

6. Сомалық будандастыру цитология, генетика, молекулалық биология, физиология салаларында теориялық мәселелерді зерттеу үшін, сондай-ақ практикалық селекцияда қолданылады.
Сомалық будандастыру селекция үшін өте жаңа технология. Ол жыныстық жолмен будандаса алмайтын өсімдіктердің формалары мен түрлерін будандастыруға мүмкіндік береді, яғни түраралық гибридизация процесін шектейтін генетикалық сыйымсыздықты жеңе алады. Тіпті жыныстық будандастыру мүмкін болған кездің өзінде де, сомалық будандардың жыныстық будандардан өз артықшылықтары бар. Себебі, сомалық будандарда цитоплазмалық гендер ата-ананың екеуінен де тұқым қуалайды. Ал жыныстық будандарда цитоплазмалық гендер тек қана аналықтан тұқым қуалайды.
Сомалық будандарды селекция процесінде пайдалану үшін олар гүлдеуге және тұқым беруге қабілетті болулары қажет, өйткені сорт шығару үшін бірнеше ұрпақ кері қайтара аталық немесе аналықпен будандастыруға түседі. Ұрпақсыз будандардың селекцияға пайдасы жоқ.
Әдетте сомалық клеткалар қосылғанда, алынған буданда хромосомалар саны екі еселенеді. Вегетативтік жолмен көбейетін кейбір өсімдіктерге (картоп, қант қамысы т.б.) хромосомалардың саны өзгергендігі қауіп туғызбайды. Ал көптеген ауыл шаруашылық дақылдары жыныстық жолмен көбейетіндіктен, оларға бұндай хромосомалық өзгерістер зиян болады. Сондықтан сомалық буданында плоидтылығы өзгермеу үшін, гаплоидтық өсімдіктің протопластарын қосу керек немесе сомалық буданның тозанқаптарын іn vitro өсіріп олардан гаплоидтық өсімдік шығарып алу керек.
Сомалық будандастырудың практикалық жетістіктері Мсойапа, Зоіапшп, Оаіига туыстары ішіндегі түраралық будандарды шығарумен байланысты болды. Мысалы, практикалық селекция үшін бағалы темекінің кейбір жабайы түрлері шаруашылыққа құнды №соІіапа ІаЬасит сорттарымен жыныстық жолымен будандаспайды. Ал парасексуальдық будандастыру арқасында ұрпақ беруге қабілетті амфидиплоидтық өсімдіктер алынып, кейін олар селекция жұмыстарында пайдаланылды. Сасықмеңдуананың өнеркәсіпте қолданылатын мол алкалоидты сорттарын шығарудағы селекция жұмыстарында сомалық будандар қолданылады. О. Шидер 1978 ж. сасықмеңдуананың жыныстық будандасуға қабілетсіз түрлерінің протопластарын қосып, нормалы ұрық бере алатын сомалық будандарды алды, және де олардың құрамында сксшоламин алкалоиды 20-25 %-тен артық болды.
Сомалық клеткаларды будандастыру арқылы картоптың селекция үшін бағалы формалары алынды. Мысалы, картоптың екпе түрімен жабайы түрі арасындағы сомалық буданының шаруашылыққа құнды қасиеттері болғандықтан, ол будан селекииялық жұмыстарда пайдаланылды. Бірақ, өкінішке орай, бірнеше жыл бойы будандастыру салдарынан кейінгі ұрпактарда бастапқы жабайы түрдің белгілері басым формалар пайда болған. Әдістің кемшілігі, ол жағымсыз белгілерден құтылудың қиындығы және стерильді ұрпақсыз өсімдіктердің шығуы. Осындай қиыншылықтар сомалық будандастырудың селекцияда кең қолдануын шектейді. Жабайы түрлердің пайдалы гендерін мәдени өсімдіктерге көшіру үшін генаралық рекомбинация өтуі керек немесе белгілі бір хромосома басқа гомологиялық хромосоманың орнын басуы қажет.
Бұл цитогенетикалық процестерге қосымша түсінік бере кету орынды болар. Будандастыру кезінде аталық пен аналық өсімдіктердің геномдары қосылып бір ядрода орналасады да гомологиялық хромосомалар арасында кроссинговер өтеді. Сонда өз бойының әр жерлерімен гомологиялық хромосомалар жанасып, қайтадан ажырасқанда ұзынды-қысқалы бірінің бөлшектері біріне ауысады. Соның салдарынан сол бөлшектерде орналасқан гендер де алмасады. Генаралық рекомбинация хромосомалық кроссинговер негізінде өтеді. Бұл жағдайда хромосомадағы гендердің бірнеше тобы ғана жабайы түрден мәдени өсімдікке көшеді. Солардың қатарында пайдалы геннің де болуы ықтимал. Бірақ кроссинговер кездейсоқ өтетіндіктен, селекционер көздеген нақтылы генді бақылап көшіру мүмкін емес. Соған байланысты будандастыру кезінде қажетті геннің алмасуы да кездейсоқ уақиға болады. Сәтті өткен генаралық рекомбинацияны «мыңда бір» гибридтік ұрпақты талдау арқылы ғана тауып, сұрыптап алуға болады.
Қажетті генді будан клеткаға (организмге) енгізуді қатаң бақылауға да болады. Ол үшін құрамына нақ сол ген кіретін хромосоманы бүтін күйінде жабайы түрден мәдени өсімдікке көшіру керек. Алдымен мәдени өсімдіктің моносомалық линияларын (әр линияда белгілі бір хромосома жетіспейді) жасайды. Мысалы бидайдың Чайниз Спринг деген сортында сондай моносомалық линиялар жасалған. Будандастырған кезде жабайы түрдің керекті хромосомасы мәдени өсімдіктің жетіспейтін гомологиялық хромосомасының орнын басады. Сонда селекционер алдына қойған мақсатқа сәйкес ген будан клетканың (организмнің) геномына енеді, алынған будан жаңа пайдалы қасиетке ие болады.
Д. Эванс мынадай қызықты экспериментті өткізді. Ол темекінің сомалық буданын ата-ананың біреуі ретінде қолданып, жеке гендердің комбинациясын алды (44-сурет). Шыққан будан үш түрдің де генетикалық информациясын иеленді, сондықтан ауруға төзімділіктің жаңа көзі болып, кейін екпе сортымен қайыра будандастырылды.
Ата-ананың біреуінің толық геномын, екіншісінің - тек бірнеше хромосомаларын өзіне дарытқан асимметриялық сомалық будандардын селекция үшін маңызы зор. Бұндай будандарды алу үшін протопластар құйылысқан соң біреуінің хромосомалары жойылып кетуі қажет. Ол үшін сол ата-ананың протопластарына хромосомаларды тұрақсыздандыратын радиоактивтік сәулені түсіру керек. Сәуленің күшіне және әсерлік мерзіміне қарай ядро толық инактивтелуі мүмкін. Егер осындай протопласты кәдімгі пормалы протопластпен қосса, цибрид (цитоплазмалық габрид) шығады. Бұл цитоплазмалық буданда ядролық гендер радиоактивтік сәуле әсер етпеген аталықтан немесе аналықтан І аиа, ал цитоплазмалық гендер екеуінен де немесе біреуінен ғана болады (45-сурет). Асимметриялық будандарды (бір ата-ананын геномы толығымен, екіншісінің геномы жартылай) филогенезде түпкі тектері алыс түрлерді будандастыру негізінде алуға болады. Бірақ бұндай будан клеткалардан құнды ұрпақ беретін өсімдіктер шығуы неғайбыл болғандықтан, оларды маңызды заттар түзетін жана клеткалық линиялар алу үшін жүргізілетін селекция процесінде қолдануға болады.

45-сурет. Протопластар құйылысуының нәтижесінде пайда болатын асимметриялық будандардың әр түрлі варианттары (5).
Аталық ұрықсыздық, кейбір гербицидтерге, токсиндерге төзімділік фотосинтездің тиімділігі тағы басқа белгілер цитоплазмалық гендер арқылы тұқым қуалайды. Парасексуальдық будандастырудың әдістері цитоплазмалық «вдермен айлалы әрекет жүргізуге мүмкіндік береді. Мысалы, бір түрдің оплазмонынын бір бөлігін (пластомды), ал екінші түрдің дитоплазмонының басқа бөлігін (митохондрионды) иеленген будандарды шығаруға болады (46-сурет). Осы жолмен цито плазмон темекі, шырайгул жазушаларына көшірілген. Кызығарлықтай мақсат, осы тәсілді қолданып жабайы тұқымдастарға тән тербицидтерге төзімділікті екпе өсімдіктерге көшіру, мысалы, картопқа, өйткені ол барлық гербицидтерге
сезімтал.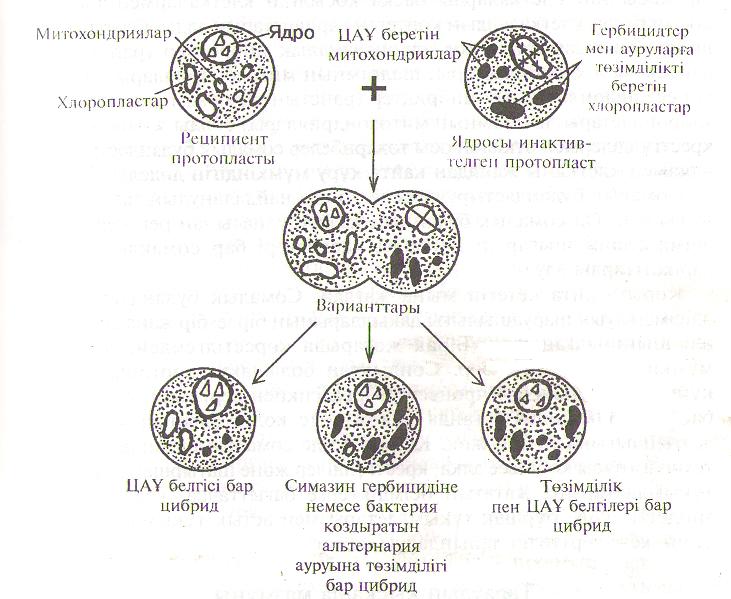
X, Борнман өз тәжірибелерінде рапстын шалғам мен турнепс негізінде шығарылған екі сортын қолданған. Бұл сорттарды алу үшін шалғам мен турнепс цитоплазмалық аталық ұрпақсыздық бслгісінің көзі болған. Ралстың екі сортының, шалғамнын және турнепстың протопластарын, цитопластарын (ядросыз), хлоролластарсыз, протолластарды өзара әр қилы тіркестерде қосып және бір қосылған клеткаларды басқа қосылған клеткалармен тағы қосып будан клеткалардың көптеген варианттарын алды. Олардың ішінде ең қызықтары қос аллоплазмалық цибридтер (рапстың ядросы мен хлоропластары, шалғамның митохондриялары) мен үш есе аллоплазмалық цибридтер (рапстың ядросы, турнепстың хлоропластары, шалғамның митохондриялары) болды. Сонымен, крестгүлділермен өткізген осы тәжірибелер сомалық будандастыру әдісімен клетканы жаңадан қайта құру мүмкіндігін дәлелдеді.
Сомалық будандастыруды селекцияда пайдаланудың тағы бір жолы бар. Ол сомалық буданның каллус ұлпасынан регенерант өсімдіктерін шығарып пайдалы қасиеттері бар сомаклондық варианттарды алу.
Қорыта айта кететін мына жағдай. Сомалық будандастыру әдісімен ауыл шаруашылығы дақылдарының бірде-бір жаңа сорты әлі шығарылған жоқ. Бірақ жоғарыда көрсетілгендей, оның мүмкіншіліктері өте зор. Сондықтан болашақта протопластар құйылысу негізіндегі пролестері іргелілікпен зерттеліп, оларды басқару мүмкіндігі туғанда бұл жұмыс қолға алынып жүзеге асатындығына күмән жоқ. Қазіргі кезде сомалық будандастыру технологиясы көбінесе алқа, крестгүлділер және шатыршагүлділер тұқымдастарына жататын өсімдіктерге бағытталса, келешекте міндетті түрде бұршақ тұқымдастары мен астық тұқымдастары үшін жете зерттеліп дайындалуы қажет.
Дәріс №26
Гендік инженерия – рекомбинанттық ДНҚ-ны құрастыру
Жоспар
Гендік инженерия - молекулалык және клеткалык генетика-ІІІ.Іи колданбалы саласы. Белгілі касиеттері бар генетикалык матсриалдарды (гендерді) іп УІІГО жағдайында алдын ала күрастырып, оларды тірі клеткаға енгізіп, көбейтіп, зат алмасу ІІроцесін өзгеше жүргізу. Бүл әдісп.ен организмдердегі генетикалык информацияны көздеген максатка сай өзгертіп, олардың геномдарын белгіленген жоспармен кайта күруға болады.
Гендік инженерия ол функционалдык активті генетикалык күрьшымдарды рекомбинанттык (будан) ДНК. молекулалары түрінде колдан күрастыру. Гендік инженериянын мәні жеке Іхчідерді бір оргаңизмнен алып баска организмге көшіріп орпаластыру. Бүған рестриктаза меы лигаза ферменттерінін шиылуы мүмкіндік туғызады. Рестриктазалар (рестрикциялык •Іилоиуклеазалар) ДНК молекуласын белгілі жерлерден жеке V Ііиділерге киып бөлшектейтін ыдыратушы фермент. Казір ДНК молскуласын бір-бірінен өзгеше 120 жерінен үзетін 500-ден астам рійс гриктазалар аныкталған. Алынған полинуклеотид бөлшектерінін (ДНК фрагменттерінін) комплементарлык немесе «жабыскыш» үштарын ДНКлигазасы бір-біріне «желімдеп» рет-теп жалғастырып косады. Осы ферменттердің көмегімен бір ДНК молекуласынан кажетті ген бөлініп алынып, баска ДНК, молскуласының үзінділерімен күрастырылып рекомбинанттык, иі ии жаңа будан ДНК жасалады.
Одан кейін рекомбинанттык ДНК бірнеше әдістермен тірі клеткаға енгізіледі. Жана геннін экспрессиясы өтеді де, клетка І'ол гсн белгілейтін белокты синтездей бастайды. Сонымен, клсіхаға рекомбинанттык ДНҚ молекуласы түрінде жана генетикалык ияформацияны енгізіп, акырында жана белгісі бар организмді алуға болады. Бүндай организмді трансгеңцік немесе трансформацияланган организм деп атайды, себебі бір организм-нін өзгеріп баска касиетке ие болуын трансформация дейді.
Сөйтіп, гендік инженериянын дамуына негіз болған молекулалык биология мен молекулалык генетиканын мынадай жетістіктері: 1) рестриктазалар мен лигазалардын ашылуы; 2) генді химиялык және ферменттерді колдану аркылы синтездеу әдістері; 3) бөтен генді клеткаға тасымалдаушы - векторларды лайдалану; 4) бөтен генге ие болған клеткаларды тандап бөліп алу жолдарынын ашылуы.
Алғашкы рет рекомбинанттык ДНК 1972 жылы АКДІ-та Стэнфорд университетінде П.Бергтын лабораториясында жасалды. Онда пробирка ішінде үш түрлі микроорганизмнін ДНК-лары - лямбда фаггын және ішек таякшасы бактериясынын ДНК фрагментгері мен маймылдын онкогендік вирусынын толык геномы косылған еді.
Өсімдіктердін гендік инженериясы саласында бірінші жүмыстар іп үііто өсірілетін клеткалармен 1980 жылы жүргізілген. 1983 жылы алдымен күнбағыстьщ трансгендік каллусы, кейін сол каллустан табиғатта мүлдем болмаған санбин өсімдігі алынды. Санбин (ағ. виппоу/еі - күнбағыс, Ьееп - бүршак) деген ол геномында бүршактьщ белогы фазеолинді кодтайтын гендері бар күнбағыс өсімдіғі еді.
Гендік инженерия гендерді тасымалдау тәсілі ретінде болашакта екпе өсімдіктердін селекциясыньщ тиімді аспабы бола алады. Казіргі кезде гендік инженерия алғашкы кадамдарын басып, екпінді дамып келеді.
Гендік инженериянын әдістемелік негізі жапырактың мезофилл клеткаларынын немесе каллус үлпасының протошіастары болады. Жана генетикалык информацияға ие болған лротопласты өсіріп, одан регенерант өсімдігін алуға болады. Генетикалык трансформация үшін сомалык клеткалардан баска тозан клеткалары, жүмырткаклеткасы колданылады. Сонымен, іn vitro өсірілетін клеткаларға гендік инженерияньщ әдістерін колданып, өсімдіктердін бағалы белгілері бар негізінде жаңа формаларын күруға болады.
Гендік инженериянын жүмысы мынадай кезендерден тұрады:
1) баска организмге көшірілетін күрыльшдык (структуралық) генді алу;
2) оны вектордын кұрамына енгізу, яғни рекомбинанттык ДІНК ны жасау;
3) рекомбицанттык ДНК-ның өсімдік клеткасына тасымадцау;
4) осімдік клеткаларында бөтен ДНҚ-нын экспрессиясын Ішідау;
5) геномы өзгерген жеке клеткалардан регенерант өсімдігін алу.
Басқа организмге тасымалданатын кажетті генді бөліп алу
Врбір полипептид тізбегінін, яғни белоктын, өзінін күрылым-дык гені (структуралык) болады. Ол ген нактылы белок күрамын-шігы амин къ>Ішкылдарынын бір-бірімен ізділігін, жалғасу ретін ЛелІ ілейді. Геіідік инженериянын мақсаты - әрбір дербес күры-ІІымдық генді бір өсімдіктен баска бағалы сортгың өсімдігіне оны Ігдаи әрі жаксарту үшін енгізу.
Кдзіргі уакытта молекулалык биологияишн жетістіктерінін ІІркасында күрыльшдык гендерді таза күйінде және де жеткілікті мклінсрде бөліп шығару әбден болады. Бірак бүл жүмыстын •к-імдіктермен өткізгендегі киыншылыктары, ол өсімдіктердін Існомдарының едәуір күрделілігі. Онын күрамына 150 мьщнан иггам гендер кіреді, ал соның ішінде тек 5-10 % бірегей ДНК Ісио І икалык код кызметін орыңдай алады, яғни 15-25 мын гендер күрмлымдык гендері болғаны. Өсімдіктер геномында функциясы Гюіп ісіз көп кайталанған ДНК элементтері орасан зор (90-95 %). < 'оидғ>іктан, өсімдіктердін бір белгісін кодтайтын жеке гендерін гснссгіру өте киын да ауыр жүмыс. Одан баска, біркатар манызды Гчмплер тек бір генде емес, көптеген гендерде жазылған. Мысалы, омімділік, тез пісіп жетілу, азотты сініру, ортанын колайсыз фіікюрларына төзімділік белгілері полигендік болады. Бірак пилрдын биохимиялык негіздері белгісіз.
Гспдік инженерияның әзірше алға койған максаты - анык бір Іі жазылған карапайым гендермен айналысу. Мысалы, кейбір Пелоктары, гербицидтер мен пестицидтерге төзімділік гендері, І І\с. Әсімдіктердін күрылымдык гендерін бөліп алу үшін гендік инжснсриянын әдістері колданылады.
Гендердін көптеген геномдык көшірмелері ДНК-ның комплементарлык тізбегін (қЦВДС) кері трансқршггаза (ревертаза) көмегімен матрицалык РНҚ-да синтездеу аркылы алынған.
Ревертаза көмегімен уйлесімді иРНК болса, көрінген дербес генді синтездеуге болады. Ал иРНҚ-ны бөліп алу әдістері жаксы дайындалған.
Будан баска, белоктын алғашкы курьшымын зерттеу әдістері-нің жетілдіруі аркылы сол белокты кодтайтын генді химиялык-биологаялык жолымен синтездеуге болады. Соньшен катар, ол ушін ДНК-нын нуклеотид калдыктарынын ізділігін тура аныктауға (секвенирование) колдануға болады.
Кдзіргі уакытта әр турлі өсімдіктердін 100-ден астам курылым-дык гендері бөлініп алынған және жан-жакты талданған. Бул гендерге кор белоктарыньш, турлі ферменттердін гендері, ыстык-пен немесе суыкпен, анаэробтык жағдаймен немесе патогендік микроорганизмдермен индукцияланатын гендер жатады. Өсім-діктер курылымдык гендерінін бір бөлігі курамы әр турлі мульти-гендер (көпгенділік). Мысалы актин, гистон, леггемоглобин, буршак және астык тукымдастарынын коры ретівде жиналатын кейбір белоктарынын гендері.
Кор белоктарынын молекулалык салмағы және компоненттер саны бойынша едәуір линияаралык полиморфизмі болады. Олардьщ курылымы жөніндегі информация ДНК деңгейінде де сакталады. Мумкін, бул полиморфизм жылжымалы генетикалык элементтерге (транслозондарға) байланысты болар.
Сонымен, кажетті белгіні кодтайтын гендерді бөліп алу киьшдығы, олардын бірегейлігіне, геномда сирек кездесуіне байланысты немесе олардьщ топтасып (кластер курып), хромосомаиьщ әр турлі жерінде орналасуына байланысты.