

Вывод уравнения Нернста.
Нернст
изучал поведение электролитов при
пропускании электрического тока и
открыл закон. Закон устанавливает
зависимость между электродвижущей
силой (разностью потенциалов) и ионной
концентрацией. Уравнение Нернста
позволяет предсказать максимальный
рабочий потенциал, который может быть
получен в результате электрохимического
взаимодействия, когда известны давление
и температура. Таким образом, этот закон
связывает термодинамику с электрохимической
теорией в области решения проблем,
касающихся сильно разбавленных растворов.
![]() ,
,
где
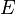 —
электродный
потенциал,
—
электродный
потенциал,
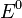 —
стандартный электродный потенциал,
измеряется в вольтах;
—
стандартный электродный потенциал,
измеряется в вольтах;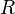 —
универсальная
газовая постоянная,
равная 8.31 Дж/(моль·K);
—
универсальная
газовая постоянная,
равная 8.31 Дж/(моль·K);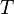 —
абсолютная
температура;
—
абсолютная
температура;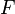 —
постоянная
Фарадея,
равная 96485,35 Кл·моль−1;
—
постоянная
Фарадея,
равная 96485,35 Кл·моль−1;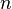 —
число
молей
электронов,
участвующих в процессе;
—
число
молей
электронов,
участвующих в процессе;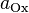 и
и
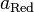 —
активности
соответственно окисленной
и восстановленной форм вещества,
участвующего в полуреакции.
—
активности
соответственно окисленной
и восстановленной форм вещества,
участвующего в полуреакции.
Если
в формулу Нернста подставить числовые
значения констант
и
и
перейти от натуральных
логарифмов к десятичным,
то при
![]() получим
получим
![]()
6. Потенциал действия. Последовательность изменения ионной проницаемости мембраны в процессе развития потенциала действия. Проведение по миелинизированным и демиелинизированным нервным волокнам.
Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляет электрический разряд — быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса.
Потенциалы действия могут различаться по своим параметрам в зависимости от типа клетки и даже на различных участках мембраны одной и той же клетки. Наиболее характерный пример различий: потенциал действия сердечной мышцы и потенциал действия большинства нейронов. Тем не менее, в основе любого потенциала действия лежат одни и те же явления.
Проведение по миелинизированным и демиелинизированным нервным волокнам.
Распространение потенциала действия По немиелинизированным волокнам.
По немиелинизированному волокну потенциал действия распространяется непрерывно. Проведение нервного импульса начинается с распространением электрического поля. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые потенциалы. Сам потенциал действия не перемещается, он исчезает там же, где возник. Главную роль в возникновении нового потенциал действия играет предыдущий.
Если внутриклеточным электродом раздражать аксон посередине, то потенциал действия будет распространяться в обоих направлениях. Обычно же потенциал действия распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник потенциал. Одностороннее проведение потенциала действия обеспечивается свойствами натриевых каналов — после открытия они на некоторое время инактивируются и не могут открыться ни при каких значениях мембранного потенциала (свойство рефрактерности). Поэтому на ближнем к телу клетки участке, где до этого уже «прошел» потенциал действия, он не возникает.
При прочих равных условиях распространение потенциала действия по аксону происходит тем быстрее, чем больше диаметр волокна. По гигантским аксонам кальмара потенциал действия может распространяться почти с такой же скоростью, как и по миелинизированным волокнам позвоночных (около 100 м/c).