

Біосинтез вуглеводів
Біосинтез глюкози й інших вуглеводів із простіших попередників у кількісному відношенні є найбільш важливим процесом у біосфері. Фотосинтезуючі (автотрофні) організми утворюють велику кількість гексоз із вуглекислого газу і води, а в клітинах гетеротрофних організмів центральне місце в обміні вуглеводів займає піровиноградна і молочна кислоти, амінокислоти та інші прості попередники глюкози і глікогену.
Перетворення глюкози у піровиноградну кислоту, що каталізується ферментами гліколізу, є центральним шляхом катаболізму вуглеводів у більшості клітин як в аеробних, так і в анаеробних умовах. Точно такий самий оборотний процес – перетворення пірувату у глюкозу – є найважливішим шляхом біосинтезу моно- і полісахаридів.
Центральний біосинтетичний шлях складається із двох головних шляхів, що починаються з двох різних наборів невуглеводних попередників.
Один з них складається з ряду реакцій, завдяки яким проміжні продукти циклу трикарбонових кислот перетворюються у піровиноградну кислоту. Процес проходить в 0усіх організмах і називається глюконеогенезом.
Другий важливий шлях складається з реакцій, які ведуть до відновлення СО2 в глюкозу. Цей шлях характерний тільки для автотрофних організмів.
З глюкозо-6-фосфату можуть утворюватися різні вуглеводи:
вільна глюкоза;
запасні полімери – крохмаль і глікоген;
різні моносахариди та їх похідні;
дисахариди й олігосахариди;
компоненти клітинної стінки й оболонки, такі як целюлози, ксилани, муреїни, кислі мукополісахариди.
1. Утворення глюкози з пірувату.
Більшість стадій біосинтетичного шляху утворення глюкози із пірувату каталізується ферментами гліколітичного циклу, отже вони є оборотними реакціями гліколізу. Але в нормальному гліколітичному шляху є три необоротні стадії, які використати при перетворенні пірувату в глюкозу не можна. При біосинтезі глюкози процес проходить в обхід цих стадій.
Перша з них – перетворення пірувату в фосфоенолпіруват відбувається за рахунок кількох реакцій, що каталізуються як ферментами цитоплазми, так і ферментами мітохондрій. У цій послідовності перша реакція каталізується ферментом – мітохондріальною піруваткарбоксилазою за схемою:
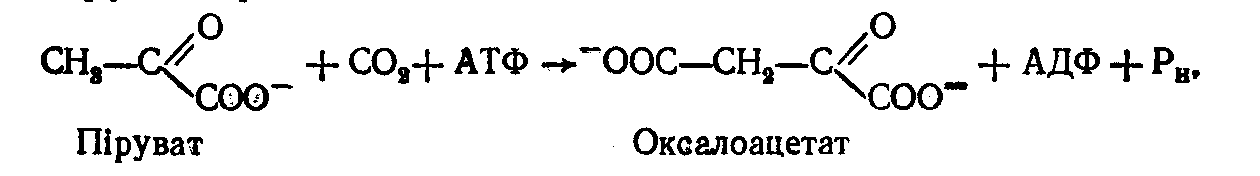
Піруваткарбоксилаза є регуляторним ферментом, який повністю неактивний у відсутності ацетил-КоА. Оксалоацетат (щавелевооцтова кислота) потім відновлюється в мітохондріях у малат (яблучна кислота):
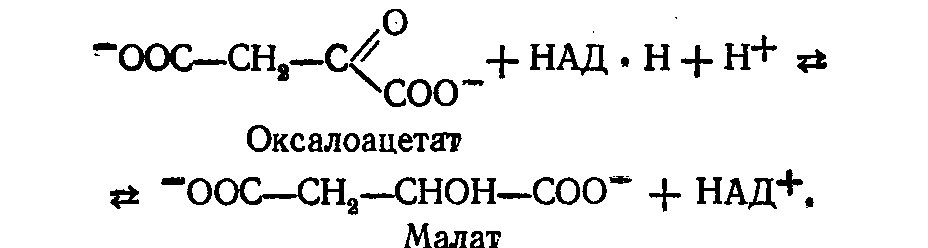
Потім малат дифундує з мітохондрій у цитоплазму, де він окислюється цитоплазматичною НАД-залежною малатдегідрогеназою з утворенням позамітохондріального оксалоацетату:
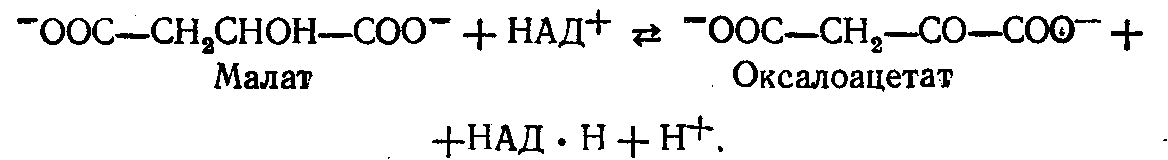
Під дією ферменту фосфоенолпіруваткарбоксилази із оксалоацетату (щавелевооцтова кислота) утворюється фосфоенолпіруват; донором фосфату в цій реакції є ГТФ:
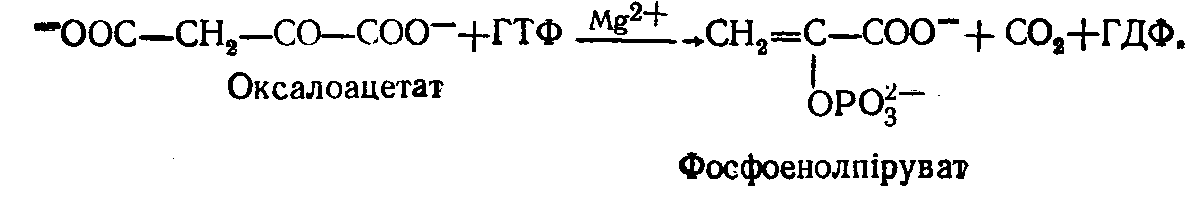
На фосфорилювання однієї молекули пірувату використовується енергія двох макроергічних фосфатних зв'язків – АТФ і ГТФ

Фосфоенолпіруват далі легко перетворюється у фруктозо-1,6-дифосфат за рахунок оборотних реакцій гліколізу. Фруктозо-1,6-дифосфат у гліколітичному циклі утворюється за участю ферменту фосфофруктокінази, і ця реакція необоротна. Біосинтез глюкози проходить в обхід цієї реакції за допомогою ферменту фруктозодифосфатази (гексозодифосфатази), що каталізує гідроліз першої фосфатної групи.
На наступній (оборотній) стадії біосинтезу фруктозо-6-фосфат перетворюється у глюкозо-6-фосфат за участю ферменту фосфогексозоізомерази.
У більшості клітин глюкозо-6-фосфат використовується як попередник при біосинтезі запасних полі-, ди-і моносахаридів. Але в клітинах печінки, нирок, епітелії кишок хребетних глюкозо-6-фосфат може дефосфорилюватися глюкозо-6-фосфатазою з утворенням вільної глюкози. Гексокіназна реакція, яка приводить до утворення глюкозо-6-фосфату, є необоротною.
Сумарне рівняння утворення глюкози з пірувату має такий вигляд:
![]()
Отже, на утворення однієї молекули глюкози з пірувату використовується шість високоенергетичних фосфатних зв'язків і дві молекули НАДН2 для відновлення.
2. Біосинтез дисахаридів – здійснюється за реакціями трансглікозування. При цьому процес перенесення глікозильного залишку на один моносахарид проходить з фосфорного ефіру другого моносахариду. Реакція каталізується глікозилтрансферазою.
Біосинтез дисахаридів відбувається за участю нуклеозиддифосфатсахарів. Так, у рослин сахароза утворюється за участю серії реплікацій:
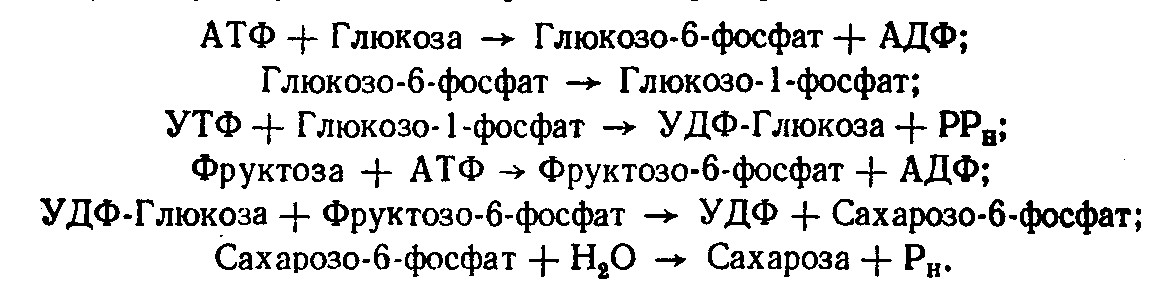
С![]() умарне
рівняння цього процесу таке:
умарне
рівняння цього процесу таке:
Дисахарид лактоза утворюється з D-глюкози і УДФ-галактози під дією лактатсинтетазної системи. У свою чергу D-глікозильний залишок перетворюється у D-галактозильний через нуклеозиддифосфатгексози, утворення яких каталізується пірофосфорилазами. Перетворення D-глікозильного залишку в D-галактозильний відбувається шляхом ферментативної епімеризації УДФ-глюкози при четвертому вуглецевому атомі залишку глюкози. Каталізує цю реакцію фермент УДФ-глюкозоепімераза, який має НАД у своєму складі.
D-галактоза спочатку фосфорилюється за участю ферменту галактокінази з утворенням D-галактозо-1-фосфату:
![]()
Далі галактозо-1-фосфат перетворюється у УДФ-галактозу або за допомогою реакції, яку каталізує фермент гексозо-1 -фосфат- уридилтрансфераза:
![]()
або за допомогою ферменту УДФ-галактопірофосфорилази
![]()
Отже, у біосинтезі і перетворенні основної кількості вуглеводів беруть участь нуклеозиддифосфатсахари, що виконують коферментні функції.
1 Фумарат може акцептувати електрони, які поставляються коферментами, що переносять водень, і вже прошли частину шляху по дихальному ланцюгу; тому він робить можливим окислювальне фосфорилювання. Такого роду фосфорилювання з фумаратом в якості кінцевого акцептора електронів можна віднести до «анаеробних дихальних процесів» і говорити про фумаратне дихання.