

4.8. Механизм проведения потенциалов действия
Чем более сложные задачи решает мозг, тем большее количество нейронов ему необходимо. Однако вся масса нейронов должна поместиться в ограниченном черепом и позвоночным каналом пространстве, а потому нервные клетки должны быть небольшими, а их отростки - достаточно тонкими. Но, как известно, чем тоньше и длиннее проводник, тем большее сопротивление он окажет распространяющемуся по нему току. Действующее напряжение в нейроне (V) не может быть больше, чем амплитуда потенциала действия, т.е приблизительно 100-120 мВ, а ток (I), согласно закону Ома, прямо пропорционален напряжению и обратно пропорционален сопротивлению: I = V/R
Из этого следует, что потенциал действия обычным для проведения электричества способом не может распространяться далеко. Очень тонкая мембрана аксона, окруженная электропроводной средой, имеет очень высокую ёмкость, что тормозит распространение электрического сигнала. Если сказать проще: тонкий цитоплазматический отросток - это очень плохой проводник. Но, несмотря на это, потенциалы действия распространяются по аксону с высокой скоростью, достигающей 100 м/с. Как это происходит?
К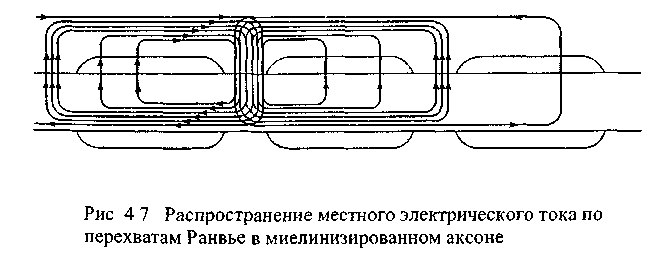 огда
в возбуждённом участке мембраны
повышается натриевая проницаемость и
возникает потенциал действия, начинается
электротоническое распространение
положительных зарядов к невозбуждённому
участку - этот процесс представляет
собой круговой ток (рис. 4.7). Такой ток
деполяризует ещё не возбуждавшийся
соседний участок, а когда эта
деполяризация достигнет порога -
возникнет потенциал действия. Теперь
уже этот участок становится источником
кругового тока, действующего на
следующую область мембраны, теперь уже
в этой области возникнет потенциал
действия, все параметры которого будут
стандартными для данного типа нейрона.
огда
в возбуждённом участке мембраны
повышается натриевая проницаемость и
возникает потенциал действия, начинается
электротоническое распространение
положительных зарядов к невозбуждённому
участку - этот процесс представляет
собой круговой ток (рис. 4.7). Такой ток
деполяризует ещё не возбуждавшийся
соседний участок, а когда эта
деполяризация достигнет порога -
возникнет потенциал действия. Теперь
уже этот участок становится источником
кругового тока, действующего на
следующую область мембраны, теперь уже
в этой области возникнет потенциал
действия, все параметры которого будут
стандартными для данного типа нейрона.
Вслед за повышением натриевой проницаемости в процессе формирования потенциала действия усиливается ток калия из клетки. Вместе с калием из клетки уходят положительные заряды и восстанавливается прежнее значение мембранного потенциала. При любой длине аксона амплитуда потенциалов действия повсюду оказывается неизменной, поскольку в каждом отдельном участке аксона они формируется фактически заново. В физиологическом смысле это важно потому, что постоянство сигнала означает передачу информации по аксону без искажений.
В миелинизированных аксонах круговой ток распространяется к соседнему перехвату, где и возникает потенциал действия. Плотность натриевых каналов в перехватах Ранвье гораздо выше, чем в обычной немиелинизированной мембране и пришедший сюда электротонически круговой ток легко деполяризует перехват до порогового значения. Появившийся вследствие этого потенциал действия служит источником кругового тока для следующего перехвата.
Проведение возбуждения в нерве или мышце можно зарегистрировать с помощью внеклеточных электродов, приложенных к двум различным точкам их поверхности и соединённым с регистрирующей аппаратурой. При распространении потенциала действия мембрана поочерёдно деполяризуется сначала под ближним к источнику возбуждения электродом, а затем - под дальним. И в том, и в другом случае между электродами регистрируется разность потенциалов, поскольку один из них будет находиться на деполяризованном, а значит электроотрицательном снаружи мембраны участке, а второй - в интактной электроположительной точке, где возбуждение ещё не началось, либо уже закончилось.
Регистрация проходящих по мембране потенциалов действия с помощью двух электродов называется биполярной. При таком способе регистрируются две фазы потенциала действия: положительная и отрицательная. Если расположенный под одним из электродов участок сделать невозбудимым (для этого можно подействовать на него каким-либо анестезирующим веществом, например, новокаином), то останется только одна фаза потенциала действия. Такое отведение называется униполярным (или монополярным).
При некоторых аутоиммунных и вирусных заболеваниях миелиновая оболочка разрушается, что приводит к многочисленным неврологическим нарушениям, вплоть до полной утраты некоторых функций; при этом может нарушаться и эмоциональная деятельность, и интеллект. Примером демиелинизирующих заболеваний может служить рассеянный склероз.
Резюме
Возникновение электрических сигналов связано со свойствами клеточной мембраны. Мембранные насосы создают концентрационные градиенты ионов. Открытые в покое ионные каналы для калия позволяют ему выходить из клетки и, тем самым, создавать мембранный потенциал покоя, близкий к равновесному потенциалу для калия. В случае его уменьшения до порогового значения открываются потенциалзависимые каналы для натрия и происходит саморегенерирующая деполяризация, значение мембранного потенциала становится положительным, Это вызывает закрытие натриевых каналов, которые на время инактивируются. Выходящий ток ионов калия восстанавливает прежнее значение мембранного потенциала. Возникновение потенциала действия вызывает появление кругового электрического тока, который деполяризует соседний участок мембраны до порогового значения. В связи с этим потенциал действия распространяется по аксону без уменьшения амплитуды.