

10.5. Планирование будущих действий и вторичные моторные области
П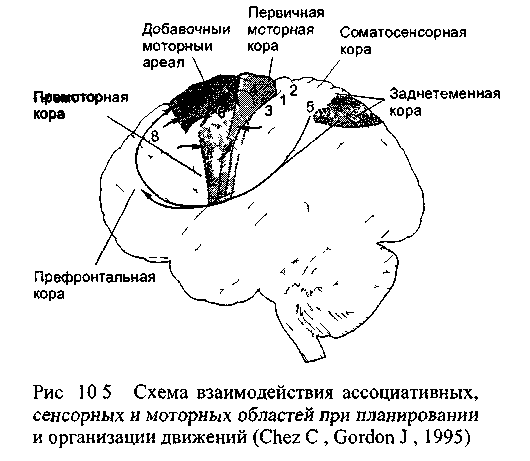 ланировать
предстоящие действия начинает
дорсолатеральная префронтальная
ассоциативная кора на основе информации,
поступающей, в первую очередь, от
заднетеменных областей, с которыми ее
связывает множество нейронных путей
(рис 10 5) Полученная информация содержит
карту находящихся в поле зрения предметов,
эту карту префронтальная кора включает
в свою рабочую память Эксперименты на
обезьянах, у которых повреждали
префронталь-ную область коры, показали
нарушенную способность узнавать позицию
необходимых для деятельности объектов
и серьезное ухудшение рабочей памяти
Так, например, на глазах обезьяны
накрывали орех одним из трех
перевернутых непрозрачных сосудов, а
затем всего лишь на несколько секунд
загораживали все эти предметы непрозрачной
ширмой Если нормальные обезьяны после
открытия ширмы сразу же брали сосуд,
под которым находился орех, то животные
с повреждением префронтальной коры,
начинали беспорядочно перевертывать
сосуды в поисках ореха, как будто не
видели, где он был спрятан. Выходная
активность префронтальной ассоциативной
коры адресована премоторным или
вторичным моторным областям, которые
создают конкретный план предстоящих
действий и непосредственно готовят
моторные системы к движению Ко вторичным
двигательным областям относятся
премоторная кора и добавочный моторный
ареал (поле 6), причем обе эти области
организованы сома-тотопически Впервые
они появляются у приматов и совершенствуются
в процессе дальнейшей эволюции их
относительная величина у человека
примерно в шесть раз больше, чем у макаки
Премоторная кора находится
непосредственно перед первичной моторной
корой, а добавочный моторный ареал - на
верхней стороне лобных долей у медиальных
поверхностей продольной борозды Недавно
были открыты еще две области вторичной
моторной коры, расположенные в поясных
извилинах вентральней от добавочных
ареалов Выходная активность нейронов
вторичной моторной коры направлена
к первичной моторной коре и к подкорковым
структурам, кроме того некоторые
аксоны ее нейронов входят в состав
волокон кор-тикоспинального пути,
спускающегося в спинной мозг
ланировать
предстоящие действия начинает
дорсолатеральная префронтальная
ассоциативная кора на основе информации,
поступающей, в первую очередь, от
заднетеменных областей, с которыми ее
связывает множество нейронных путей
(рис 10 5) Полученная информация содержит
карту находящихся в поле зрения предметов,
эту карту префронтальная кора включает
в свою рабочую память Эксперименты на
обезьянах, у которых повреждали
префронталь-ную область коры, показали
нарушенную способность узнавать позицию
необходимых для деятельности объектов
и серьезное ухудшение рабочей памяти
Так, например, на глазах обезьяны
накрывали орех одним из трех
перевернутых непрозрачных сосудов, а
затем всего лишь на несколько секунд
загораживали все эти предметы непрозрачной
ширмой Если нормальные обезьяны после
открытия ширмы сразу же брали сосуд,
под которым находился орех, то животные
с повреждением префронтальной коры,
начинали беспорядочно перевертывать
сосуды в поисках ореха, как будто не
видели, где он был спрятан. Выходная
активность префронтальной ассоциативной
коры адресована премоторным или
вторичным моторным областям, которые
создают конкретный план предстоящих
действий и непосредственно готовят
моторные системы к движению Ко вторичным
двигательным областям относятся
премоторная кора и добавочный моторный
ареал (поле 6), причем обе эти области
организованы сома-тотопически Впервые
они появляются у приматов и совершенствуются
в процессе дальнейшей эволюции их
относительная величина у человека
примерно в шесть раз больше, чем у макаки
Премоторная кора находится
непосредственно перед первичной моторной
корой, а добавочный моторный ареал - на
верхней стороне лобных долей у медиальных
поверхностей продольной борозды Недавно
были открыты еще две области вторичной
моторной коры, расположенные в поясных
извилинах вентральней от добавочных
ареалов Выходная активность нейронов
вторичной моторной коры направлена
к первичной моторной коре и к подкорковым
структурам, кроме того некоторые
аксоны ее нейронов входят в состав
волокон кор-тикоспинального пути,
спускающегося в спинной мозг
Премоторная область получает переработанную сенсорную информацию от задне-теменных областей; она содержит, прежде всего, сведения о комплексе зрительных и соматосенсорных ощущений. Выходы из премоторной области, направленные к стволу, включаются в состав медиального пути, в основном ретикулоспинального тракта. С этим обстоятельством связана главная задача премоторной области коры: контролировать мышцы туловища и проксимальных отделов конечностей. Эти мышцы особенно важны в начальной фазе выпрямления тела или движения руки к намеченной цели.
В отличие от этого, добавочный моторный ареал программирует последовательность движений, которые выполняются билатерально: при повреждении этой области коры у обезьян отдельные движения могут выполняться правильно, но координировать их в общем потоке действий животные уже не могут так, как прежде, и в особенности в тех случаях, когда надо действовать обеими руками. Добавочная кора получает информацию главным образом от соматосенсор-ной коры, а выход из неё происходит двумя способами: для проксимальных мышц конечностей двигательные команды поступают непосредственно в спинной мозг, а для дистальных мышц - опосредованно через первичную моторную кору.
Сравнительно недавно участие разных областей моторной коры в организации движения у человека было исследовано в зависимости от интенсивности кровотока в этих областях: известно, что с повышением активности ЦНС кровоток усиливается. Для оценки кровотока в сонную артерию вводили помеченный короткоживущим радиоизотопом ксенон (Roland P. E., 1993): распределение этого инертного газа в крови зависит только от интенсивности кровотока. Испытуемые выполняли серию стандартных движений пальцами (при этом активными оказались префронтальная область, добавочный моторный ареал, область руки в первичной моторной коре и в соматосен-сорной коре), затем не двигались, но представляли себе эти же движения (активен добавочный моторный ареал), затем делали несколько энергичных взмахов одним пальцем (активны области руки первичной моторной и сенсорной коры) и, наконец, при закрытых глазах по команде изменяли положение пальца в ячейках проволочной решётки («два поля направо», «одно поле вперёд» и т.п.) - при этом тесте повышалась активность кровотока задне-теменной и префронтальной областей, добавочного моторного ареала, премоторной коры, первичной моторной и сенсорной коры.
На основании сопоставления характера кровотока при выполнении этих опытов, а также в сравнении с кровотоком в состоянии покоя и при закрытых глазах были сделаны следующие выводы об участии различных областей коры в организации движений. Задне-те-менные области обеспечивают сенсорной информацией префронталь-ную и вторичную моторную кору. Добавочный моторный ареал участвует в создании модели двигательной программы и её исполнении. Премоторная кора модифицирует созданную программу на основе поступающей сенсорной информации. Первичная моторная кора обеспечивает простые движения, причём даже без участия ассоциативной или вторичной коры. Эти выводы в общем совпадают с представлениями о роли разных областей коры в движении, сложившимися прежде на основе многолетних исследований двигательного поведения обезьян, изменённого в результате повреждений различных регионов коры.