

Гендерді тасымалдайтын векторлар
Курылымдык гендерде тек кана метаболизм өтудін нәтижесівде тузілетін заттардьщ (белоктьш, иРНК-нын) коды жазылған. Оларда ген активтілігін реттейтін бөлшек мулдем жок. Сондыктан, жада курылымдык гендерді иеленген клеткаларда ол гендер өз бетімен тиісті кызметін аткара алмайды. Гендердің клеткадағы әрекетін баскаратын репликация және транскриіщия сигнаддарын оларға вектор камтамасыз етеді.
Ііотен генді клетка ішіне тасымалдап алып баратын арнаулы ІІІК, молекуласын вектор дейді. Оған мынадай талаптар І< иіылады: 1) өз алдына репликациялану, яғни клетка ішіне бөтен '• ІІлІ алі.іп кірген сон клеткамен бірге немесе өз алдына көбейе ІІ.нмІІ болуы керек; немесе вектор клетка хромосомасынын І \рамына еніп, онымен бірге урпак клеткаларға беріліп отыруы І І'|ч'к; 2) Ірансформацияланған клеткаларды аныктау ушін онын І І>І'кІІІс геиетикалык белгілері (маркерлері) болуы керек (мысалы ІІІІІІбмотикке төзімділігі); 3) күрамында рестриктазалар үзе м.ІІшІ нуклеотидтер тізбегі болуы керек және репликацияға І ІИІ.ІКЧ ІІІ жогалтпауы керек; 4) векторға орналастырылған бөтен І н ІІІІын аткаратын кызметін бүзбауы керек, ал вектор болса, ол 11 сні і Ііліеп геннің ішінде дүрыс реттеліп жүмыс істеуін камтама-ІІІ стетін болуы керек; 5) вектордын көлемі кішігірім болуы керек.
'Чдс'Ітс, күрылымдык ген өте кыска болып келеді (бірнеше І \ І нуклеотид). Оны бірден көп мөлшерде бөліп алу киын. • чцдыктан онын көшірмелерін (молекулаларынын санын) ммсрліктей көбейту керек. Генді клондау үшін бактериялар міц.чаиылады. Мысалы, өте жаксы тексерілген, зияны жок, кең гІІ|КІлған бактерия - ішек таякшасы (Е.соіі). Керекті ген орна-ІІІІстырылған векторды бактерияға енгізеді. Бактериятез бөлінетін-/ІІкІсІІ, оның ішіндегі вектор да, ген де бактериямен бірге көбейеді. Лкырында өскеи бактерия биомассасынан вектор мен ген, яғни |Н'комбІІнанттык ДНК көп мөлшерде бөлініп алынады.
Бактерия плазмңцалары және рекомбинанпык ДНК күрастыру
ИскІорлар ретінде көбінесе ішектаякшасы Е.соіі және де баска ГІіІмсрмялардын плазмидалары колданылады. Бактерияларда бас-І һі хромосомадан баска көптеген (200-ге дейін) кішкентай сакина ІіІрІ Іді болып түйыкталған кос тізбекті ДНК, молекулалары кезде-ІІ-лІ, Олар бірнеше мын нуклеотидтерден түрады, плазмидалар ;К'ІІ агалады. Сакина сиякты ДНҚмолекулалары бір-біріне оралып нурпелі спираль күрайды. Плазмидалар өз бетіне репликациялана ІІлІІІыІІ ДНК, молекулалары (генетикалыкэлементтері). Оларбак-11'рияІІІ.Ің басты хромосомасымен тіркеспеген, клетка ішінде өз ІІ'І;ІІ,ІІІ;І еркін орналаскан. Плазмида күрамында тетрациклин 111'МІчт каііамишін сиякты антибиотиктерге төтеп беруді камтама-І'Ы 11'Іх'ІІІІ ферменттердін гендері бар. Плазмидаларды хромосома-иыі ,41 ІК-нап бөлекше таза түрінде алуға болады.
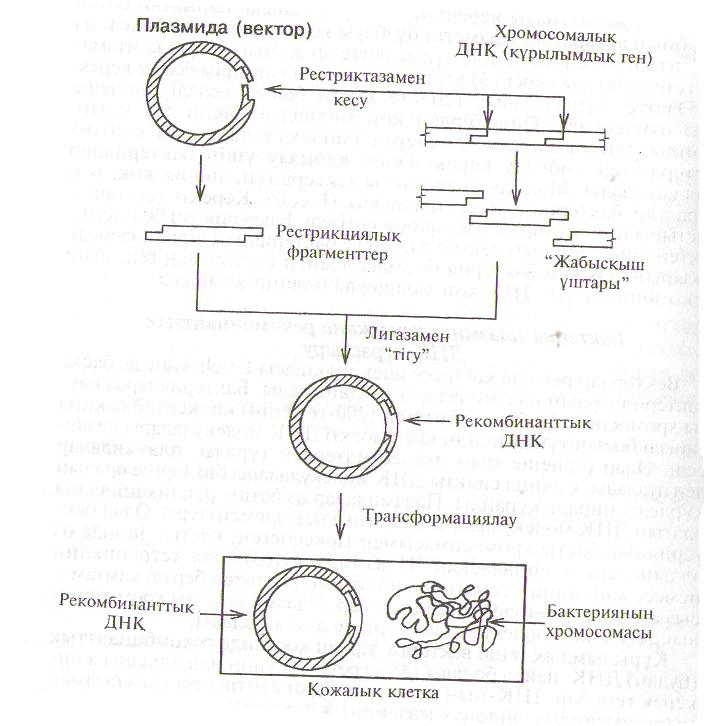
Күрылымдык генді векторға тіркеп коскавда рекомбинаштык (ПудаІІ) ДНК, пайда болады (48-сурет). Ол ушін плазмиданы және ксрск Іхчіі бар ДНҚ-нын бөлшегін (фрагментін) рестриктазамен ||нч Ірпкпиялык эндонуклеазамен) жіп сиякты кеседі. Олардын үіиІііры біртізбекті немесе мүкал басты болады. Мүкал және кыска піріІ (бі-кіі үштарды трансфераза ферментінін көмегімен бірнеше аденинді (А) немесе бірнеше тимидинді (Т) жалғап узартады. Сонда курылымдык геннін бір үшы А-А-А-А, ал екінші үшы Т-Т-Т-Т болады. Плазмиданьщ жіп сиякты болған молекуласы-ІІІ.мІ да екі ұшы сондай болады. Сөйтіп, А мен Т комплементарлык болғандыктаи, рестрикцияланған фрагменттерде жабыскак уштар паііда болады. Оларды кейін ДНК.лигазасымен бір-біріне жабыстырып косып, будан рекомбинанттыкДНКжасалады. Оны клондап көбейту үшін бактерия клеткасына енгізеді.
Плазмиданьш бір ерекшелігі, ол бір бактериянын клеткасынан кслесі клеткаға онай ене береді. Бактериянын ішінде оларрепли-
 48-сурет.
Рекомбинанттык ДНК молекулалар,
48-сурет.
Рекомбинанттык ДНК молекулалар,
Бактерияның хромосомасы
<ын жасау схемасы (41).
І аиияланып көбейе бастайды. Сонымен, бөтен ДНК-нын тран-< крипциясы да, трансляциясы да өтеді. Сөйтіп бөтен генде «жа-и.іліан» белок синтезделеді, яғни клетка трансформацияланады.
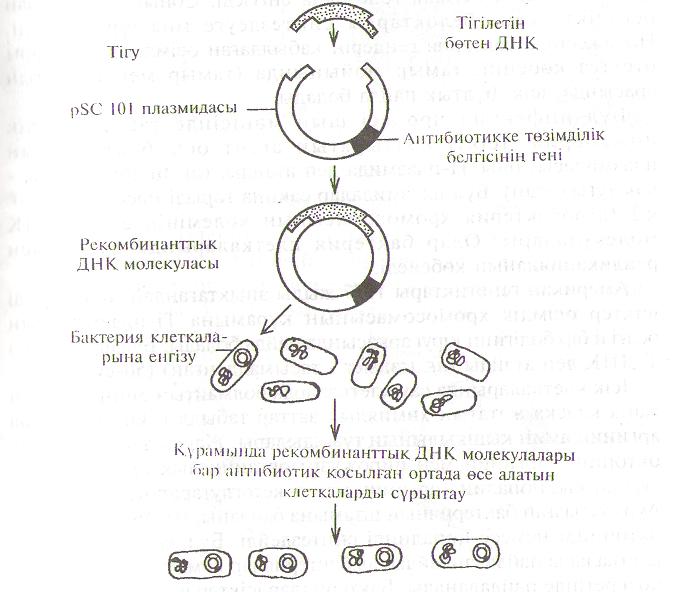
Г.соіі клеткасында эукариоттык геннін кайсысынын болса да Іксирессиясы өте алады. Кұрамына керек гені бар рекомбинант-ІІ,Ік ДНК молекуласын өзіне дарыткан бактерияларды сұрыптау \ІІІІІІ коректік ортаға антибиотикті (тетрациклинді немесе І лиамииинді) косады. Себебі, плазмидаларда антибиотикке ииімділік гені болады (49-сурет). Бүл жағдайда плазмидасы жок І,нггкалар кұриды. Сонымен, плазмидалар бөтен генді көбейту уиіііі нәтижелі колданылады.
Гожірибенін. максаты мен міндетіне байланысты әр түрлі ІІиачмидалар пайдаланылады. Өсімдіктер үшін ең лайықты вектор Д^ІоһасГепа деген топырак бактерияларынын плазмидалары.
 49
-сурег. Д//К фригментін
плазмидада клондау, яғни көбейту (23).
49
-сурег. Д//К фригментін
плазмидада клондау, яғни көбейту (23).
Лгробактврия шіазмидаларын вектор ретіиде колдану
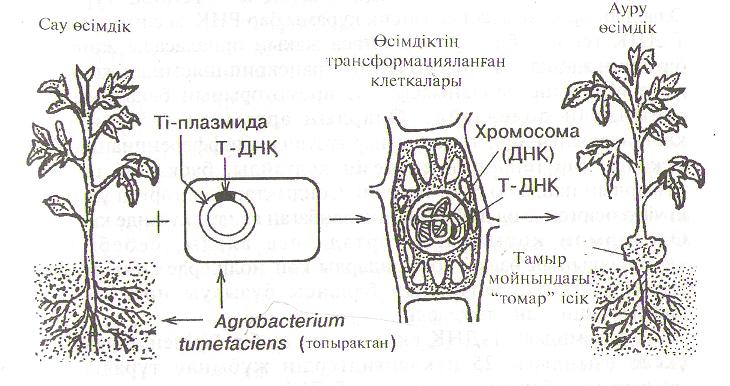
АцгоЬасгегіа деген топыракта мекендейтін бактериялардьщ тоОы. Олар өсімдіктерге жүғып тәж тәрізді өсіндіні, яғни ісікті, бултыкты пайда болғызады. Агробактерияньщ бірнеше турі бар. Бүл бактерияларға косжарнакты кен жапыракты өсімдіктердін барлығы дерлік сезімтал келеді, ал астык тукьшдастары мен баска да даражарнактыларға олар жукпайды. Тәж тәрізді ісік клеткалары іп УІІГО шексіз өсе береді, тіпті коректік ортаға гормондарды косуды да кажет етпейді, себебі оларды өз клеткаларында тузеді. Ісік ауруын ен онай коздыратын А§гоЬасІегіа Іипіеіасіеш. ЖалІІы айтканда, бул бактерия гендік инженерия көздеген максатты табиғатта іс жузінде аткарады. Бул бактерия өсімдіктін жаракаттанған улпасы аркылы клеткаға кіргенде өзінін гендерін біргс ала келіп, өсімдік геномына енгізеді. Сонын салдарынан осімдікті жана белоктарды синтездеуге мәжбур еткізеді. Нәтмжесінде бактерия гендерін кабылдаған өсімдік клеткалары оте тез көбейіп, тамыр мойынында (тамыр мен сабактьш арасында) ісік, бултык пайда болады.
Бул инфекция процесі шын мәнісінде табиғи гендік ипжеперия. Ісікті коздыратын агент осы бактерияньш плазмидасы, оны Ті-плазмида деп атайды. (ағ. штог іпс!исіп§ -ісік туғызатын). Бул плазмидалар сакина тәрізді массасы 1,2х1(Ғ кД (агробактерия .хромосомасынын көлемінін 3-5%) ДНК молекулалары. Олар бактерия клеткаларында өз бетімен реіІлнкацияланып көбейеді.
Американ генетиктары 1977жылы аныктағандай, тәж тәрізді ісіктср өсімдік хромосомасынын курамьша Ті-плазмиданьш белгілі бір бөлігінін кіруі аркасында пайда болады. Бул фрагментті Т-ДНҚдеп атайды (ағ. ІгапзҒег - тасымалданған) (50-сурет).
Ісік клеткаларында сау клеткаларда болмайтьш опиндер деген жапа класка жататын химиялык заттар табылды. ОІшндер - ол аргинин амин кьшікылыньщ туындьшары. Жаксы зерттелгендер: октолин - аргинин мен пирожузім органикалык кышкылъшьщ ІуІ.Іплысы, нопалин - аргинин мен а-кетоглутараттың туындысы. Ауру гуғызған бактериянын штамына байланысты ісік клеткалары октоминді немесе нопалинді синтездейді. Бул заттарды өсімдік кле Іжалары пайдаланбайды, ал бактериялар көміртегі мен азоттьщ кезі ретінде пайдаланады. Бактериялар ісіктерді туғызумен бірге, өсімдіктердін метаболизмін біраз өзгертеді. Өсімдік клеткалары гек бактерияларға кажет амин кышкылдарын синтездеуге кіріседі. Бул паразиттік өзара катынас генетикалык отарлау деп аталады. Сойпп бүл табиғатта нәтижелі өтетін генетикалык инженерияның үлгісі.
 50-сурет.
Тибиғн жағдайда агробактерия гендерінін
өсімдікке енуі (65).
50-сурет.
Тибиғн жағдайда агробактерия гендерінін
өсімдікке енуі (65).
Ті-ІІлазмидалар өздерінін күрамында жазылған опин турі боііынша жіктеледі. А.ІитеҒасіепз клеткасында плазмида-иың бір турі ғана, октопин немесе нопалин синтездейтін турі болады (51- сурет). Т-ДНК-ны өсімдік клеткасына тасымалдау
51-сурет. Октоплндік және нопалиндік Ті-плазмидаларынын гснетикялык карталары (65)
және хромосомалык ДНК күрамына кірістіру процесіне жауапты гендер плазмида бойында Т-ДНҚ-дан алшак орналаскан. Сол сиякты опиндерді ыдырататын гендер де Т-ДНК-дан алшашрак орналасады. Қазір Т-ДНК-нын толык картасы жасалып, онда кандай гендер калай орналасатыны белгілі болды.
Опинді ісік клеткаларьшда Т-ДНК-ы 7 геннен тұрады. Олардын әр кайсысында өзіндік күрамы бар РНҚ-ы синтезделеді. Т-ДНК гендері бір-біріне жалғаса жакын орналасады және де бір-бірін жаппайды. Бүл әр геннің транскрипциясынын жекеден жеке бір-біріне байланыссыз, өз промоторының бакылауымен өтетіндігін дәлелдейді, Олардын арасында 4-5 ген ісік клеткаларының өркен мен тамыр түзушілік дифференциациясын тежейді. Бір ген опин синтезін кодтайды, баска екі гендер тамырдын пайда болуьш тежейді. Сондыктан тәж тәрізді үлпаны іп үііго өсіргенде ол дифференцияланбаған каллус күйівде калады. Ол гормон косылмаған ортада өсе алады, себебі ісік клеткаларының өздері гормондарды көп мөлшерде синтездейді. Өсуді реттейтін заттардьш балансы бүзылуы нәтижесінде дифференциялану тежеледі.
Ті-плазмидада Т-ДНК, екі үшынан да бір-бірінен аумайтын, үксас үзындығы 25 нуклеотидтердін жүбынан түрады. Ол нуклеотид тізбектерінің, мүмкін, Т-ДНҚ-ны өсімдік клеткасына тасымалдауына катысы бар шығар. Ісіктін пайда болуына Ті-плазмида дәл өзі себепші екені (хромосомалык гендер емес), плазмидалары жок немесе мутанттык Ті-плазмидалары бар агробактерия штамдарын колдану аркылы дәлелденді. Ті-плазмидалары жок агробактериялар жүккан өсімдіктерде ісік пайда болмайды және де опиндер синтезі өтпейді. Мутанттармен өткізілген генетикалык зерттеулер көрсеткендей, Ті-плазмиданын Т-ДНК фрагменті ғана ісіктін пайда болуына, опиңцер синтезіне және дифференцияланудын тежелуіне жауапты.
Ісік тудыратын кабілеті төмен мутант - октопин түзгіш плазмида алынған. Онын бағалылығы, ол ісік үлпасына жакын орналаскан нормалы клеткалардын аномальдык дифференциров-касын арттырады. Мысалы, тамыр мен өркеннің күшті өсуіне себепкер болады. Осы плазмидамен трансформацияланған темекі клеткалар популяциясында кейбір жеке клеткалар (өте сирек болса да) регенерант өсімдігін берген. Бүл өсімдіктердің барлык ұлпалары октопинді синтездеу кабілетін Т-ДНК-нын болуы нәтижесінде сактаған. Осы трансформацияланған өсімдіктерді нормалы темекі өсімдігімен будандастырып, кейіннен сол буданнан алынған үрыктарды өсірген. Сонда көрсетілгендей, октопин синтездеуге кабілеттілік (яғни Т-ДНК) Мендель зандылығына сәйкес аналык және аталык жағынан да түкым қуалаған. Демек, Т-ДНК хромосомаға тіркескеннен кейін өсімдіктін әдеттегі гені болып сініп кетеді.
Өсімдікке енгізілген Т-ДНҚ Мендель зандары бойынша түкым куалайтын болғандыктан және онын гендерінін өздерінін промоторлары (сол промоторлардын бакылауында бөтен гендердін экспрессиясы өте алады) болу арқасында Ті-плазмиданың ДНК,-сын гендік инженерия жүмыстарында вектор ретінде пайдалануға
болады.
Агробактерияларды кабылдап, оларға пана бола алатын өсімдіктер өте көп, олар косжарнакты өсімдіктердін барлығы дерлік. Өкінішке орай, көптеген ауыл шаруашылығында маңызы зор дакылдар, атап айтканда: бидай, кара бидай, сүлы, күріш, жүгері, даражарнакты өсімдіктерге жатады. Сондыктан олар агробактерияларға төзімді келеді. Даражарнакты өсімдіктердін тәж тәріздІ ісік ауруына табиғи төзімділігі жөнінде түрлі пікірлер бар. Мүмкін, бактериялар олардын клеткаларына мүддем кіре алмайды немесе кіргеннін өзінде, Т-ДНК-ны олардын хромосомаларына тіркестіре алмайды.
Бірак әдебиеттегі бірен-саран деректер бойынша, кейбір даражарнакты өсімдіктер, мысалы каскыржем (спаржа), лалагүл, нәркес, жүгері, өздеріне агробактерияларды жүктырады және ісік ауруына шалдығады. Ісік ұлпалары гормондар косылмаған ортада іп ҮІІго өсе алады, олардын күрамында Т-ДНК, кодтайтын опиндер түзілетіндігі аныкталған. Сөйтіп, бүл деректер бойынша, тым болмаса кейбір даражарнакты өсімдіктердін де агробактериянын Т-ДНҚ-ны кабылдауы және оларда сол Т-ДНҚ гендерінін экспрессиясы өтуі мүмкін.
Плазмиданы вектор ретінде пайдаланудын бір киыншылығы, онын көлемінін үлкендігі (18 кД). Т-ДНК-ны өсімдік клеткасына тасымалдау және хромосомаға тіркестіру үшін онымен шекаралас жаткан нуклеотидтер тізбегінін және де оған коса ісік тудыратын вируленттік (УІГ) бөлігінін кажет екендігі аныкталды. Сондыктан, казір бүтін Ті-плазмиданы толышмен емес, тек онын жоғарыда аталған УІГ бөлшегін ғана пайдаланады. Ті-плазмиданын Т-ДНК фрагментін вектор ретінде колданудағы тағы бір кедергі, ол Т-ДНК аркылы трансформацияланған клеткалардан бүтін регенерант өсімдіктердін шыкпауы.
Егер де Т-ДНҚ-нын орта шенін киып, бөліп алып тастаса, Т-ДНК, өзінін ісік туғызатын онкогендік кабілетінен айырылады, бірак өсімдік клеткасы хромосомасымен тіркесе алады. Соның нәтижесінде, осылай трансформацияланған клеткалар регенерант өсімдіктерді жаксы түзе алады. Табиғи жағдайда Т-ДНК, -нын «тамыр бергіш» деп аталатын мутанттары болады екен, Олар өсімдік клеткаларьш трансформациялай алады және регенерацияны тежемейді. Сондай, «тамыр бергіш» мутацияға үшыраған Т-ДНК,, транспозондар көмегімен де алынған. Бүл өзгеріске үшыраған Т-ДНК, өсімдік клеткасынын хромосомасына оп-онай орналасып, оны трансформациялап, регенерация процесінщ өтуіне кедергі жасамайды. Ол мутация Т.-ДНҚ картасынын белгілі бір бөлімінде орналаскан.
Сонымен, вектор ретінде пайдалануға жарамды Т-ДНК мына талаптарға сай болуы керек:
1) ДНК-ны өсімдік клеткасыньщ ядросына ендіру үшін және оны хромосомаға түракты орныктыру үшін барлык сигналдары бар;
2) осы процестерді камтамасыз етуге кажет метаболиттік заттарды түзу үшін генетикалык информация «жазылған»;
3) клеткадан регенерант өсімдік шығаруға кедергі жасамау;
4) өсімдік геномына енгізілген бөтен гендердің экспрессиясын жүзеге асыру үшін арнайы жүйесі бар;
5) трансформацияланған клеткаларды сүрыптау үшін маркеры бар;
6) бөтен ДНК, бөлшегін осы вектор ретінде колданылатын Т-ДНҚ-ға енгізу жүмысы онай әдіспен орындалуы керек.
Күрамына кажетті ген тіркестірілген Т-ДНҚ-ны (рекомбинант-тык ДНК-ны) өсімдік клеткасьша енгізудін екі әдісі жете зерггеліп дайындалған.
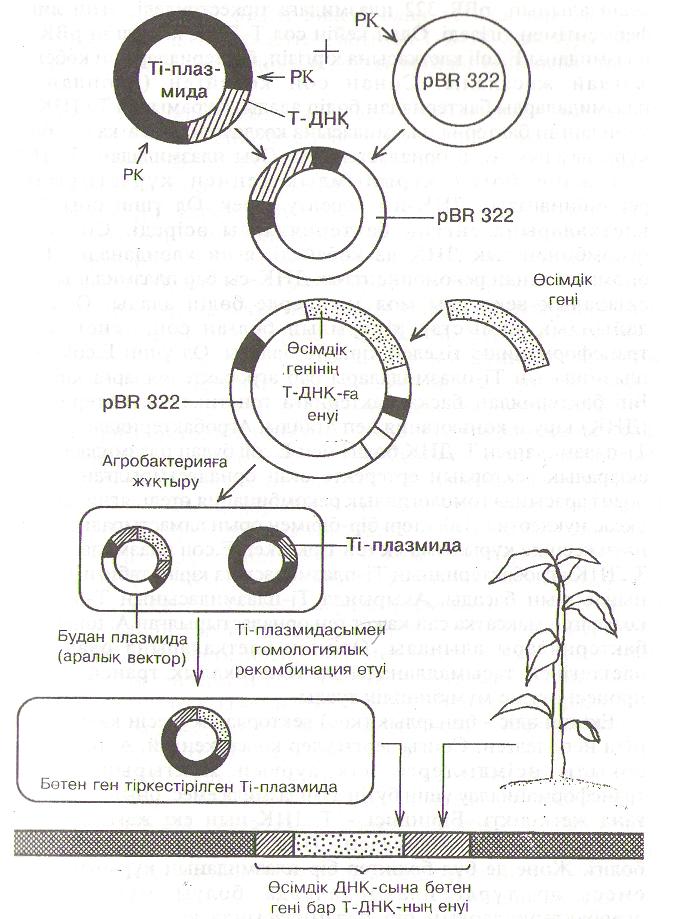
Бірінші әдіс - «екіаралык векторлар» әдісі - ішек таякшасы бактериясынын (Е.соіі) рВК 322 плазмидасын колдануға негізделген (52-сурет). Рестриктазамен Т-ДНК Ті-плазмвдадан
 52-сурет.
Ті-шизшдааы вектор ретіндс коддану
әдісі (23), РК - рестриктазамен кесу.
52-сурет.
Ті-шизшдааы вектор ретіндс коддану
әдісі (23), РК - рестриктазамен кесу.
кесіп алынып, рВК. 322 плазмидаға тіркестіріледі, яғни лигаза ферментімен тігіледі. Одан кейін сол Т-ДНК косылған рВК 322 Ішазмиданы Е.соіі клеткасына кірғізіп, бактериялардын көбеюіне жағдай жасалады. Сонан сон көбейген (клонданған) плазмвдаларды бактериядан бөліп алады. Күрамывда Т-ДНК бар клонданған бактерия плазмвдасына көзделген максатка сай бөтен күрыльшдык генді орналастырады. Осы плазмидадан, Т-ДНК,-дан және бөтен күрылымдык геннен күрастырылған рекомбинанттык ДНК-ны көбейту керек. Ол үшін оны Е.соіі клеткаларына енгізіп бактерияларды өсіреді. Сол кезде рекомбинанттык ДНҚ да көбейеді, яғни клонданады. Е.соіі биомассасынан рекомбинаштык ДНК-сы бар плазмиданы, яғни екіаралык векторды мол мөлшерде бөліп алады. Осындай дайындык жүмыстар аткарылып болған сон, генетикалык трансформацияға тікелей кірісуге болады. Ол үшін Е.соіі будан плазмидасын Ті-плазмидалары бар агробактерияларға кіргізеді. Бір бактериядан баска бактерияға генетикалык материалдын (ДНК) кіруін конъюгация деп атайды. Агробактериядағы табиғи Ті-плазмиданың Т-ДНК бөлігі мен Е.соіі будан шіазмидасындағы екіаралык вектордың ертеректе оған орналастырылған Т-ДНҚ бөлігі арасында гомологиялык рекомбинация өтеді, яғни олардын үксас нуклеотид тізбектері бір-бірімен орын алмастырады. Соньщ нәтижесінде күрылымдык ген тіркескен Е.соіі плазмидасындағы Т-ДНК агробактериянын Ті-плазмидасына кіріп, табиғи Т-ДНК-нын орнын басады. Акырында Ті-плазмидасынын Т-бөлігінде көзделген максатка сай кажет ген орналастырылған А. ІитеҒасіепз бактериялары алынады. Өсімдік клеткаларына олар кейін әдеттегідей тасымалданады да генетикалык трансформация проиесі өтуіне мүмкіншілік туады.
Екінші әдіс - бинарлык (кос) векторлар жүйесін кальштасты-руға негізделген. Сонғы зерттеулер көрсеткендей, А. ШтеҒасіеш аркылы өсімдіктерге ісік ауруын жүктырып, оларды трансформациялау үшін бүтін Ті-плазмида емес, онын екі бөлшегі ғана жеткілікті. Біріншісі - Т-ДНҚ-нын екі жағындағы тек шекаралык бөліктері, баскасы - Ті-плазмиданын вируленттік (УІГ) бөлігі. Және де бүл бөліктер бір плазмиданын күрамында ғана емес, әр түрлі плазмидаларда болуы мүмкін. Егер агробактериялардың бір плазмидасында үіг бөлігі болса, ал екіншісінде Т-ДНК бөлігі болса, олар өсімдік клеткаларын трансформациялауға кабілетті. Сонымен Т-ДНҚ-ның күрамында кандай да бөтен ген ендірілсе де, ол өсімдік геномына тіркесе алады.
Бірак солай болғанньщ өзінде де, Ті-плазмидалык векторлар арқылы өсімдікке енгізілген бөтен гендердің экспрессиясы өтпейді. Мысалы, темекі геномына Ті-плазмидалык векторлар көмегімен өз промоторларымен бірге енгізілген бактерия (антбиотиктерге карсы ферменттер гендері), жануар және өсімдік (леггемоглобин гені) гендерінін экспрессиясы жүрмеген. Әр геннін алдында 40-50 нуклеотидтен түратын тізбекті промотор деп атайды. Промотор геннін транскрипциясы (иРНК-нын синтезі) басталуына жауапты. Бөтен гендердін экспрессиясын кадағалау үшін міндетгі түрде Т-ДНК-нын күрамында ерекше промотор болады. Ті-плазмиданын нопалинсинтетазаны кодтайтын гені бөлініп алынып, онын күрамы зерттелді. Онын промотор бөлшегі де аныкталды және жеке бөлініп алынды. Осы промоторды октопинсинтетаза және хлорамфениколацетил-•грансфераза күрылымдык гендерінің алдына кіргізіп койып будан ДНК жасалған. Сондай будан ДНҚ-ны клондап көбейткеннен кейін өсімдік клеткаларына енгізген. Тек сонда ғана бөтен гендердін экспрессиясы жүре бастаған, яғни бүл процесті нопалинсинтетаза генінін промоторы реттеп отырған.
Сөйтіп, Т-ДНК-нын промоторы күшті болғаны, яғни онын нуклеотид тізбегі РНК-полимераза ферментін тез танитындай немесе онымен онай байланыска түсетіндей болып күралған. Тәжірибелер көрсеткендей, нопалинсинтетазанын промоторы қүрылымдык гендермен Ті-плазмида онай тіркеседі және өсімдік клеткаларыида өз кызметін аткара алады. Бүл промотордын тағы бір артыкшылығы, ол каллустарда да, өсімдіктердің көптеген мүшелерінде де өз кызметін аткара алады.
Т-ДНҚ-ға тігілген геннін экспрессиясын байкау ушін трансформацияланған және өзгермеген клеткаларды ажырата білу керек. Бүл үшін селективтік маркер (белгісі) колданылады, мысалы антибиотиктерге төзімділік белгісі. Химералык гендер, яғни колдан күрастырылған нопалинсинтетазаның промоторы, бактериянын канамицинге төзімділік гені және нопалинсинтета-иіиың терминальдык сигнал гендері темекі клеткаларына енгізілгенде жаксы нәтиже алынған. Трансформацияланған өсімдік клеткалары канамицинге төзімділік көрсеткен. Осы химералык гендер кейін трансформацияланған клеткаларды сурыптау үшін селективтік маркер ретінде колданылып, фенотип жағынан нормалы және үрыктанушы регенерант өсімдіктерін алуға мүмкіндік берді. Бүл өсімдіктерде антибиотикке төзімділік генінің экспрессиясы барлык үлпаларында өткен, кейін ол касиет Мендель заны бойынша түкым куалаған. Сонымен, агробактерия-лардың модификацияланған Т-ДНК-сы көмегімен өсімдіктер клеткаларын трансформациялау әдісі казіргі уакытта ен тиімді болып табылады.
А§гоЬас1егіит гһігоёепез «сакалды тамыр» аталатын ісік ауруын тудыратын топырак бактерияларынын Кі-плазмидаларын (ағ. гооі іпсһісіпё - тамыр шығарғыш) да өсімдіктер гендік инженериясыңда вектор ретінде колдануға болады. Өсімдіктердін жараланған жерлерінен сол бактериялар жүкса, онда сакал кылшыктары сиякты жінішке тамырлар өсе бастайды. Бактериялар болмағаннын өзінде, ауру жүккан тамыршалар іп УІІГО тез өседі, себебі олардьщ клеткаларында бактерия Кі-плазмидасынын Т-ДНК-нын бірнеше көшірмелері және опиндер бар. «Сакалды тамыр» ауруының салдарынан пайда болған тамыршалардың тәж тәрізді ісіктерден айырмашылығы, олардан іп УІІГО жағдайында регенерант өсімдіктер онай шығады. Ол регенерант өсімдіктер аурудан сау және үрык бере алады, жапырактары мен тамырларында опиндер болады. Олардын геномдарында Кі-плазмиданың Т-ДНК, бөлігі жыныстык жолмен түкым куалайды.
Француз ғалымы ЖТампе сәбізбен мынадай тәжірибе өткізген. А.гһІ2О§епез бактериясын жүктыру нәтижесінде сәбіз кесіндісінде көптеген тамыр кылшыктары өсіп шыккан. Жеке тамыршаны кесіп алып агарлы ортаға көшіріп, одан каллус алған. Каллусты сүйык ортада өсіргенде эмбриоидтар түзілген. Кейін олардан регенерант өсімдіктер шыккан. Демек, Кі-плазмидалар табиғи, зиянсыз, гендер тасымалдаушылары бола алады. Кі-плазмида жүйесін колданудағы басты артыкшылык, ол үлпа емес, мүше өсіруге негізделген. Сонын нәтижесінде әр өсімдік түріне тән хромосомалар саны (плоидтылығы) сакталады.
Сонымен, Ті-плазмидалар мен Кі-плазмидалар жоғары сатылы өсімдіктерге гендерді тасымалдайтын өте колайлы векторлар.
Бірак, өкінішке орай, агробактериялардын Т-ДНК-ны өсімдік клеткасынын ядросына тасымалдау механизмі жөнінде мәлімет өте аз. Октопин және нопалин плазмидаларылын Т-ДНК сегменттері кожалык клетканын хромосомаларының кездейсок жерлеріне барып тіркеседі. Сонымен катар, олар ешкашанда митохондриялар мен хлоропластар ДНК-мен әрекеттеспейді. Бүл жағдай агробактериялар аркылы жүргізілетін генетикалык трансформация цитоплазмалык геном емес, тек кана ядролык геном аркылы өтетінін дәлелдейді.
Өсімдік клеткаларына Т-ДНК калай енгізілуі жөнінде әр түрлі көзкарастар бар. Мүмкін, агробактериянын өзінде Т-ДНК, плазмидадан кесіп алатын ерекше фермент бар шығар. Одан кейін бос Т-ДНК өз бетімен өсімдік клеткасына еніп кетуі мүмкін. Т-ДНҚ плазмиданъщ ісіктерде түракты сакталатын жалғыз бөлігі болғанымен, онын нуклеотидтік күрамында баска маңызды гендер (өсімдік клеткасын тану, онымен әрекеттесіп ішіне кіру, кейін хромосомаға Т-ДНК-ньщ тіркесуіне жауапты гендер) табылмаған. Шамасы, Т-ДНК-нын өсімдік геномьша тұракты тіркесуі үшін осімдік гені активтелінеді, болмаса плазмиданын үіг бөлігінде орналаскан гендер кодтайтын бірнеше белоктар Т-ДНК-мен коса тасымалдануы керек шығар. Бәрінен де дүрысы, тасымалдау ІІроцесі Ті-плазмидада және бактериянын хромосомасында орналаскан УІГ гендерінің бакылауында өтетін болуы ыктимал. Ті-плазмиданьщ вируленттілік гендері өсімдіктің закымданған клеткаларындағы фенол косындысынын әсерімен индукцияланады.
Баска гипотеза бойынша, Ті-плазмида түгелімен, тіпті бактерия геномы толығымен, өсімдік клеткасына тасымалданады, бірак осімдік ДНК-мен тек кана бактерия плазмидасынын Т-ДНК-сы тіркеседі. Онын себебі, өсімдік хромосомаларына тек кана Т-ДНКорналасады. Т-ДНКядроға енген кезде, оньщ молекуласы әр күйде болуы мумкін: линиялык немесе сакиналык, біртізбелік немесе костізбелік, бос, жеке өзі ғана немесе бактериялык белоктармен байланыскан түрде. Т-ДНК өсімдік клеткасында өз бетімен жеке репликациялана алады немесе лезде өсімдік геномына еніп кетеді де сонымен бірге көбейеді.
Т-ДНК өсімдік клеткасына калай енеді және хромосоманың күрамына калай кіреді? Мүмкін, бактериялар өсімдік клеткасына белгісіз бір жолмен әсер етіп, оны өзінін ДНК-сы оңай енетін ерекше күйге айналдырады немесе бактерияларды өздерін кабылдауға бейімділігі бар өсімдік клеткаларын тану үшін ерекше туыстығы болады. Сірә, Т-ДНК плазмидадан кесілуі және оның кожа клеткасынын хромосомасына енуі кезінде, Т-ДНК, үштарындағы сигналдык тізбектері манызды ролъ аткарады. Бірак осы сигналдарды танитын күрылым кандай екені және де кай организмге (бактерия немесе өсімдік) жататындығы әлі белгісіз. Колдан күрастырылған Ті-плазмидаларды өсімдік клеткасына кіргізу үшін бірнеше әдістер колданылады. Ен карапайымы, ол табиғи әдіс - жасалған бактериялар штамдарын өсімдікгерге жараланған жерлері аркылы жүкгыру. Екінші әдісі, ол прото-пластарды бактериялармен бірге өсіру аркьшы трансформациялау. Осылай бірге өсіру әдісі - бүл жасанды жағдайда ісікті туғызу әдісі. Егер агробактериялар жанадан алынған немесе біркүндік протопластармен бірге өсірілсе, бактериялардын оларға косылуы және трансформациялану процесі жүрмейді. Трансформация өтуі үшін кабықтары түзілген үш күндік протопластар кажет. Клетка кабығыньщ түзілуін тежейтін заттар бактериялардын прото-пластарға косылуын да тежейді. Бір тәулік бойынша бірге өсіргенде протопластар бактериялармен косылады, одан кейін косылмаған бактерияларды шайъш алып тастайды. Сонан сон өсімдік клеткаларын гормондары жок ортаға көшіреді. Бүндай ортада тек трансформацияланған клеткалар тірі калады. Осы әдіспен темекі мен шырайгүлдін трансформацияланған регенерант өсімдіктері алынған. Бүл әдіс агробактерияларға кожа бола алатын өсімдіктердін санын көбейтуге мүмкіндік береді, солардың ішінде астык түкымдастар түрлері де болады. Бірге өсірудін тиімділігін арттыру үшін клеткалардын косылуын коздыратын заттарды колдануға болады (ГТЭГ, кальций т.б.)- Протопластардын трансформациясын оларды Ті-плазмидалармен бірге өсіру аркылы да жүргізуге болады. Бүндай тәжірибелер шырайгүл, темекі протопластарымен жасалған. Бүл жүмыстарда химиялык затгарды (ПЭГ) колдану Т-ДНК, протопластарға енуін едәуір арттырған. Осы әдісті колдану негізінде трансгендік өсімдіктер алынды. Бүл әдістін артыкшылығы, ол аралык векторлардын кажет болмауы.
Хлоропластык ДНК және митохондриялык ДНК. вектор ретінде колдану
Хлоропластар мен митохондрияларда толык және дербес генетикалык жүйе бар (ДНК, ДНҚ-полимеразалар, Ғ'НК-полимеразалар және белок синтездейтін аппараттын рибосома-лары, тРНК, аминоацил-тРНК-синтетаза). Хлоропластык ДНҚ және митохондриялык ДНК вектор ретінде колдану мүмкіндігі ғалымдардьш назарын аударады. Бүл органоидтардың генетикалык жүйелерінде өзара айтарлыктай айырмашылыктары бар.
Хлоропластар мен барлык баска пластидалардың (лейкопласт, хромопласт) ДНК-ларындағы генетикалык информация бірдей, оны пластом деп атайды. Жоғары сатыдағы өсімдіктерде хлоропластык ДНК молекуласы сакина тәрізді, үзындығы 150 мын жүп нуклеотидтерден (мжн) түрады. Онда орналаскан гендер жүздей белоктарды кодтауға жарайды. Бірак пластидаларды толығымен күрастыру үшін одан едәуір көп белоктар кажет. Сондыктан, баска бірталай белоктар ядродағы ДНК гендерінде жазылып, цитоплазмада синтезделіп, кейін хлоропластарға тасымалданады. Хлоропластардын кейбір манызды белоктары бірнеше бөлшектен түрады. Ол бөлшектердін кейбірі хлоро-пластык ДНК-да кодталып, өзінің ішінде стромада синтезделеді, ал баскалары ядролык ДНК-да кодталып, цитоплазмада синтезделіп, кейін хлоропластка өтіп, бөлшектер бір-бірімен косылып, күрделі белок молекуласын күрайды. Сөйтіп, активті хлоропласт пайда болу үшін ядролык геном мен шіастомдык геном экспрессиясы үйлесімді жүруі керек.
Жоғары сатыдағы өсімдіктердін митохондриялык ДНК көлемі 200 мжн 2400 мжн дейін жетеді. Бірак ДНК көлемімен онын синтездейтін полипептидтер саны арасында ешкандай байланыс жок. Бүл күбылысты митохондриялык ДНК күрамында пайдасы жок ДНҚ («эгоистык», «надан» ДНК) мол болуы ыктимал деп түсінуге болар.
Митохондриялык ДНК, 20-дан астам күрылымдык гендер (митохондрияда синтезделетін полипептидтерді кодтайды) және де рРНК гендерін камтиды. Бірақ митохондриядағы белоктардын көбі хлоропластардағыдай ядролык гендермен кодталады. Хлоропластар геномы ірі сакина тәрізді ДНК молекулаларынан тұрса, митохондриялық ДНҚ молекулалары кіші де, ірі де сақина тәрізді келеді. Аталық цитоплазмалық ұрықсыздық (АЦҰ) белгісі кішкентай сақина тәрізді ДНҚ молекулаларының қызметімен байланысты болуы мүмкін. АЦҰ белгісі деген ол пісіп жетілген өсімдіктің кәдімгі сапалы тозаңды түзуге қабілетсіздігі. Бұл белгі Мендель заңы бойынша емес, цитоплазмалық жолмен, аналық жағынан тұқым қуалайды. АЦҰ белгісі көбінесе митохондриялық геномдағы өзгерістермен байланысты. Жүгері митохондрияларындағы әр түрлі ДНҚ, арасында осы ұрықсыздық белгісі шамасы S1, және S2 нуклеотид тізбектерімен байланысты. Бұл плазмидаға ұқсас ДНҚ молекулалары, ұзындығы 6,4 және 5,4 мжн линиялық молекулалар болып бөлініп алынады. Олар тек қана АЦҰ белгілері бар линияларында (АЦҰ-S) кездеседі. Осы жүгері өсімдіктерінің ядросында S1 , гомологиялық нуклеотид тізбектері байқалған. Мүмкін, S1 және S2 нуклеотид тізбектері АЦҰ белгісі бар N линияның митохондриялық хромосомасынан ажырап бөлініп шыққан. Ол былай өтуі мүмкін: 1) S1 мен S2 тізбектері митохондриялық хромосомадан ажыраған; 2) вирустың ДНҚ-сы сияқты олардың амплификациясы жүрген; 3) S1 тізбегі кейін ядроға тасымалданған; 4) геномда S1 тізбегі өз орнын өзгерткен.
Сонымен, S1 және S2 тізбектері, жүгері клеткасының табиғи компоненттері болғандықтан, өсімдіктерді трансформациялау үшін вектор ретінде қолдануға олардың бірегей мүмкіндіктері бар. S1 тізбектің негізінде жасанды вектор құрастыруға әбден болады. Сол векторға бөтен генді тіркестіріп, оны хромосомалық ДНҚ-на S1 тізбек арқылы транспозиция немесе рекомбинация жолымен орналастыруға мүмкіншілік туады.
Хлоропластык ДНҚ да вектор ретінде гендік инженерияның айлалы әрекетіне жарайды. Себебі, оның құрамында жеткілікті генетикалық информация бар, ал амплификация арқылы бөтен ДНҚ, экспрессиясын қамтамасыз етуге болады. Пластидалардың рибосомалық РНҚ гені мен рибулозобисфосфаткарбоксилаза ферментінің (РуБФК) үлкен бөлшегінің гені хлоропластық геномда орналасқан. Мүмкін, осы гендерге мықты промотор қойып және олардың қосымша модификациясын өткізу арқылы өсімдіктің фотосинтездік активтілігін арттыруға болар. Хлоропластқа бөтен генді енгізіп, өсімдікке гербицидке төзімділік қабілетін қамтамасыз етуге болады. Бұл мағынада хлоропластарды Ті-плазмиданың көмегімен трансформациялау қызықтырады, себебі Ті-плазмидаларды пайдаланып хлоропластың геномына генетикалық информацияны енгізуге болады.
Өсімдік гендерінің Е.соіі клеткаларында экспрессиясы өтуі жөнінде деректер бар. Бұл РуБФК үлкен бөлшегінің гені. Хлоропласт гендерінің артықшылығы, олар бактериялық ДНҚ-мен оңай қосылады және де жеңіл экспрессияланады. Себебі олардың екеуінің де ДНҚ-да трансляция процесінің басталуына қажетті инициация кодонының алдында бірдей нуклеотид тізбектері орналасқан.
Эукариот гендерін бактерияларға тасымалдап олардың экспрессиясын қамтамасыз етудің мақсаты неде? Ол эукариот полипептидтерінің (белоктарының) синтезін арттыру үшін прокариот гендері экспрессиясының қуатты механизмдерін пайдалану. Сонда, мысалы, өсімдік полипептидтерін бактерияларда синтездеуге болады. Бұл экономика жағынан құнды полипептидтерді мол алуға мүмкіндік береді, және де белокгардың жеке бөлшектерінің құрылымын зерттеу үшін маңызы зор.
Жылжымалы генетикалық элеметтерді вектор
ретінде қолдану
Жылжымалы генетикалық элементтер (транспозондар, «секіргіш», «көшпелі» гендер) - ДНҚ құрамында болатын және өзіндік құрылымы мен генетикалық қасиеттер жағынан ерекшеліктері бар нуклеотид тізбекгерінде орналасқан гендер тобы, олардың клетка геномында орын ауыстыруға қабілеті өте мол. Жүгері дәндерінің түрлі түсті болуының генетикалық негіздерін зерттеп тәжірибелер жүргізу нәтижесінде транспозондарды алғашкы ашқан Нобелъ сыйлығының жүлдегері Барбара Мак-Клинток (1950 ж.) Ол бұларды реттеуші элементтер деп атады. Бұл жылжымалы генетикалық элементтер қасындағы көршілес гендердің экспрессиясын реттейді. Олардың хромосомада белгілі тиянақты орны болмайды. Геномның әр жерінен ажырап шығып, басқа қандай болса сондай, кездейсоқ жеріне тіркесе береді. Жылжымалы элементтер өз орындарын бір хромосома бойында ғана емес, кейде басқа хромосомаға да көшіп ауыстырады. Олар ажыраған соң жанындағы бұрын жұмыс істемей тұрған гендер активтенеді. Ал реттеуші элементтермен қосылған гендер тұрақсыз, мутацияға жиі ұшырайтын болады. Қазіргі уақытта жылжымалы генетикалық элементтердің көптеген прокариоттық және эукариоттық организмдерде болатындығы анықталған. Шамасы, олар барлық тіршілік иелеріне тән шығар.
Барлық организмдерде байқалатын өзгергіштік пен тұрақсыздықтың басты себебі, осы жылжымалы генетикалық элементтер болуы мүмкін. Өйткені олар қасындағы көршілес басқа гендердің экспрессиясын тоқтатады немесе басқа жолдармен де ықпал етеді. Олар клеткада көптеген тұқым қуалайтын өзгерістер туғызады, себебі инсерциялық мутагенезді қоздырады. Инсерция деген ДНҚ молекулалардың генге тіркесуі, соның нәтижесінде ген инактивтеледі немесе оперонда күшті полярлық эффект байқалады. Хромосомалардың әр түрлі жерлеріне келіп тіркесіп, олар клеткалық гендердің активтігін тежейді немесе арттырады, яғни хромосоманың құрылымы өзгереді. Сонымен, жылжымалы гендер өздерінің орнын өзгертуі (транспозиция) арқасында клетканың генетикалық тұрақсыздығына, мутагенезге және хромосомалардың аберрацияларына түрткі болады.
Осындай ДНҚ бөліктерінің геном бойы жылжып орын ауыстыруының механизмі белгісіз. Мүмкін, жылжымалы гендер өздерінің аумалығын және репликациясын қамтамасыз ететін арнаулы белоктарды кодтайды. Әдетте, жылжымалы генетикалық элементтер геномда көп рет қайталанады, хромосомаларда кездейсоқ орналасады, транспозаза арқылы олар геномда орнын ауыстыра алады. Транспозаза деп жылжымалы элементтермен ерекше әрекеттесіп, оларды хромосомадан кесіп бөліп алатын ферментті атайды. Егер жылжымалы элемент дәл кесіп алынса, онда ДНҚ-ның бұрынғы құрылымы өзгермейді. Бірақ-та кейде бұл процесс нақты өтпейді, сондықтан кейбір нуклеотидтердің делециясы (жойылуы) немесе инсерциясы (қосымша пайда болуы) орын алады. Соның нәтижесінде тұрақты мутациялар пайда болады. Демек, бұл құбылысты жаңа ДНҚ тізбектерінің жасалу механизмдерінің бірі деп есептеуге болады.
Жылжымалы гендердің ұзындығы бірнеше мың жпп нуклеотидтер. Олар ДНҚ-ның ерекше бөлігі. Бұл бөліктің екі жақ шетінде біріне-бірі комплементарлық қарама-қарсы айналған, қайталанған тізбектері болады. Бөліктің ішкі жағында транспозазаны кодтайтын ген орналасқан.
Бактерияларда жылжымалы гендердің ұзындығы мен құрылымы жағынан айырмашылығы бар екі негізгі класы анықталған: 1) инсерциялық тізбектер немесе 15-элементтері, ұзындығы бір мыңдай жұп нуклеотидтер, құрамында ауыспалылыққа жауапты тек бір ғана гені бар; 2) үзындығы 3-20 мжн, күрамында әр түрлі улы заттарға бактериялардын төзімді болуын камтамасыз ететін бірнеше косымша гендері бар.
Жүгері геномынан бірнеше жаксы зерттелген жылжымалы гендер бөлініп алынды, олар Ас, Дз, Ми-1, 8рт символдарымен белгіленеді. Бул гендер өсімдікгердін геномында «мылкау» (кызмет аткармайтын) компоненттерге жататындығы туралы генетикалық және молекулалык дәлелдер алынды. Олар тек кана хромосома үзілген кезде ғана активтенеді. Молекулалык зерттеулер көрсет-кендей, көптеген транспозондарға сайма-сай (гомологиялык) тізбектер жүгері геномында кездеседі екен. Активтенген сон бүл элементтер үрпактан үрпакка түкым қуалайтын өзгергіштіктерге себепшіл болатын спонтанды түраксыздыкты тудыратын мутациялар мен хромосомалардын қүрылымдык өзгерістеріне жауапты болады. Осы түраксыздык геномнын кайта күрылып жанаруында манызды кызмет аткарады және де өсімдіктер популяцияларынын әр алуандығына да ыкпалын тигізеді.
Жылжымалы гендер геном ішінде бір жерден екінші жерге оңай көше алатын болғандыктан, оларды рекомбинанттык ДНҚ-иы тасымалдайтын тиімді вектор ретінде пайдалануға болады. Транспозондар негізінде күрастырылған векторды колданып трансформация процесін ен бірінше рет дрозофила шыбынында өткізген. Р-жылжымалы элементтін көмегімен дрозофилаға коңыр бояуын кодтайтын ген тасымалданды. Транспозондар көмегімен гендерді тасымалдаудың көп артыкшылығы бар. Бөтен ген тіркестірілген сон ДНҚ күрылысы онша өзгермейді. Сонан соң бүл әдіспен ДНК-ньщ едәуір үлкен бөліктерін тасымалдауға болады. Казір өсімдіктер транспозондары жаксы зерттеліп жатыр, сондыктан болашакта олар бөтен гендерді клеткаға тасымалдау үшін генетикалык инженерияда кен колданылуы сөзсіз.
Вирустарды вектор ретіңңе колдану
Клетканын вируспен немесе вироидпен закымдануы, онда жана генетикалык материалдын пайда болуына әкеп соғады.
Әдетте вирустар репликацияланған кезде олардың генетикалық материалының (нуклеин қышқылдарының) өте көп көшірмелері түзіледі. Сондықтан, вирустар және вироидтар бөтен геннің жақсы тасымалдануын, хромосомаға тіркесуін және экспрессиясын қамтамасыз ете алады.
Вирустардың бөтен нуклеин қышқылдарын өздеріне жабыстырып, тасымалдап алып жүретін мүмкіншіліктері мол. Бірақ вирустарды вектор ретінде қолданудың екі кемшілігі бар: олар ауру қоздырады (патогендік) және қожа клетка геномына тіркесе алмайды. Ал вегетативтік жолмен көбейетін өсімдіктерге бұның кедергісі болмайды. Вирусты вектор ретінде пайдалану үшін оның патогендік қабілетін әлсірету немесе мүлдем жою қажет. Сонда ол өсімдікке зиян келтірмейді. Осындай әлсіз вирус штамдары кейде табиғатта өз бетімен пайда болады немесе тәжірибеде мутагенез арқылы алынады.
Вектор ретінде өсімдік вирустарының үш түрін қолдануға болады: 1) сыңартізбекті РНҚ-сы бар; 2) сыңартізбекті ДНҚ-сы бар; 3) қостізбекті ДНҚ-сы бар вирустар. Өсімдік вирустарының бәрінің дерлік (90 % астам) генетикалық материалы сыңартізбекті РНҚ-дан тұрады, яғни мРНҚ-дан. Және де олар жұқпалы ауру туғызады. Кері транскриптаза мен ДНҚ-полимераза ферменттерінің көмегімен сыңартізбекті РНҚ қостізбекті комплементарлық ДНҚ-ға (кДНҚ-ға) айналады. Ал одан кейін кДНҚ-ны бактерия плазмидасына кіргізіп оларды клондап көбейтуге болады. Өсімдікке енгізілетін бөтен ген осы кДНҚ-ға тігіліп клеткаға тасымалданады.
Темекінің сылдырмақ ауруын қоздыратын вирустың (ТСВ) РНҚ-сын вектор ретінде пайдалануға негізделген әдіс жете зерттеліп дайындалған. ТСВ жұқтыратын РНҚ екі типті молекула болып келеді. Олардың әр қайсысы жеке таяқша тәрізді бөлшекте орналасады. Ұзынырақ таяқшаның РНҚ өсімдікте репликацияланады, бірақ вирус қабығын құрастыратын белокты түзе алмайды. Қысқа таяқшадағы РНҚ вирустың қабыкша белогын кодтайды, бірақ оның репликациясы РНҚ-ның тек ұзын фрагменті қоса болғанда ғана жүреді. Вирустың қысқа РНҚ молекуласын бактерия плазмидасында клондап вектор ретінде пайдаланады.
Джеминивирустар тобының генетикалық материалы сыңар тізбекті ДНҚ-нан тұрады. Бұл топқа оншақты вирус жатады. Олардың әр қайсысының өз қожалары (ауру жұқтырытын өсімдіктер) бар. Вирус қабығының ішіндегі ДНҚ көбінесе сақина тәрізді келеді, бірақ кейде линиялық молекула болып та кездеседі. Бұндай ДНҚ-ның екеуі де жұқпалы ауру қоздырады.
Түсті қырққабаттың теңбіл кеселін қоздыратын вирустар (ТҚТВ) өсімдік вирустарының арасындағы генетикалық материалы қостізбекті ДНҚ тұратын жалғыз ғана топ. Бұл топқа 12 вирус кіреді. Олардың әр қайсысының белгілі қожа өсімдігі бар. Денегмен, олар бұршақ тұқымдастары мен даражарнақты өсімдіктерді зақымдай алмайды. Осы топтың ең жақсы зерттелген вирусы - ТҚТВ. Оған қожа бола алатын өсімдіктер саны көп емес (қырққабат, түсті қырққабат, турнепс, брюссельдік қырққабат, рапс, арабидопсис). Табиғатта вирустарды көбінесе өсімдік биттері тасымалдап жұқтырады.
ТҚТВ вектор ретінде пайдалану жөніндегі тәжірибелер екі бағытта жүргізілуде. Біріншісі - ол вирустың репликациясы мен экспрессиясын молекулалық деңгейде зерттеу және маңызды функциялары кодталған ДНҚ бөліктерін белгілеу. Екіншісі -ТҚТВ геномында өтетін делециялар мен инсерциялардың ауру жұқтыру қабілетіне әсерін анықтау.
ДНҚ молекуласына басқа гендер тіркескенде оның ауру жұқтыру қабілеті жоғалмауы қажет. ДНҚ бойындағы сондай нуклеотидтер тізбегінің орындары қазір анықталған, және де енгізілетін бөтен ДНҚ көлемі белгіленген. Тәжірибе дәлелдегендей, сол орынға енгізілген бактерияның ДНҚ-сы өсімдікке тасымалданған. Бұл нәтиже вирустық ДНҚ вектор қызметін атқара алатындығын дәлелдейді. Вирус ДНҚ-на бактерияның дигидрофолатредуктаза ферментін кодтайтын ген тігілген. Бұл фермент метотрексат деген антбиотикке төзімділікті қамтамасыз етеді. Вирустық геномның бұндай модификациясы оның ауру жұқтырушылық қабілетін өзгертпеген. Мысалы, турнепс жапырағынын бетіне рекомбинанттық ДНҚ молекулаларын жаққанда, олар клеткаға еніп, ауру қоздырған. Зақымданған өсімдік клеткаларында вирустардың көбеюімен бірге оларға енгізілген бактериялық ген де көбейген. Соның нәтижесінде өсімдік клеткаларында метотрексатқа төзімділікті белгілейтін дигидрофолатредуктаза ферменті түзілген. Оны өсімдікке метотрексат ерітіндісін бүркіп тексерген. Одан кейін ТҚТВ көмегімен турнепстің бүтін өсімдіктеріне және протопластарына канамицин деген антибиотикке төзімділік гені енгізілді. Бұл тәжірибелерде де вирус арқылы бөтен геннің өсімдік клеткаларына тасымалдануы дәлелденді.
Вирустарды вектор ретінде қолданудың артықшылығы, ол вирустан бөлініп алынған ДНҚ-ны (экстрактың) өсімдік жапырағына жаққанның өзінде-ақ оларды клеткаға кіргізу мүмкіндігі. Одан кейін вирус өз бетімен көбейіп, клеткалар бөлінген сайын бүкіл өсімдікке ауру жұқтырады. Ал егер де трансформация in vitro өсірген клеткаларда жасалса, түбінде бәрі бір регенерация процесін өткізіп, бүтін өсімдік шығару қажет. Бірақ жапырақ протопластарына вирус жұқтыру тиімді жол бола алмайды. Себебі, трансформацияланған клеткалардан регенерант өсімдігі дамыған кезде вирус жоғалып кетуі мүмкін, өйткені вирустың меристема клеткаларында таратылуы шектеледі. ТҚТВ-ның ДНҚ-сы қожа өсімдік геномына тікелей тіркесе алмайды. Сондықтан оны клетка хромосомасына орналастыру үшін арнаулы әдіс дайындалған, ол агроинфекция әдісі. ТҚТВ геномы агробактериялардың Т-ДНҚ-на тігіліп, кейін әр түрлі өсімдіктердің (турнепс, бұршақ, темекі) ядролық геномына енгізілген. Агроинфекция әдісі in vitro өскен өсімдіктерді де трансформациялауға қолайлы келеді.
Вироидтар - өте ұсақ, сақина тәрізді дөңгеленіп оралған, молекулалық салмағы жеңіл сыңартізбекті РНҚ жіпшесі. Қазір 15-ке жуық вироидтар анықталған. Олардың РНҚ құрамына 240-380 нуклеотид кіреді. Вирустардан айырмашылығы, вироидтар белок қабығымен қоршалмаған, өте қысқа дара РНҚ молекуласы. Олардың репликациялану механизмі белгісіз. Жорамал бойынша, вироидтар өсімдік клеткаларында солардың ферменттерін (РНҚ-полимераза) пайдаланып, ядрода вироидтық РНҚ синтезін жүргізеді. Вироидтарда ДНҚ түзілу стадиясы жоқ, сондықтан олар өсімдік геномына тікелей орналаса алмайды.
Вироидтардың ерекшеліктері мен вектор ретінде қолдану мүмкіндіктерін Э. С. Пирузян (1988) былай мазмұндайды:
1) вироидтар қоздыратын ауру тек зақымданған жерге ғана емес, бүкіл өсімдікке жайылады;
2) механикалық жолмен немесе сөл арқылы тасымалданады;
304
3) кейбір вироидтар ұрық немесе тозаң арқылы жұғады;
4) вироидтардың репликациясы өсімдік клеткасының ядросында өтеді;
5) өсімдіктердің кейбір жұқпалы аурулары вироидтарға байланысты.
Сонымен, вироидтарды генетикалық инженерияда вектор ретінде қалай пайдалануға болады? Ол үшін вироидтық РНҚ-ға комплементарлық ДНҚ (кДНҚ) тізбегін қолдан синтездеу керек. Ал кДНҚ репликацияланып екінші ДНҚ тізбегін синтездейді. Сөйтіп, қостізбекті ДНҚ, түзіледі. Одан кейін ол плазмида құрамына тігіліп, Е. соlі клеткаларына енгізіліп, көбейтіледі. Векторлар вироидтық РНҚ негізінде, вироид РНҚ-сында комплементарлы синтезделген ДНҚ, негізінде және де олардың Т-плазмидалармен қосылған құрылымдары негізінде дайындалады. Бөтен ген осылай дайындалған векторларға кіргізіліп өсімдік клеткасына тасымалданады. Вироидтарды вектор ретінде пайдаланғанда бөтен геннің өсімдікте экспрессиясы тоқтаусыз жүре алады.