

7.4. Нервные центры
В соответствии с классической традицией представление о нервных центрах рефлексов составляет сердцевину всей рефлекторной теории. Под нервным центром понимают функциональное объединение интернейронов, участвующих в осуществлении рефлекторного акта. Они возбуждаются притоком афферентной информации и адресуют свою выходную активность эфферентным нейронам. Несмотря на то, что нервные центры тех или иных рефлексов находятся в определённых структурах мозга, например в спинном, продолговатом, среднем и т.д., их принято считать функциональными, а не анатомическими объединениями нейронов. Дело в том, что многие интернейроны способны участвовать в замыкании не одной, а нескольких рефлекторных дуг, т.е. они могут поочерёдно входить в состав то одного, то другого центра.
Сформулировавший классические принципы рефлекторной теории Чарлз Шеррингтон (Sherrington С. S.) не был склонен их абсолютизировать, что видно хотя бы из следующей цитаты: «Возможно, «простой рефлекс» является чисто абстрактным понятием, так как все части нервной системы связаны воедино и, вероятно, ни одна из них не в состоянии участвовать в какой-либо реакции, не воздействуя и не испытывая воздействия со стороны других частей, причём вся система, несомненно, никогда не находится в состоянии полного покоя. Однако понятие «простая рефлекторная реакция» оправдано, хотя и несколько проблематично».
Центры спинномозговых двигательных рефлексов испытывают влияние двигательных центров ствола мозга, которые, в свою очередь, подчиняются командам нейронов, входящих в состав ядер мозжечка, подкорковых ядер, а также пирамидным нейронам моторной коры. На каждом иерархическом уровне существуют локальные сети нейронов, по которым возбуждение может циркулировать, таким образом сохраняя информацию в пределах этого уровня. Нейроны разных уровней контактируют друг с другом, оказывая возбуждающее или тормозное действие. За счёт конвергенции и дивергенции в процесс переработки информации вовлекается дополнительное количество нейронов, что повышает надёжность функционирования иерархически организованных центров.
Свойства центров целиком определяются деятельностью центральных синапсов. Именно поэтому возбуждение через центр передаётся только в одном направлении и с синаптической задержкой. В центрах происходит пространственная и последовательная суммация возбуждения, здесь возможно усиление сигналов и трансформация их ритма. Феномен посттетанической потенциации демонстрирует пластичность синапсов, их способность изменять эффективность передачи сигналов.
7.5. Рефлексы растяжения - простая модель стереотипной реакции
Шеррингтон изучал эти рефлексы на собаках, мозг которых перерезался на разных уровнях: например, между продолговатым мозгом и спинным или между верхними и нижними буграми четверохолмия. С помощью таких экспериментальных моделей удалось детально изучить многие двигательные рефлексы спинного мозга и обнаружить принцип субординации в отношениях между спинным и головным мозгом.
Известно, что каждое движение требует координированных действий нескольких мышц: например, для того, чтобы взять карандаш в руку потребуется участие около дюжины мышц, из которых одни должны сокращаться, а другие - расслабляться. Совместно действующие мышцы, т.е. сокращающиеся или расслабляющиеся одновременно называются синергистами, в отличие от противодействующих им мышц-антагонистов. При любом двигательном рефлексе сокращения и расслабления синергистов и антагонистов безупречно координированы друг с другом.
По каким правилам взаимодействуют нейроны, управляющие сокращающимися и расслабляющимися мышцами? Рассмотрим самый простой случай - рефлекс растяжения, впервые обнаруженный Шеррингтоном у собак с перерезанным на уровне среднего мозга стволом. У таких животных возникает т.н. децеребрационная ригидность (лат. rigiditas - жёсткость, оцепенение), которая проявляется резким повышении тонуса всех мышц-разгибателей, поэтому лапы максимально разогнуты, а спина и хвост прогибаются дугой. В норме тонус разгибателей и сгибателей уравновешивается двигательными ядрами ствола мозга, а после перерезки ствола красные ядра среднего мозга, поддерживающие тонус сгибателей, отделяются от спинного мозга и на этом фоне наблюдается стимулирующее влияние вестибулярных ядер на разгибатели. При попытке согнуть лапу такой собаки, а значит растянуть находящиеся в тоническом сокращении мышцы-разгибатели, исследователь обнаруживает в ответ рефлекторно возникающее сопротивление и дополнительное сокращение мышц. При этом выявляются два компонента рефлекса: 1) сначала сильный кратковременный фазный - в ответ на изменение длины мышцы, т.е. в самый момент сгибания и 2) слабый продолжительный тонический - когда принудительно согнутой лапе не позволяют разогнуться, удерживая растянутое состояние мышцы, т.е. её новую длину.
Р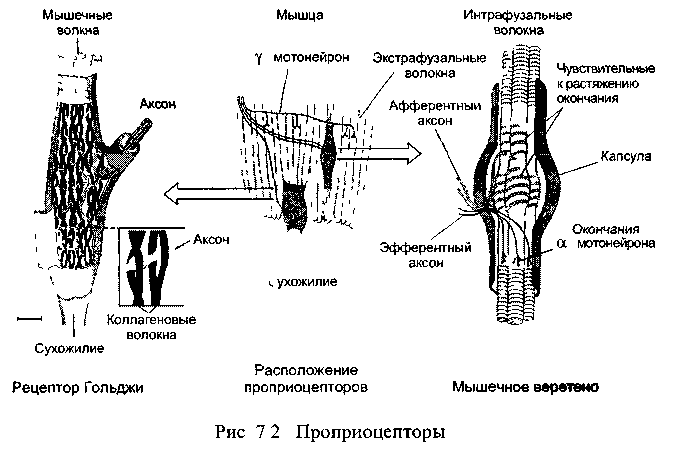 ефлексы
растяжения можно обнаружить и у интактных
животных, однако они оказываются
слабее, чем у децеребрированных, а их
стереотипность будет выражена меньше,
что обусловлено характером активирующих
и тормозных влияний двигательных центров
головного мозга. Как впоследствии стало
известно, в ответ на растяжение мышцы
внешней силой возбуждаются реагирующие
только на изменение длины рецепторы
мышечных веретён (рис. 7.2), которые
связаны с особым типом мелких интрафузальных
(от лат. fusus
-веретено) мышечных волокон. От этих
рецепторов возбуждение передаётся по
чувствительному нейрону в спинной мозг,
где окончание аксона разделяется на
несколько ветвей. Одни ветви аксона
образуют синапсы с мотонейронами
мышц-разгибателей и возбуждают их,
что, естественно, приводит к сокращению
мышц: здесь моно-синаптический рефлекс
- его дуга образована лишь двумя нейронами
Одновременно с этим остальные ветви
афферентного аксона активируют
деятельность тормозных интернейронов
спинного мозга, которые тотчас подавляют
активность мотонейронов для
мышц-антагонистов, т.е сгибателей.
Таким образом, растяжение мышцы вызывает
возбуждение мотонейронов мышц-синергистов
и реципрокно тормозит мотонейроны
мышц-антагонистов (рис. 7.3).
ефлексы
растяжения можно обнаружить и у интактных
животных, однако они оказываются
слабее, чем у децеребрированных, а их
стереотипность будет выражена меньше,
что обусловлено характером активирующих
и тормозных влияний двигательных центров
головного мозга. Как впоследствии стало
известно, в ответ на растяжение мышцы
внешней силой возбуждаются реагирующие
только на изменение длины рецепторы
мышечных веретён (рис. 7.2), которые
связаны с особым типом мелких интрафузальных
(от лат. fusus
-веретено) мышечных волокон. От этих
рецепторов возбуждение передаётся по
чувствительному нейрону в спинной мозг,
где окончание аксона разделяется на
несколько ветвей. Одни ветви аксона
образуют синапсы с мотонейронами
мышц-разгибателей и возбуждают их,
что, естественно, приводит к сокращению
мышц: здесь моно-синаптический рефлекс
- его дуга образована лишь двумя нейронами
Одновременно с этим остальные ветви
афферентного аксона активируют
деятельность тормозных интернейронов
спинного мозга, которые тотчас подавляют
активность мотонейронов для
мышц-антагонистов, т.е сгибателей.
Таким образом, растяжение мышцы вызывает
возбуждение мотонейронов мышц-синергистов
и реципрокно тормозит мотонейроны
мышц-антагонистов (рис. 7.3).
С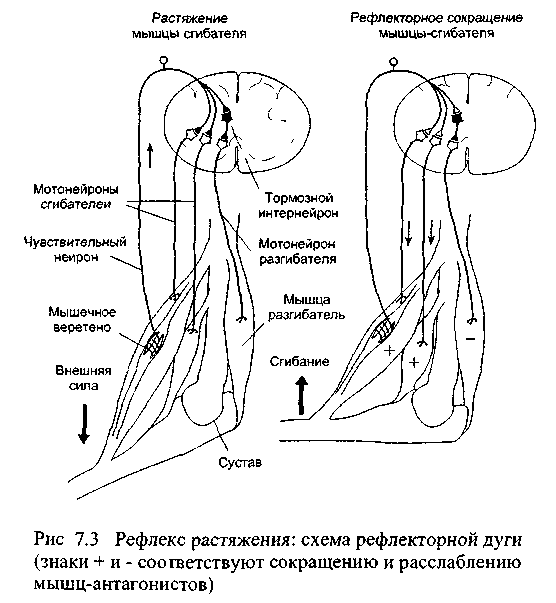 илу,
с которой мышцы сопротивляются изменению
своей длины, можно определить как
мышечный тонус. Он позволяет сохранять
определенное положение тела или позу.
Сила гравитации направлена на растяжение
мышц-разгибателей, а их ответное
рефлекторное сокращение этому
противодействует. Если растяжение
разгибателей увеличивается, например,
когда на плечи опускается тяжёлый
груз, то и сокращение усиливается - мышцы
не позволяют себя растягивать и благодаря
этому сохраняется поза. При отклонениях
тела вперёд, назад или в сторону
определённые мышцы растягиваются,
а рефлекторное повышение их тонуса
сохраняет необходимое положение
тела.
илу,
с которой мышцы сопротивляются изменению
своей длины, можно определить как
мышечный тонус. Он позволяет сохранять
определенное положение тела или позу.
Сила гравитации направлена на растяжение
мышц-разгибателей, а их ответное
рефлекторное сокращение этому
противодействует. Если растяжение
разгибателей увеличивается, например,
когда на плечи опускается тяжёлый
груз, то и сокращение усиливается - мышцы
не позволяют себя растягивать и благодаря
этому сохраняется поза. При отклонениях
тела вперёд, назад или в сторону
определённые мышцы растягиваются,
а рефлекторное повышение их тонуса
сохраняет необходимое положение
тела.
По тому же принципу осуществляется рефлекторная регуляция длины у мышц-сгибателей. При любом сгибании руки или ноги поднимается груз, которым может быть и сама рука или нога, но любой груз - это внешняя сила, стремящаяся растянуть мышцы. И здесь можно обнаружить, что ответное сокращение регулируется рефлекторно в зависимости от величины груза. В этом легко убедиться на практике: попробуйте перекреститься, а потом повторите эти же движения с пудовой гирей в руке, как это делали силачи в старом российском цирке.