обонятельного ощущения; появление нового запаха; последовательная смена запахов; увеличение чувствительности к одному запаху после действия другого и др. явления, возникающие при смешении запахов.
57) Чувство вкуса связано с раздражением не только химических, но и механических, температурных и даже болевых рецепторов слизистой оболочки полости рта, а также обонятельных рецепторов. Вкусовой анализатор определяет формирование вкусовых ощущений, является рефлексогенной зоной. С помощью вкусового анализатора оцениваются различные качества вкусовых ощущений, сила ощущений, которая зависит не только от силы раздражения, но и от функционального состояния организма.
Структурно-функциональная характеристика вкусового анализатора.
Периферический отдел. Рецепторы вкуса (вкусовые клетки с микроворсинками) — это вторичные рецепторы, они являются элементом вкусовых почек, в состав которых входят также опорные и базальные клетки. Во вкусовых почках обнаружены клетки, содержащие серотонин, и клетки, образующие гистамин. Эти и другие вещества играют определенную роль в формировании чувства вкуса. Отдельные вкусовые почки являются полимодальными образованиями, так как могут воспринимать различные виды вкусовых раздражителей. Вкусовые почки в виде отдельных включений находятся на задней стенке глотки, мягком нёбе, миндалинах, гортани, надгортаннике и входят также в состав вкусовых сосочков языка как органа вкуса.
Периферический отдел вкусового анализатора представлен вкусовыми луковицами, которые расположены главным образом в сосочках языка. Вкусовые клетки усеяны на своем конце микроворсинками, которые называют еще вкусовыми волосками. Они выходят на поверхность языка через вкусовые поры.
На вкусовой клетке имеется большое число синапсов, которые образуют волокна барабанной струны и языкоглоточного нерва. Волокна барабанной струны (ветвь язычного нерва) подходят ко всем грибовидным сосочкам, а волокна языкоглоточного нерва — к желобоватым и листовидным. Корковый конец вкусового анализатора находится в гиппокампе, парагиппокамповой извилине и в нижней части заднецентральной извилины.
Вкусовые клетки непрерывно делятся и непрерывно гибнут. Особенно быстро происходит замещение клеток, расположенных в передней части языка, где они лежат более поверхностно. Замена клеток вкусовой почки сопровождается образованием новых синаптических структур
Проводниковый отдел. Внутрь вкусовой почки входят нервные волокна, которые образуют рецепторно-афферентные синапсы. Вкусовые почки различных областей полости рта получают нервные волокна от разных нервов: вкусовые почки передних двух третей языка — от барабанной струны, входящей в состав лицевого нерва; почки задней трети языка, а также мягкого и твердого нёба, миндалин — от языкоглсточного нерва; вкусовые почки, расположенные в области глотки, надгортанника и гортани, — от верх-пегортанного нерва, являющегося частью блуждающего нерва.
Эти нервные волокна являются периферическими отростками биполярных нейронов, расположенных в соответствующих чувствительных ганглиях, представляющих первый нейрон проводникового отдела вкусового анализатора. Центральные отростки этих клеток входят в состав одиночного пучка продолговатого мозга, ядра которого представляют второй нейрон. Отсюда нервные волокна в составе медиальной петли подходят к зрительному бугру (третий нейрон).
Центральный отдел. Отростки нейронов таламуса идут в кору больших полушарий (четвертый нейрон). Центральный, или корковый, отдел вкусового анализатора локализуется в нижней части соматосенсорной зоны коры в области представительства языка. Большая часть нейронов этой области мультимодальна, т. е. реагирует не только на вкусовые, но и на температурные, механические и ноцицептивные раздражители. Для вкусовой сенсорной системы характерно то, что каждая вкусовая почка имеет не только афферентные, но и эфферентные нервные волокна, которые подходят к вкусовым клеткам из ЦНС, благодаря чему обеспечивается включение вкусового анализатора в целостную деятельность организма.
58) Ноцицептивная чувствительность (noceo – повреждаю + receptivus – восприимчивый) – чувствительность к действию раздражителя, вызывающего в организме ощущение боли. Полагают, что возникающая в ответ на раздражение боль как комплексная функция в наиболее полной мере свойственна только организму человека. У животных также возникают подобные процессы, но они не идентичны тем, которые наблюдаются у человека. Раздражение воспринимается как экстеро-Боль - психофизиологическая реакция животных и человека на повреждающий раздражитель, вызывающий в организме органические или функциональные нарушения. Важнейший компонент боли - субъективные ощущения, носящие характер страдания. Боль - врожденная сигнальная реакция, но в течение жизни условнорефлекторные компоненты могут облегчать или усиливать ее.
Принято рассматривать боль как нейрофизиологический феномен, имеющий периферический и центральный механизмы, причем последние играют ведущую роль в формировании боли. С развитием электрофизиологических методов было установлено, что кроме проведения возбуждения спинной мозг выполняет функции модулятора афферентных возбуждений, в частности болевых. Особую роль при этом играют клетки так называемой желатинозной субстанции, находящейся в боковых рогах спинного мозга.
 значение
то, что возникают разные синдромы.
значение
то, что возникают разные синдромы.
59) электроэнцефалография (ЭЭГ) - метод исследования деятельности головного мозга животных и человека; основан на суммарной регистрации биоэлектрической активности отдельных зон, областей, долей мозга. ЭЭГ применяется в современной нейрофизиологии, а также в неврологии и психиатрии.
|
|
|
|
Дельта (delta) ритм – колебания частотой менее 3.5 Гц (Walter, 1936). Нижней границей дельта в клинической ЭЭГ является, как правило, 0.3-0.5 Гц. В зависимости от уровня бодрствования и локализации дельта колебания свидетельствуют либо о локальной/диффузной церебральной дисфункции (опухоль, кровоизлияние, кома, и др.), либо отражают физиологическое снижение уровня активации головного мозга (сон). Отдельная медленная волна может являться также компонентом патологических комплексов, например, эпилептиформного паттерна спайк-волна, и др. Тета (theta) ритм – 4-7.5 Гц (Walter and Dovey, 1944). Так же как и дельта, колебания тета-диапазона могут отражать локальную или диффузную церебральную дисфункцию, или физиологически обусловленное снижение уровня бодрствования. Альфа (alpha) – 8 -13 Гц, доминирует в задних отделах, у взрослых имеет в среднем амплитуду до 70-80 микровольт, блокируется при открывании глаз, связан со зрительными структурами мозга (Berger, 1929). Альфа-ритм формируется в первые месяцы и годы жизни, имеет вначале частоту 3-4 Гц, постепенно достигая таких же значений, как и у взрослых, к возрасту 3-х лет. Это первый описанный и наиболее изученный ритм, наиболее выражен в нормальной ЭЭГ расслабленного бодрствования, с его описания обычно начинается составление заключения ЭЭГ. Бета (beta) ритм – колебания от 14 Гц и выше. На практике верхней границей бета активности обычно являются значения 30-35 Гц. Частоты выше 30 Гц иногда описываются как гамма ритм. В норме у взрослого человека в состоянии расслабленного бодрствования амплитуда бета-ритма обычно значительно ниже, чем амплитуда альфа, чаще менее 20-30 микровольт, с тенденцией к максимуму в передних отделах. Доминирование бета-активности может являться следствием воздействия различных медикаментов. Следует различать собственно бета активность мозгового происхождения от электромиографических артефактов, которые часто имеют такую же частоту. |
|
|
Другие ритмы и колебания, которые могут описываться в клинической практике. |
|
|
Мю (mu) ритм – та же частота, что и альфа, но имеет другую пространственную организацию (максимум в центральных отделах), форму, реактивность, блокируется при произвольных движениях, связан с сенсомоторной корой (Jasper and Andrews, 1938). Лямбда (lambda) волны – колебания, возникающие в задних отделах при следящих движениях глаз (Gastaut, 1951; Evans, 1952). |
|
|
Ритмы, которые в основном описываются в научной литературе, использование которых необязательно в клинической практике |
|
|
Каппа (kappa) – колебания в альфа-диапазоне с максимумом в передне-височных отделах (Laugier and Liberson, 1937) Пи (pi) - медленные колебания (3-4 Гц) в задних отделах мозга (Dutertre, 1977), отличающиеся от основного затылочного ритма и не являющиеся его гармоникой. Фи (phi) – медленные колебания дельта диапазона в задних отделах, характерно возникающие при закрытии глаз (Belsh et al., 1983) Сигма (sigma) – аналогично «сонным веретенам» (Kugler, 1981) Ро (rho) - аналогично позитивным затылочным острым компонентам (POSTS) (Kugler, Laub, 1973). |
|
Клиническое значение. Запись ЭЭГ - диагностическая процедура, которая используется в неврологической практике. Так, при диффузных органических повреждениях головного мозга, черепно-мозговых травмах наблюдаются замедленные и нерегулярные волны. При опухолях мозга часто возникают местные изменения ЭЭГ (в области опухолей). У больных эпилепсией на ЭЭГ наблюдаются пароксизмальные потенциалы, судорожные разряды, остроконечные волны и другие изменения. Запись ЭЭГ широко используется в хирургической практике для контроля глубины наркоза: при глубокой стадии наркотического сна на ЭЭГ преобладают дельта-волны. При констатации смерти в сомнительных случаях, особенно при реанимации больного, часто ориентируются на исчезновение колебаний на ЭЭГ («плоская» ЭЭГ). В клинической практике также используют метод регистрации вызванных потенциалов для получения объективных данных о характере и динамике некоторых нарушений сенсорных функций. Приведенные данные об общих закономерностях деятельности ЦНС имеют не только теоретическое значение, но используются и в медицинской практике. Так, раскрытие механизмов возникновения и проведения ПД - основной формы активной реакции возбудимой клетки, хеморецептивних свойств клеточной мембраны и ее ионных каналов, механизма межклеточной передачи возбуждения легло в основу разработки новых методов диагностики и лечения многих заболеваний в психоневрологической, кардиологической и других областях клинической медицины.
60) Принципы реоэнцефалографического исследования Метод современной диагностики с помощью реоэнцефалографии осуществляется двумя приборами – записывающим устройством и реографической приставкой. Оборудование позволяет оценить кровообращение в сосудах головного мозга. В процессе проведения реоэнцефалографии регистрируются пульсовые колебания в сосудах головного мозга и оценивается скорость кровотока, а также проходимость и состояние тонуса сосудов определенного участка мозга. Пациент при проведении реоэнцефалографии лежит на спине или на животе (в зависимости от вида диагностики) неподвижно, закрывает глаза и старается не реагировать на внешние раздражители. Специалист по диагностике проводит реоэнцефалографию, оценивая следующие показатели: - регулярность импульсных волн, - конфигурацию и наклон нисходящей и восходящей частей, - положение инцизуры, - наличие и распределение венозной волны, - дополнительные импульсные волны. Результаты диагностики обладают высокой степенью точности и позволяют раскрыть полную клиническую картину течения определенного заболевания сосудов головного мозга.
УЗИ головного мозга, или нейросонография – это метод исследования мозга и других структур, расположенных в полости черепа, с помощью ультразвука. Обычно УЗИ головного мозга проводится у детей с открытым родничком или швами, через которые ультразвук может проникать в полость черепа. Нейросонографию проводят для определения состояние головного мозга, размеров его отдельных частей, наличия некоторых дефектов развития мозга или патологических образований (гематомы, кисты и пр.). УЗИ – это абсолютно безопасный метод исследования, который не имеет противопоказаний и побочных эффектов.
Допплерография - это ультразвуковой метод исследования, основанный на изменении частоты ультразвуковых волн, отраженных от подвижных структур (эффекте Допплера).
Показания к допплерографии
Допплерография позволяет регистрировать и оценивать параметры кровотока, состояние различных сосудов (сужения, неровности и др.), скорость движения мышечных стенок и клапанов сердца. Причем, в отличие от ангиографического исследования сосудов, допплерография является совершенно безвредным для больного видом исследования - нет связи с какими-либо манипуляциями, обусловленными введением контрастных веществ в кровоток. Особенно очевидны преимущества этого исследования в акушерской практике. В связи с разработкой и внедрением допплерографии появилась возможность измерения скорости кровотока в сосудах плода, пуповины и маточных артерий, а также регистрации движений сердца плода, тогда как при ультразвуковом сканировании проводятся лишь визуальная оценка и измерение размеров матки, плаценты, околоплодных вод, плода и его органов. Допплерография является обязательным компонентом в комплексе обследования беременных. Первое исследование проводится на сроке 16-20 недель беременности, так как в этот период могут быть обнаружены патологические типы допплерог-рамм в артерии пуповины и ее мелких кровеносных сосудах, а также в маточной артерии, что имеет высокую прогностическую ценность в профилактике развития токсикозов (отеков, нефропатии, преэклампсии, эклампсии) второй половины беременности и сопровождающих их внутриутробной гипоксии и гипотрофии плода.