

5. Өсімдік шаруашылығында биотехнологияны қолдану негізгі екі бағытта жүргізіледі:
селекция , яғни трансгендік өнімдіктерді алу,
тұқым шаруашылығында, сорттарды сауықтыру
Гендік инженерия арқылы трансгендік өнімдіктерді алу биотехнология саласының қауырт дамып келе жатқан бағыттарының бірі болып отыр.
Қазіргі кезде трансгендік дақылдардың дүние жүзі бойынша жалпы егіс көлемі 83,7 млн. ие құрап отыр. Трансгендік дақылдардың егіс көлемінің басым бөлігін май, бұршақ, жүгері, мақта, рапс дақылдары алуда. Өсімдіктердің трансгенозын қолдану дүние жүзінде жылына 105 % жылдамдықпен жүргузілуде. Өйткені, трансгендік дақылдарды өсіру арқылы ауылшаруашылық өсімдіктерінің өнімділігі 22 пайыздан 49 пайызға дейін жетеді. Бұл әрине өте үлкен жетістік.
Тұқым шаруашылығында биотехнология жетістіктерін пайдалану арқылы дақыл сорттарын сауықтыру және олардың сыртқы орта ықпалдарына төзімді (ыстыққа , суыққа, топырақ ортасына және тағы басқа) жаңа сорттарын алу Қазақстан Республикасында 1995 жылдардан бері қолға алынып келеді. Өйткені, тұқым шаруашылығында биотехнологияны қолданып, дақыл өнімділігін 15-30 пайызға дейін жоғарылатуға болады.
Оңтүстік Қазақстан облысы жағдайында биотехнологияның соңғы жетістіктерін өсімдік шаруашылығына енгізу жоғарыда көрсетілген екі бағытта да жүргізілуі қажет.
Гендік өзгертілген өнімдік ретінде Республикамыздың ақ алтыны мақта дақылына қолдану арқылы өсімдіктердің ыстыққа ауа-райының тез өзгергіштікке төзімділігін арттырап, олардың ауруларға төзімділігін жоғарылатады және ең бастысы олардың өнімділігін 2-3 есеге дейін жоғарылатады.Мысалы, трансгендік сорттарды пайдалану арқылы Индияда мақтаның өнімділігін гектарына бес жылғы орта есеппен 58,7 центнерге жеткізген, ал біздегі өнімділік не бары 21,1 ц/га ғана.
Биотехнология жетістіктерін тұқым шаруашылығында қолдану өңірімізде 2001 жылдан бері қолға алынып келеді. Ақтүйнектің тұқым шаруашылығында биотехнелогияны қолдану арқылы дақыл өнімділігін 287 ц/га дейін жеткізіп, сорттарды жаңарту тізбегін 5 жылдан 3 жылға дейін қысқартуға қол жеткізілді. Осындай нақты жетістіктерге жету жолында болашақта халық шаруашылығында кеңінен сұранысқа ие жүзім, май бұршақ, қант қызылшасы сияқты дақылдарға да қолдану қажет.
Бақылау сұрақтары:
Өсімдік биотехнологиясының теориялық және технологиялық негізі не?
Өсімдіктер іn vitro жағдайында оқшауланған ұлпаларды қандай мақсатта қолданылады?
Өсімдік жасушалары мен ұлпаларын өсірудің қанша кезеңі бар?
Қазақстанда іn vitro жағдайында өсімдіктердің зерттеулерінің басталуы?
іn vitro нысандары туралы не білесіз?
Өсімдіктен оқшаулап алынған бөліктерін өсірудің қандай тәсілдерін білесіз?
Регенерант өсімдіктерді алудың қандай жолдары бар?
іn vitro жағдайындағы жасушаларды өсірудің қанша бағыты бар?
Дәіріс №3,4
Іn vіtro жағдайында өсірілетін өсімдік клеткаларынын ерекшеліктері
мақсаты: Өімдік жасушалары мен ұлпаларында болатын өзгерістерді анықтап білуі үшін студенттерді табиғаттағы тіршілік формаларымен, өсімдіктер жасушасының жануарлар жасушасынан айырмашылығымен, құрылысымен, органоидтарының қызметтерімен таныстыру.
Жоспар:
1. Табиғаттағы тіршілік формалары
2. Өсімдіктер жасушасының жануарлар жасушасынан айырмашылығы
3. Жасушалардың құрылыс ерекшеліктері
4. Жасуша органоидтарының қызметтері мен кұрылысы
1. Қазіргі кездегі тұжырым бойынша тіршіліктің 2 формасы белгілі:
Тіршіліктің жасушасыз формасы - оған вирустар жатады. Вирустар ете кішкентай, тіпті жай микроскоп арқылы көрінбейтін, денелер. Олар нуклеин қышқылдарынан және белоктан тұрады. Олардың тіршілігі тек жасушаға енгеннен кейін ғана байқалды, ал өз беттерінше оларда тіршілік құбылыстары байқалмайды. Вирустарды 1892 ж. орыс ғалымы Д.И.Ивановский ашқан.
Тіршіліктің жасушалы формасы. Оның 2 түрі белгілі, а) прокариотты жасушалар (бактериялар, көк-жасыл балдырлар) - түйіршіктелген, цитоплазмадан қос қабат мембрана арқылы шектелген, ядросы болмайды түқым қуалаушылық материялы ретінде сақиналанған ДНҚ кездеседі; рибосомадан баска органоидтары болмайды; мөлшері жағынан өте үсакболып келеді 0,1-0,5 мкм; митоз кездесггейді.
Эукариотты жасушалар-түйіршіктелген, цитоплазмадан қос қабат мембрана арқылы шектелген, ядросы болады; тұқымқуалаушылық материалы ретінде хромосомалар кездеседі, барлық органоидтары болады; клетка мөлшері біршама ірі болып келеді-15-65 мкм., митоз жолымен бөлінеді. Эукариоттарды бір жасушалы ағзалар жасушасы және көп жа-сушалы ағзалар жасушалары деп жіктейді, Ал, соңғыларын өсімдіктер және жануарлар жасушалары деп бөледі.
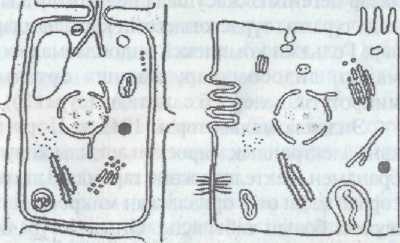

а б в
1 сурет. Жасуша түрлері
а прокариотгар; б эукариоттык өсІмдік жасушасы; в эукариогтық жануар жасушасы;
Бір жасушалы ағзалардың жасушасы (қарапайымдылар) - өздері бір жасуша болып тұрып, тұтас ағзаға тән қызметтер атқарады: қозғалу, тітіркену, көбею, бөліп шығару, ас қорыту.с.с. Ал көпжасушалы ағзалардың жасушалары белгілі-бір қызмет атқаруға маманданады, оларда әртүрлі белоктар синтезделінеді, мысалы; эпителий ұлпасының жасушаларында-мелонин, бұлшықет жасушаларында-миозин т.б.
Өсімдіктер жасушасының жануарлар жасушасынан ерекшелігі мынадай:
1) жасуша сыртын калың целлюлоза кабығы қаптап тұрады;
2) цитоплазмада пластидтер (хлоропластгар, хромопластгар, лейкопласттар) кездеседі;
3) вакуолялары болады.
Қазіргі деректер бойынша жасушаның негізгі заты болып оның тірі заты -протопласт саналады. Протопласт цитоплазмаға және ядроға жіктеледі. Ол сыртқы ортадан шеткі мембрана - плазмолемма арқылы шектелген. Цитоплазма өз кезегінде гиалоплазмаға (цитоплазманың негізгі заты - матриксы) және органеллаларға жіктелген.
Гиалоплазма 2 мембранамен (плазмолемма, тонопласт) шектелген қоймалжың сұйықтық. Ол органикалық және бейорганикалық заттардан тұрады. Оның 80-90 пайызын су құрайды. Органикалық заттардың ішінен негізгілері-белок, нуклеин қышқылдары, майлар, көмірсулар, АТФ т.б. Гиалоплазмада органеллалар бытыраңқы орналасады. Органеллалар дегеніміз-жасушада белгілі-бір қызмет атқарып, цитоплазмада тұрақты түрде кездесетін кұрылымдар. Оларға — митохондриялар, Гольджи комплексі, эндоплазмалықтор, пластидтер, рибосомалар, лизосомалар, жасуша орталығы, микроденешіктер, микротүтікшелер т.б.с. жатады.
Эндоплазмалықторды 1945 ж. Портер ашкан. Ол өте ұсақ, тек қана электрондық микроскоп арқылы көруге болатын, қос қабат мем-бранамен шектелген және тарамданып гиалоплазманы өне бойына торлап тесіп өтіп орналаскан микроарнашыктар мен микроқуыстар жүйесі болып табылады. Оның 2 түрі белгілі: гранулалы немесе кедір-бұдырлы және агранулалы немесе тегіс эндоплазмалық тор. Гранулалы (кедір - бұдыр) эндоплазмалық тордың мембранасына рибосомалар бекінген, ал тегіс эңдоплазмалық торда рибосомалар болмайды. Цитоплазмада, әдетге гранулалы тор агранулалы торға қарағанда әлдеқайда жақсы жетілген. Агранулалы тор кейбір ерекше қызмет аткаратын, яғни майлы заттарды көп синтездейтін жасушаларда ғана жақсы жетілген. Гранулалы тордың қызметі-белок синтездеу, жасуша мембраналарын пайда ету орталығы болып саналады. Сол сияқты, ол вакуоля, лизосома, микроденешіктерді де пайда ете алады. Эндоплазмалық арналар арқылы макромолекулалар, иондар тасымалданады. Агранулалы тор липофильдік заттарды синтездеуге қатынасады.
Гольджи комплексін 1878 жылы италян ғалымы Гольджи жану-арлар жасушасынан ашқан және соңғы кездерге дейін ол тек жануарлар жасушаларына ғана тән деп келінген. Бірақ, кейінірек ол өсімдіктер жасушаларында да табылған. Сондықтан Гольджи комплексі барлық эукариотты жасушаларға тән органелла болып саналады. Гольджи комплексі өте жұқа, жалпақ, бірінің үстіне бірі орналаскан. 5-20 калташықтардан-диктосомалардан құралған. Әрбір қалташықтың диаметрі 1 мкм, ал қалыңдығы небәрі 20-25 нм болып келеді. Қалташыктардың жиектері тесіліп бірте-бірте торға айналған. Гольджи комплексінің кызметі-полисахаридтерді синтездеу, жинақтау және тасымалдау болып саналады.
Митохондриялар жасушаның міндетті органеллаларының бірі. Оның пішіні тұрліше болып келеді: таяқша тәрізді, дөңгелек, со-пакша т.с.с, ал мөлшері 0,5-7 мкм тең. Митохондриялар қос кабат мембранамен шектелген. Сыртқы мембранасы тегіс, тұйық, ал ішкі мембранасы митохондрияның ішіне қарай қатпарлар пайда етеді.
Оларды кристер деп атайды. Кристер арасында митохондрияның негізгі заты-матриксі орналасқан. Онда ДНҚ, рибосомалар, белоктар т.б. кездеседі. Митохондриялар органикалық заттарды ыдырату, АТФ синтездеу қызметтерін атқарады.
Рибосомаларды 1955 ж. Палладе ашқан. Ол екі бөлшектен (кіші бөлшегі, үлкен бөлшегі) тұрады. Олар өте ұсақ, тек электрондык, микроскоп арқылы көруге болатын органеллалар. Оның мөлшері небәрі 15-35 нм болады. Рибосомалар р-РНК-дан және белоктан тұрады, оның негізгі қызметі белок синтездеуі болып саналады.
Лизосомалар - диаметрі 2 мкм, әртүрлі ферменггерден тұратын көпіршіктер. Олар органикалық заттардың гидролиздену процессіне қатынасады, яғни жасуша ішілік ас қорыту қызметін аткарады.
Жасуша орталығы - жануарлар жасушаларына тән органелла. Ол 2 центриолядан тұрады. Әрбір центриоля диаметрі 150нм, ұзын-дығы 300-500 нм болып келетін куыс цилиндр. Оның қабырғасы үш - үштен 9 топқа топтасқан 27 микротүтікшелерден құрылған/ Жасуша оратығының қызметі митоздың калыпты жүруін кдмтама-сыз ету яғни хроматидалардың ажырасуын қамтамасыз ету болып табылады.
Микротүтікшелер - түрліше болып келетін үзын түтіктер, оның диаметрі 24 нм тең.
Пластидтер тек өсімдік жасушаларына тән органеллалар. Олар-дың 3 түрі белгілі: жасыл пластидтер-хлоропластар (Компаретти, 1791 ж.) сары, қызыл пластидтер - хромопластар (Берцелиус 1837 ж.), түссіз пластидтер-лейкопластар (Крюгер, 1854 ж.).
Цитологияның соңғы кездердегі ең маңызды жетістіктеріне мыналарды жатқызуға болады:
1) Цитоплазманың мембраналық құрылыс принциптерін тұжы-рымдау;
2) Жасушаның ашық биологиялық жүйе екендігін тұжырымдау, яғни заттар, энергия және ақпарат ағындары туралы тұжы-рымның калыптасуы;
Цитоплазма және оның органеллалары биологиялық мембрана-лардан тұрады, оның қызметі, касиеттері сол мембраналарға байланысты болады. Шынында да, цитоплазманың құрғақ затының 90 пайызын биологиялық мембраюялар құрайды; гиалоплазма 2 мембранамен (плазмолемма, тонопдртг) шектелген; органеллалар да мембраналардаи тұрады.
Биологиялық мембрана өте жұқа, қалындығы небәрі 5-10 нм болып келетін қабықша. 1972 ж. С.Сингер мен Г.Никольсон үсын-ған биомембрананың сұйықтық-мозайкалық моделіне сәйкес, ол 2 биополимерден тұрады: белок және липидтер(сурет 3). Липидтер молекуласы биологиялық мембраналардың қаңқасын құрайды, ол екі қабат болып орналасып, сұйық фаза күйінде болады, ал оның бетінде немесе оған еніп, кейде оны түгел тесіп етіп белок молекулалары орналасқан. Биологиялық мембраналар жартылай еткізгіштік (таңцамалы өткізгшпік) қасиетке ие, оның ұштары үнемі тұйықталған. Биологиялық мембраналардың арқасында жасушада көптеген дербес, тұйық куыстар (органеллалар) түзіледі. Мембраналардың аркасында осы қуыстарда тек өздеріне ғана тән химиялық құрамы қалыптасып, бір мезгілде түрліше химиялық реакциялардың жүруіне мүмкіндік туады.
Бақылау сұрақтары:
Табиғаттағы тіршіліктің қанша формасы бар, олардың айырмашылығы қандай?
Эукариотты және прокариотты жасушалардың ерекшелігі қандай?
Өсімдіктер жасушасының жануарлар жасушасынан айырмашылығы қандай?
Жасуша органоидтарының атқаратын қызметі?
Биологиялық мембраналардың маңызы неде?
Жасушаның химиялық құрамы неден тұрады?
Дәріс № 5,6 Клеткалардың қоректенуі және қоректік орталардың жалпы сипаттамасы
Мақсаты: Студенттерді өсімдік жасушалары мен ұлпаларын өсірудегі қоректік ортаның маңызымен, құрамымен, қоректік орта құрамына кіретін заттардың ерекшелігімен, қоректік ортаны дайындау әдісімен таныстыру.
Жоспар:
1. Қоректік ортаның құрамы
2. Қоректік орта құрамына кіретін заттардың маңызы
3. Қатты қоректік орта дайындау
1. Өсімдік клеткаларын өсіруге арналған қоректік ортаның құрамы өте күрделі. Ол құрамға минералды тұздар, көмірсулар, амин қышқылдары, витаминдер, түрлі өсуді реттегіштер кіреді. Әр өсімдік ұлпасы өсуі, сонымен қатар бір өсімдіктің, түрлі мүшелері және ұлпаларының өсуі үшін де қоректік ортаның, белгілі бір құрамын талап етеді. Пайда болған каллусты одан әрі өсіру үшін, сонымен қатар органогенез процессін қоздыру үшін де, қоректік ортаның құрамын өзгертіп отыру қажет. Зерттеуші өз жүмысы барысында зерттелетін обьектіге сәйкес қоректік ортаны жетілдіруге талаптанады, сол себептен қоректік ортаның саны да, түрлері де көп.
2. Қоректік ортаның құрамына минералды тұздар макро және микроэлементтер түріде кіреді. Көптеген қоректік орталар құрамына азотнитрат немесе аммони тұздары, фосфор-фосфат, күкірт сульфат түрлерінде, ал темір анорганикалық тұздар және хелат түрінде қосылады. Барлық қоректік орталар құрамында сонымен қатар К+,Са2+ және Ме2+ иондары болады, бірқатар микроэлементтер кіреді. Микроэлеметтер әсіресе сұйық қоректік ортада өсетін суспензиялық клеткаларға аса қажет.
Ұлпалар мен өсімдік жасушаларын өсіру орталарының компоненттерін (жинақтарын) 6 негізгі топқа бөлуге болады, бұл әдетте макроэлементтерді, микроэлементттерді, темір көзін, витаминдерді, көміртегі көздері мен фитогармондардың концентрлі енртінділерін дайындау ретіне байланысты болады.
Өсімдік экспланттарын культивирлеу үшін барлық қоректік орталарға минералды тұздар қоспалары негіз болып табылады.Бұл азот қоспасының нитрат, амони тұздары, нитрит түрінде,фосфор-сульфат түрінде; сондай-ақ ертілетін тұздар K+,Na+, Ca++, Mg ++ -болып келеді.Темір хелат түрінде [FeO4 немесе Fe2O4 +ЭДТА (этилендиаминтетрасірке қышқылы) қолданылады немесе ас тұзы NaЭДТА(трилон-Б)] өсетін ұлпалармен игеріліп кетуіне мүмкіндік береді.
Азот, фосфор, күкірт органикалық қосылыстардың яғни ақуыз, май, нуклейн қышқылдар құрамына жатады. Темір, цинк, марганец, молибден, кобальт порфириндер қасында тотығу-тотықсыздану ферментттері (каталаздар, периоксиездар, полифенолоксидаздар) фотосинтез макромолекулаларын құрайды.Осыған орай,барлық қосылыстар жасушалар мен ұлпаларда структуралы функциялы орындайды.Сонымен бірге K+, Na+, Ca++, Cl-, H+ иондары ортаның рН реттеуге және жасушалардың физиологиялық градиенттерін (тургорды,полярлықтың осмотикалық қысымын) ұстап тұру үшін қажет.
Биологиялық макромолекула үшін, сондай-ақ гетеротрофты ұлпаларды культивирлеудегі көміртегі көзі ретінде (каллустың және суспензияның) қоректік ортаға 20-60г/л концентрлі көміртектерін қосады. Әдетте бұл дисахаридтер(сахароза), моносахаридтер(гексоздар:глюкоза және фруктоза,кентонздар: ксилоз бен және т.б.).Полисахаридтерді қоректік орталарда қолдана бермейді.Тек гидролитті ферменттерді құрайтын кейбір ұлпалар түрін (ісікті) крахмалмен, целлобиозы бар орталарда ғана өсіреді.
Биохимиялық реакцияларға жағдай жасау үшін жасушада витаминдер тобын (В1,В6,В12) С (аскорбин қышқылы) РР(никотин қышқылы), мезоннозиттер сияқты биологиялық катализаторды қолданады.
Тиамин (В1)пируватексрбоксилаз құрамына кіреді,және көмірсуларына айналу барысына қатысады.Тиаминпирофосфат кетоқышқылының тотықтырғыш декарбоксилирлену ферменттер құрамына кіреді және транскетомездар кофертенті болып табылады.
Пиродоксин (В6)фосфорқышқылды эфир түрінде ферменттің декарбоксилирлеуі мен аминқышқылдарының аса аминирлену құрамына кіреді.
Никотин қышқылы (РР) амид түрінде катализденетін донорлы-акцепторлы тізбе Н+(органикалық заттар молекуласынан Н+алу),НАД және НАДФ дегидрогенез құрамына кіреді.
Ұлпалар культурасындағы түр түзуші процестерді дамыту үшін биологиялық даму реттегіштері мен фитогармондардың жетілулері қажет.Бұл заттар жасушалар мен ұлпалардың дедифенциясына және дифференциясына әсерін тигізеді,гистогенезді қорғайды, жасушалардың созылуын және жіктелуді индуцирлейді, жетілу мен қартаю процестеріне қатысады, немесе жағдай жасайды болмаса дамушылық қабілетін және жасушалы культураның жетілуін ингибирлейді,жыныстық қалыптасуды шарттастырады. Биотехнологиялық ізденістерде көбінесе ауксиндер, цитокининдер және гиббнрилиндердің дамуы мен жетілуіне жағдай жасайтын гармондарды пайдаланады.
Ауксиндер: ИУК-β индолил-3-сірке қышқылы, ИМК-индолил-3-май қышқылы,НУК-α-нафтилсірке қышқылы, 2,4-Д-2,4-дихлорфеноксисірке қышқылы.
Цитокининдер кинетин-6-фурфуриламинопурин, зеатин, NH-дифенилмочевина, 6-БАП-6-бензиламинопурин, 2-изопентениладенин(2 ір).
Гибберилиндер: гибберелді қышқыл.
Алғашқы каллус индукциясы үшін қажетті биологиялық қоспа ретінде (ортаның жалпы көлемінен 10-15%) өсімдік экстрактын;кокос сүтін (кокос жаңғағының сұйық эндоспермі)зеатин және кинеин цитокининдерді құрайтын пісіп жетілмеген жүгерінің дәнінен шығатын (ең жақсы сүт кезінде) ( орнын басатын 6 аминопурининдер) және N-дифенилмочевинаны қолдануға болады.In vitro культурасында сұйық және қатты орталарды қолданады.Сұйық орталар суспензияларды, каллусты, оқшауланған денелер мен ұлпаларды регенерант өсімдіктерін культивирлеу үшін пайдаланады. Бұл үшін пробиркалаларға синтетикалық сілтілеу материалдарды немесе сүзгіш қағаздардан арнайы ұстап тұрғыш орналасады.
Қатты (агарланған) ортаны 5,6-6-рН-ында сумен гелді құрайтын теңіз балдырлары құрамына кіретін агар-агармен-полисахарид негізінде дайындайды, ал кейде тығыздығын және орыналмасчтырғыш ретінде Р10,Р100(биогель)полиакриамильді гельдерін қолданады.
Жасанды қоректік опрта үшін қажетті макро және микро тұздар алдын-ала дайындайды және бірнеше рет пайдаланады.Бұл концентрлі ертінділер. Оларды арнайы орындарда:макро және микро тұздарын 0+40С мұздатқышта тесік жабығы бар ыдыста сақтайды, витаминдерді, фитогармондарды, ферментттерді өсінді экстракталарын 200С-та жабыны бар 5-10 мл ыдыстарда сақтайды.(пеницилин флокондары)
Макро тұздардың концентрлі ертінділері.Әдетте жұмнатқарушы концентрациясы бойынша 10-40 есеге,микротұздар 100-1000 есеге,витаминдер -1000 есеге асып түседі.
Фитогармондар ертінділерін қоректік орталармен бірге жұмыс алдында дайындаған дұрыс болады.
Макро және микро тұздардың концентрлі ертінділерін дайындау үшін әрбір тұздарын және стаканға салып ысыту барысында ерітеді, одан соң құйып алып қажетті бір белгіге дейін жеткізеді.Суытылған микротұздар қоспасына молибден тұзының ертіндісін қосады.(тұнба түзуіне мүмкіндік бермеу үшін).
Темір хелатының және кальций хлоридінің концентрлі ертінділерін (күкірт қышқылды темір+ЭДТА, немесе ЭДТА-трилон-Б)басқа тұздардан бөлек сақтап дайындайды.
Витаминдердің концентрлі ертінділерін келесідей тәртіппен дайындайды:10 еселі өлшенген затты 10 мл дистильденген суда әрқайсысын бөлек-бөлек ерітіп алады.
Фитогармондар-бұл суда нашар еритін зат.Сондықтан 100мг затты алдын-ала ащдаған мөлшердегі (0,5-1,2 мл)спиртте (ауксиндер мен гибберилиндер),0,5н HCl немесе KOH (цитокининдер),одан соң (абезинді қышқыл мен кинетининнен баққаны)толық ерігенге дейін қыздырады және көлемін 100 мл көлемге дейін жеткізеді. (1мл-1мг затты құрайды)
Жасушаларды, ұлпаларды және өсімдік денелерін культивирлеу үшін әр түрлі құрамдағы қоректік орталар қолданылады.Көбінесе Мурасиге-Скуга,Уейт,Гамборг (В5)орталары кеңінен қолданылады.
Ұлпалар жасанды қоректік ортада гетеротрофтар болғандықтан, оларға қоректену үшін көмірсулар кажет. Көмірсулардың және энергияның көзі ретінде негізінен концентрациясы 20-40 г/л сахароза, кейде глюкоза қолданылады. Өсімдіктердің кейбір түрлері жасанды қоректік ортада қанттардың басқа түрлерін пайдаланғанмен, әдетте олардың қажетке жарауы шамалы.Көміртегі көзі ретінде бірқатар органикалық қышқылдар мен спирттер сынақтан өткізілген, бірақ олардың жарамдылығы көмірсуларға қарағанда төмен болған.
Қоректік орталардың барлығының құрамына дәрілік витамиңдер кіреді.Витаминдердің ең маңыздылары деп, В тобына жататындары есептеледі. Олардың ішінде: тиамин /В,/, рибофлавин /В^/, пиридоксин /В,/. Көптеген өсіруге алынған клеткаларға бұлардан басқа никотин кышқылы, фоли, иантотен қышқылы, мезоинозит қажет.
Көптеген ұлпалардың жасаңды қоректік ортада витаминдер синтездеуге мүмкіндік болғанмен, зат алмасу кабілетін толық көрсету үшін ол мөлшер жеткіліксіз. Сондықтан қоректік ортаға витаминдерді қосымша беру ұлпалардың өсуін ынталандырады.
Қоректік ортаның құрамы қандай күрделі болса да, оған ен қажет компоненттердің бірфитогормоңдар, себебі бұл қосылыстар өсімдіктерде жүретін физиологиялық процесстерге қатысады.Өсімдіктер клеткалары өсуі мен дифференциялануы үшін ауксиндер мен цитокининдер керек. Өсуді реттегіштер жоқ қоректік ортада тек ісік ұлпалары және қалыптасқан ұлпалар өседі.
3. Қатты қоректік орта дайындау үшін, гель түзетін зат ретінде агар-агар пайдаланылады. Агар-полисахарид, құрамында көптеген қоспалары азот, минералды элементтер, витаминдер бар, сондықтан ағын судың астыңда 4-6 сағат жуу қажет. Өсіруге алынатын ұлпалардың қажетіне қарай қоректік ортадан агар-агардың мөлшері 0,8 ден 1,5 % -ке дейін болады. Ең алғаш өсімдіктердің ұлпаларын жасанды қоректік ортада өсіру агарланған қоректік ортада жақсы нәтиже берген. Бұндай табысқа 1930 жылдары француз Р.Готре және американдық ғалым Ф.Уайт ие болған. Қазіргі уақытта жер шарының кейбір лабароторияларында сәбіздің тамыр сабағынан 1938 жылы Р.Готре алған клеткалары штаммдары сақталған. Каллустың клеткаларын қанша ұзақ уақыт болса да өсіруге болады, тек оларды әрбір 3-4 аптадан кейін жаңа дайындалған қоректік ортаға кайта отырғызып отыру қажет. Осы тәріздес ұзақ, қайта алмастырылып отырылатын қоректік ортада клеткалар мен каллус ұлпалары өсіріледі.
Өсімдік клеткаларын өсірудің техникасының дамуының нәтижесінде бөлініп алынған ұлпалар мен клеткаларды суық қоректік ортада өсірудің әдістері жете зерттелді. Клеткаларды сұйық қоректік ортада өсіру немесе суспензияда өсіру деп, қалқыған клеткаларды немесе кішігірім клеткалар тобын аппаратура арқылы таза ауамен канықтырып және араластыра отырып сұйык қоректік ортада өсіруді атайды. Сұйық ортада клеткалардың таза ауамен канықтырылуының әр түрлі тәсілдері бар: ыдысты тоқтаусыз айналдыру немесе шайқады тағы бір жолы сұйық ортаға залалсыздандырылған ауа үрлеу арқылы. Клеткаларды сұйық қоректік ортада өсірудің өзіндік артықшылыктары бар: олар қоректік заттармен жақсы қамтамасыз етіледі және токсинді заттары тез бөлініп шығады. Ұлпаларды қатты немесе сұйық орталарда болсын өсіргенде, ортаның рН-ы мадызды роль атқарады, ұлпаларды өсіруде жетістікке жету үшін орта рН-ы қолайлы және тұрақты болу керек.
Макромолекулалардың активтілігі мен кұрылымы ортаның рН-на тәуелді, әсіресе ұлпаның құрамындағы ферменттік белоктарға әсері жоғары. Сонымен бірге, ортаның рН-ы қоректік ортаның компоненттерінің тұрақтылығымен оларды клеткалардың қабылдап алуына, әсіресе есуді реттегіштер мен витаминдердің сіңуіне әсер етеді. Егер ортаның рН-ы төмен болса, агар қатпайды, қоректік ортаның рН-ы белгілі деңгейге дейін, қыщқыл немесе сілті қосып жеткізіледі. Көптеген ұлпаларды өсіргенге орта рН-ы 5,5 - 5,8 тең болу керек.
Қоректік орталарды дайындағанда және клеткаларды өсіргенде кейбір физикалық факторларды ескеру керек, мысалы: қоректік ортаның осмостық қысымы, температура және жарық. Жоғары семостық ыдысын қоректік заттардың сіңуін қиындатады. Температура метаболизм процесстеріне әсер етеді, көптеген өсімдіктерді өсіру үшін ең қолайлы температура 24-27 градус. Әдетте үлпаларды қараңғыда немесе шашыраңқы жарықта өсіреді. Тура түскен жарық өсуді бәсендетеді. Хлорофилі жоқ ұлпаларға жарықтың тигізетін әсері фитохром жүйесіне байланысты. Жасыл ұлпалардың өсуі мен морфогенезіне жарықтың тигізуіне» әсерін тек фотосинтездің активтілігімен байланысты деп түсінбеу керек, себебі, ұлпаларды қоректік ортада өсіргенде оларда жүретін фотосинтез қарқындылығы өте төмен.
Дәріс №7,8
Каллусты алу жəне оны өсіру. Каллустың пайда болуына əсер ететін факторлар.
Жоспар: 1. Каллу туралы жалпы түсінік
2. Бүтін өсімдіктердің өсуі
3. Дифференциялану.
4. Морфогенез процестерінің жүру принцптері
Кез келген өсімдіктің тірі ұлпасын қолайлы /лайықты қоректік заттар мен гормондары бар/ ортаға отырғызса, одан маманданбаған клеткалар колониясы пайда болады, оны каллус деп атайды. Каллус-қоректік ортадағы клеткалардың ретсіз бөлінуінің нәтижесінде пайда болған ұлпа. Бұрыннан бар клеткалардың көбейуінен жаңадан клеткалар мен ұлпалар тузілуін пролиферация деп атайды. Сөйтіп каллус пролиферацил нәтижесінде шығады.
Жасанды қоректік орта жағдайында, яғни іп vitro жағдайында өсімдіктердің барлық ұлпалар типтері клеткалары каллус клеткаларына айналады. Бұл процесс дифференциялану деп аталады, оның есебі фитогормондарға байланысты. Қөректік ортаның құрамын өзгерте отырып, ретсіз өсіп жатқан ұлпалардан регенерант- өсімдік алуға болады. Сонымен бір клеткадан тұтас бір өсімдікті өсіріп алуға болады, яғни өсімдік клеткалары ерекше касиетін – тотипотенттілігін көрсетеді.
Каллус ұпаларының әр типті морфогенезге икемді болғандықтан оларды көптеп өсіреді. Каллустың түзілуі мен өсуін ауксиндер мен цитокиниңдер реттеп отырады. Көптеген жағдайда каллус түзілуі /индуюдиялау/ үшін кейінгі өсуге қарағанда ауксилдерді 10 есе артық қажет етеді. Каллус клеткалары өсу қарқындылығымен ерекшеленеді, түсі арқылы /ақ- ақшылдан -қара қоңырға дейін, тығыздығына қарай /бос- борпылдақ, тығыз, т.б. түрлері , жарықта түсін өзгертіп көгеретіндігімен тағы сол секілді белгілерімен бір-бірінен ажыратылады. Каллус ұлпаларды 3-4 апта өткеннен кейін, коректік ортадан шығарып алады да, бірнеше бөлікке бөліп, басқа жаңадан дайындалған қоректік ортаға отырғызылады, себебі қоректік ортаның күші әлсірейді, ұлпаның қоректенуі және ұлпаның ішкі бөліктерінің аэрациялануы төмендейді. Жаңа қоректік ортаның құрамы эксперименттің мақсатына байланысты болады, Егер каллустың бір қалыпты өсуін ұстап (сақтау) отыру керек болса, онда бастапқы қоректік орта құрамы өзгермейді.
Кейбір тәжірибелерде клеткалардың бір-біріне тигізетін әсерін ескере отырып, каллустарды өсірудің арнайы әдістері қолданылады. Қоректік ортада өсіру әдісінің бірі " бағушы" немесе " асыраушы қабат" , яғни бір каллус өсу үшін өсіру әдісінің бірі - екінші бір каллус ынталандырушы ретінде қолданылады. Бұндай жағдайда ұлпаны қоректік орта сіңген фильтр қағазына салады да, есімдіктің сол түрінің жаксы өсіп тұрған каллус массасына отырғызады. Кейде бір ыдыстың ішінде әр түрлі өсімдіктердің ұлпаларын өсіруге болады. Кейбір жағдайларда ұлпаларды бір-біріне немесе мүшелерді ұлпаларға телуді қолданады. Жақсы өсіп тұрған ұлпалар мен мүшелерден бөлініп шыққан кейбір заттар, тәжірибеге алынған каллустың өсуін жылдамдатады.
Организм құрылысын жасайтын элементтердің жаңадан түзілуіне байланысты организмнің дене көлемі мен массасының қайтымсыз үлкейуі өсу деп аталады. Клетканың, құрылымдық элементтері: органеллалар, мембраналар, макромолекулалар және метаболиттер,ұлпаны құрайтын-клеткалар,ал мүшелер-олардың құрамына кіретін әр түрлі ұлпалар типтерінен тұрады.Сонымен,өсу дегеніміз сандық жағынан тірі компоненттердің молекуладан организмге дейін көбейу процессі /молекулалар-клеткалар-ұлпалар-мүшелер-организмдер/.
Өсімдіктің өсуі клеткалардың, ұлпалардың және мүшелердің өсуінен тұрады. Солай бола тұрса да, қандай да болсын өсудің негізі клеткадан басталады. Клетканың өсуі бірінен соң бірі кезекпен келіп тұратын процесстерден тұрады: клетканың бөлінуі, протоплазманың өсуі, созылуы және дифференциялануы. Өсу процестері негізінен меристемада жүреді.
Меристемалар жылдам бөлінетін клеткалары бар түзуші ұлпа. Клетка бөлінуінің көбірек кездесетін жолы митоз, ол төрт стадиядан тұрады: профаза, метафаза, анафаза, телофаза. Клетка бөлінер алдында оның компоненттері екі еселенеді. Клетканың бөліну циклінің, анық ең қолайлы маркері ол ДНҚ - ның мөлшері. Әрбір екі митоз аралығында ДНҚ - ның екі еселенуі /репликациясы/ жүреді. Клетка циклінің түрлі фазалары мынадай символдармен белгіленеді, М, Оі , С2 және 8 фазасы.
Клетка циклінің М фазасы ядроның бөлінуі /митоз/ және цитоплазманың (цитокинез) бөлінуінен тұрады, осы фазадан кейін, жаңадан түзілген клеткалар жаңа клеткалық циклдің интерфазасына өтеді. Интерфаза О1 фазасынан басталады
Митоз кезінде төмендеген биосинтездік процесстер бұл кезде қарқынды түрде жаңадан басталады.Циклдің 3-фазасы ДНҚ-ы синтезделетін кезең, ядродағы ДНҚ-ы екі еселенгенде хромосомалар түгел репликацияланады /әрбір хромосома екі тең хроматидтермен тұрады/. Одан кейін клетка С фазасына өтеді,бұл кезде клетка бөлінуге дайындалады. М фазасы митоздан басталады да /атының шығуы осыған байланысты/ цитокинезбен аяқталады. М фазасының басталуы кезеңінде екі еселенген хромосомалар, енді тығызданып жарықтық микроскоптан көрінетіндей болады. Ядро қабығы бұзылады да хромосомалар ретті түрде орналасып жеке хроматидтерге жіктеледі. Клетканың полюстерінде орныққан хромосомалардың сыртынан ядролық кабық пайда болады, жана ядролардың кайта құрылуымен қатар цитоплазма бөлінеді,бір ядросы бар екі жаңа клетка түзіледі М фазасы цитокинез процессімен аяқталады да, келесі клеткалық циклдің интерфазасы қайта басталады.
Кейбір жас клеткалар митоздан кейін созылу фазасына ауысады.Созылу өзіне суды сіңіру аркылы жүреді,сонымен қатар бұл кезде белоктар нуклеин қышқылдары, көмірсулар, майлар және басқа заттар жаңадан түзіледі. Жаңадан бөлінген эмбриондық клетканың клетка қабығы және цитоплазма компоненттері түзіле бастайды. Клетканың созылуы аяқталғаннан кейін келесі кезеңге дифференциялану яғни мамандану жағдайға ауысады.
Сонымен,клетка бөлінгеннен кейін, әрбір жас клетканың алдында үш мүмкіндік болады. 1/Эмбриондық жағдайда қалып, клетка цикліне түсіп, митоз арқылы қайта бөліне алады. 2/Циклден тыс қалып, /О0/бөлінбей, тыныштық кезеңіне өтеді. З/Компетенцияға иеленіп жаңа бағыты біртіндеп айқындалып одан кейін ерекшелену /дифференциялану/ жолына түседі.
Комнетенция- клеткалардың, ұлпалардың, мүшелердің, организмнің индукторлық әсерді қабылдап алуға кабілеттігі және әсер еткен факторға жауап ретінде өзінің даму бағытын өзгертуі.
Өзгерістерді әр түрлі факторлар туғызуы мүмкін. Мысалы: гормондар,көрші клеткалардың немесе басқа ұлпалардың, метаболиттері, электрофизиологишшқ сигналдар және т.б.
Клетканың белгілі бір тұқым куалауды жүзеге асыруға әзір тұру қалпына тусуі-детерминация деп аталады. Қабілеттілігі бар клеткалық детерминациялануы оның дамуының белгілі бір жолын калыптастырады да,жетілудің басқа бағыттарда жүруін тежейді.Қабілетті клетканың детерминациялануы клетка бөлшегінен кейін бірден басталуы мүмкін яғни протоплазманың өсуі алдында. Белгілі бір ретаен детерминацияланған клетка өте бір тар мамандыққа ие болады яғни дифференцияланады (қандай да бір ұлпаның клеткаларына айналады).
Дифференциация - жас клеткалар арасындағы құрылысы мен кызметі жағынан болатын өзгерістерді,аналық клеткалары мен жаңа пайда болған клеткалар арасында айырмашылықтарды тудыратын күрделі процесс. Дифференциялану (дифференцировка) - клетканың маманданған жағдайы, басқа клеткалардан ерекшеленуі. Дифференциялану деген түсінік ол эмбриондық клетканың маманданған клеткаға айналуы дегенді білдіреді.
Меристемалық клеткалар атқаратын қызметі және құрылысы жағынан біркелкі, дифференциялану нәтижесінде әр жолдармен дами бастайды да,түрлі мүшелердің ұлпаларын түзеді. Клетканың қандай да бір даму жолына тусуі, синтезделуі гендер аркылы басқарылатын белгілар құрамымен анықталады.
ДНҚ-ы молекуласында шифрленген генетикалық информацияның іске асуы арнайы и-РНҚ түзілуі /транскрипция/ арқылы және одан кейін генетикалық кодына сәйкес белоктар синтезі /трансляция/ арқылы жүретіні белгілі. Бірақ, барлық генетикалық информация бір уақытта іске аспайды, клетканың өсу және дифференциялану процесстері кезінде қатаң тәртіппен кезектесіп көрініп отырады. Геномдағы гендердің бір тобы бір кезеңде активті болады, яғни иРНҚ-ның мағыналы өнімі бар, ал келесі бір тобы активтілігін көрсете алмайды, себебі, ондай гендер активтілігі репрессор әсерінен тежелген. Сондықтан, клеткалар арасындағы айырмашылықтар, яғни олардың дифференциаялануы гендердің әр түрлі дифференциалды/ активтенуіне байланысты.
Клетка құрамындағы белоктар фенотипті және клетканың қызметін анықтайды. Клетка қасиетін, құрылыстарын транскрипция процессі кезінде иРНҚ-ы анықтайтын ферменттер мен басқа да белоктардың әсеріне байланысты болады. Бұл жағдай 2 суретте анық көрсетілген.
Клетка ерекшеленіп маманданған клеткаға айналғанда, /дифференцияланғанда/ оның құрылысы және онда жүретін зат алмасу да өзгереді, ал гендер жиынтығы организмнің барлық даму кезеңінде өзгермей бір қалыпты болады. Клеткада жүретін өзгерістер тек гендер активтілігіне байланысты. Бүл жағдайдан клетка ерекшеленген маманданған клеткаға айналуын /дифферанциялануын/ түсіну үшін, негізгі мәселені анықтауымыз қажет; неге бір типті клеткаларда гендердің бір бөлігі активті, ал басқа типті клеткаларда басқа гендер ахтивті.
Гендер активтілігінің реттелуі мынадай алты деңгейде жүруі мүмкін:
1/ ДНҚ-ы синтезделуі /редукцикация/ деңгейінде: 2/ РНҚ-ы синтезделуі /транскрипция/ деңгейінде: 3/ продессинг деңгейінде:
4/ и РНҚ~ның ядродан цитоплазмаға тасымалдану деңгейінде:
5/ рибосомалардағы трансляция деңгейінде:
6/ цитоплазмада иРНҚ-ның кері кету деңгейінде.
Гендер активтілігінің ДНҚ синтезі деңгейінде реттелуі гендер репликациясының бір қалыпты жүруін бұзуы мүмкін, сондықтан әр түрлі клеткалардағы гендер жинағы да бірдей болмайды. Сонымен бірге гендер активтігі ДНҚ-ның күйі өзгеруіне де тәуелді. Гендер активтілігіне әсер ететін, ДНҚ-ның тұрақты өзгерістерінің үш типі белгілі. Біріншіден, хромосомадағы гендердің өзара орналасу ретінің өзгеруі, олардың атқаратын қызметіне әсер етуі мүмкін. Транслокация немесе инверсия кезінде ген орын алмастырып ДНҚ-ның басқа бөлігі арасына орналасады. Бұл жағдайда ген активтілігінің төмендеуіне немесе өте жоғары деңгейде көрінуіне әкеліп соғады. Гендердің орын алмастыру құбылысын, яғни жылжымалы генетикалық элементтерді алғаш рет Барбара Мак-Юшнток 1947 жылы ашқан. Кейінгі кезде белгілі болғандай транспозондар немесе жылжымалы генетикалық элементтер бір хромосомадан басқа хромосомаға орын ауыстыра алады, осыған байланысты гендердің жоғары дәрежеде мутациалануын жүргізеді немесе бұрын активтілігі жойылған гендердің активтілігін арттыра алады. Екіншіден, геномдағы қандай да бір геннің санының көбеюі де /амплификация/ геннің іске асу дәрежесіне /экспрессиясына/ әсер етеді. Өте жиі амплификацияға ұшырайтын рРНҚ- ның гендері. Геномдағы бір гендердің көбеюінің /амплификациясының/ клетканың ерекшелініп маманданған клеткаға айналуындағы /дифференциялануындағы/ рөлінің қаншалықты маңызы бар екендігі өлі белгісіз. Үшіншіден, ген кұрылысының сапалық жағынан (мутация, делеция) өзгеруі мүмкін.
Прокариоттардағы транскрипция деңгейіндегі белок синтезінің реттелуін Жакоб және Моно жасаған оперон (бір репрессордың бақылауындағы гендер тобы) моделі жеткілікті дәрежеде түсіндіріп береді. Эукариоттар гендерінің активтілігінің реттелуін зерттеуше қиын, себебі, геномы күрделі және көлемі үлкен. Эукариот гендерінің дифференциялды активтілігі транскрипция денгейінде геномның белгілі бөліктерінің ашылу немесе көріну жолымен реттелуі мүмкін, яғни матрицадан информация алудың мүмкіншілігі өзгеруі арқылы.
Жоғарғы сатыдағы организмдер гендерінің реттелуі туралы гипотезаны Бритген мен Дэвидсон жасаған. Олардың болжауы бойынша гендердің экспрессиясының реттелуі ядрода алғашқы иРНҚ-ның кайсысы сақталып қалып, процессингке ұшырап, кейін цитоплазмаға өтетіні анықталған кезінде жүреді.
Цитоплазмаға өткен и РНК-ның трансляциясы бірден жүрмейді. Эукариоттардың эволюция барысында қол жеткізген жетістіктерінің бірі информосомалар. Информосомлар белок синтезінің реттелуін қамтамасыз етеді. Гендер экспрессиясының реттелуінің тағы бір кезеңі, ол трансляция процессі. Бірақ трансляция деңгейінде реттелу механизмдері туралы деректер жоқ деуге болады.
Сонымен, өсімдіктерде жүретін дифференциялану процессі өсімдіктердің қандай да бір бөліктерінің немесе тіршілік циклінің кез келген стадиясында өтетін гендер экспрессиясының айырмашылықтары арқылы анықталады.
Морфогенез дегеніміз пішін /форма/ құру процессі, яғни өсімдік мүшелерінің бастапқы элементтерінің құрылуы, өсуі және өнуі /органогенез/, ұлпалардың /гистогенез/ жөне өсімдік клеткаларының жетілуі /цитогенез/ немесе клетканың дифференциялануы. Сөйтіп, морфогенез көп кезеңдерден тұратын күрделі процесс.
Организмнің дамуы - өсу, дифференциалануы, морфогенез процесстерінің бір - бірімен тығыз байланыста жүруінің нәтижесінде болады. Бүтін организмде бүл процесстерді бір - бірінен ажыратып қарау өте қиын. Сонымен қатар, бұл процесстер бір - біріне мүлде ұқсамайды, әрқайсысының өзіндік ерекшеліктері бар. Сол себепті, морфогенез процессін түсіну үшін оны өсу мен дифференциациядан бөлек алып қарау қажет. Сонымен қатар, морфогенез өсу мен дифференциациясының арқасында жүретінін естен шығармау қажет. Атап айтқанда сандық өзгеріс және сапалық жағынан қайта құрылудың нәтижесінде кеңістікте құрылысы және формасы бар құрылым пайда болады, яғни морфогенез процессі жүреді.
К. Уоддингонның анықтамасы бойынша, "Морфогенез дегеніміз - гомегендік, яғни жеке бөліктерден тұрмайтын массасының белгілі бір құрылымға айналуы". Тұтас организмде әр түрлі бөліктердің бір - біріне тигізетін әсерін камтамасыз ететін, көптеген корреляциялар, байланыстар болғандықтан, бұндай жүйеде морфогенездің көрінуін зерттеу өте қиын шаруа .
Морфогенезді зерттеудің өте қолайлы моделі, ол ұлпаларды, клеткаларды және протопласттарды жасанды қоректік ортада өсіру. Моделді қолданып жүргізген зерттеулердің артықшылығын, ол өсірілетін клеткаларға бір бағытта әсер ету арқылы, зерттеуші түрлі факторлардың әсерін бақылай алады.
Дәріс № 9,10
Іn vіtro жағдайында өсетін өсімдік клеткаларының биологиясы, дифференциялану
Жоспар : 1 Қоректік ортада өсірілген клеткалар биологиясы.
2. Дедиффереңциялану және каллустың түзілуі
3. Жасанды ортада өсірілген клеткалардың әр түрлілігі.
4. Клеткалардың жасанды ортада өсуі.
1. Көп уақыт бұрын бөлінуін тоқтатқан ұлпалардың дифференцияланған клеткалары қайтадан митоз жолымен бөліне бастағанын көрсетеді. Маманданған, бөлінбейтін клеткалардың қайтадан бөлініп, жаңа клеткалар мен ұлпалар түзу жағдайына ауысуы - дедифференциация деп аталады. Бұл процесстің негізі, ол дифференциациядағы сияқты гендердің активтігінің өзгеруіне байланысты. Клетка атқаратын қызметінің өзгеруі, гендер активтілігнің өзгеруіне байланысты. Осы уақытқа дейін іске косылмаған геңдердің активтенуі және бұрын жүмыс істеп тұрған кейбір гендердің тежелуі себебінен, клеткадағы құрылымлдық белоктар мен ферменттердің құрамы өзгереді. Қоректік ортада міндетті түрде фитогормондар / соның ішінде ауксиндер/ болған жағдайда клеткалар дедифференцияланып каллустың клеткаларға айналады. Маманданған клетканың дедифференциялануының бірінші этапы ол клетканың бөліну қабілетінің қалпына келуі. Дедифференциациялану және каллус түзілу механизімін зерттеп білетін модель ретінде жасанды ортада өсірілген темекі сабағының өзектік паренхимасын пайдаланған өте қолайлы. Себебі, өзектік паренхимада бөлінуді козғау мен каллус түзуші үшін ортада ауксин мен цитокининнің қатысуына қатан тәуелділік бар.
Дедифференциялану калай жүреді? Р.Г.Бутенконың келтіруі бойынша, қоректік ортада өсірілген темекінің өзектік паренхима клеткаларының дедиф- ференциялануы, қоректік қор заттарды пайдаланудың және клетканың арнайы маманданған органеллаларының бұзылуынан басталады. Маманданған клетканың өз қасиетін жоя бастағаннан /дедифференциялану/ кейін 6-І2 сағ. өткен соң олар клетканың нәзік құрылысында болатын өзгерістерді бақылаған. Клетка қабығы жұмсарып, ісінген, рибосомалар саны, эндоплазмалық тор және Гольджи аппараты элементтерінің саны өскен, ядрошықтың көлемі және саны көбейген. Клеткаларда тРНҚ және рРНҚ синтезі артып, ДНҚ синтезі басталған, жаңадан түзілген белоктардың мөлшері көбейген. Бастапқы ұлпаларға тол емес, арнайы белоктардың-антигендердің пайда болуы, әсіресе тұтас темекі өсімдігінің меристемалық белогының біріне сәйкес белоктың синтезделуі анықталған. Осы өзгерістердің барлығы, одан кейінгі клетканың бөлінулері тек ортада ауксиндер мен цитокининдердің катысуымен ғана жүреді. Клетка бөлінулері 48-72 сағат өткеннен соң басталатыны белгілі болды. Дмитрева Н.Н мен Линский А.Х. темекінің өзектік паренхима клеткасың циклді бөлінуіне гормондар әсерінің мерзімдік жүйелілігін анықтады, ол үшін ауксин мен кинетин әсері жеке-жеке зерттелді. Қоректік ортаға бір ғана ауксинге қосылған жағдайда, РНҚ және ДНҚ синтезделуі күшейгенімен, клеткалардың бөлінуі байкалмады, төрт күндік латентті / жасырын / фазадан кейін клетка созылып өсуге айналды. Қоректік ортада тек бір ғана кинетин қосылған жағдайда нуклеин қышқылдары синтезінде ешқандай өзгерістер байкалмаған. Клетканың бөлінуіне кажет барлық процесстер жүру үшін, ауксиннен кейін кинетиннің әсері болу керек немесе екеуінің қосылған әсері қажет. Зерттеулерден кейін мынадай қорытынды жасауға болады, клетканың бөліну процессін тек кинетин бастап жібереді. Р.Г.Бутенконың үйғаруы бойынша клетканың бөлінуі төменгі жағдайларды қамтиды: клеткалардың көбею стимуляторы /ауксин/ әсерінен клетканың сырткы мембранасы плазмалемманың өткізгіштігі артады да, оның қасиеттері өзгереді. Осыдан соң бұрыннан бар болған немесе жаңадан синтезделген матрицалық РНҚ-да цитоплазмалық белоктардың түзілу барысы активтенеді. Түзілген белоктар цитоплазмадан ядроға өтеді, арнайы гистондық белоктармен ерекше байланысқа түседі де , транспорттық және рибосомдық РНҚ-лары ситезделуіне жауапты локустарды іске қосады. Клеткадағы РНҚ-ның мөлшері белгілі бір сатылы деңгейге жеткенде ДНҚ-ның репликациясы басталады. Ортада кинетин болмаған жағдайда клетканың митозға өтуі болмайды. Шамасы, кинетин клетканың митозға өтіп бөлінуін аяқгау үшін қажет, арнайы РНҚ-лары мен белоктардың сиитезделуін белгілейтін болу керек. Ондай бөлінуге ықпал жасайтын премитоздық кезеңде синтезделген белоктың және оған сәйкес информациялық РНҚ-ы бар екендігі жануарлар клеткаларында дәлелденген.
Сонымен кез келген клетканың өзгеріп, каллустық клеткаларға айналуының алдында, көптеген күрделі биохимиялық және структуралық кайта құрылулардан өтеді. Дегенмен, өсімдік клеткаларының маманданған касиетін жойып, бөліне бастау /дедиференциялану/ процессінің терең механизмдері жете зерттелген жоқ.
Жаңа өсімдік материалынан бірінші рет каллус алу өте қиын, себебі әр түрлі өсімдіктердің ұлпалары, тіпті бір өсімдіктің әр бөлігінен алынған ұлпаларының дедиференциалануы және каллусогенезі жүру үшін, әр түрлі қоректік орталарды талап етеді, әсіресе ондай талап гормондардың мөлшері мен ара қатынасына қойылады. Кейбір өсімдіктердің, ұлпаларын кайталап отырғызып ұзак, мерзімде өсіргенде, ондай ұлпалар әжептәуір мөлшерде ауксин мен қатар цитокининді синтездеу қабілетіне ие болады, яғни бұл гормондарға байланысты автотрофтарға айналады. Бұндай каллус клеткалары "қалыптасқан",-деп аталады. Гормондарға тәуелсіз штаммдардың пайда болу себебі өлі белгісіз. Олардың шығуы мутациялардың әсерінен, немесе жаңа гендердің іске қосылуынан болуы мүмкін. Ісік клеткаларын жасанды ортада өсіргенде, оларда гормондарға тәуелсіз.
Клеткаларды қоректік ортада өсіргенде каллустық клеткаларға айналады, бірақ ондай жағдайда барлық клеткалар біркелкі болмайды. Жасанды ортада өскен клеткалардың түрлілігі/гетерогенді/ морфология, физиология және генетика түрғысынан болуы мүмкін.
Каллус клеткаларының бастапқы ұлпа клеткаларынан және бір-бірінен өлшемі және формасы жағынан айырмашылыктары бар, олардың ядросының саны мен формасы тұрақты емес. Қоректік ортада өскен клеткалардың полиморфизмі мынадай себептерден пайда болуы мүмкін. Түрдің және өсу мерзіміндегі ерекшеліктері, алғашқы плоиттылық деңгейі, қоректік орта құрамы және өсіру жағдайларының тигізетін әсері, клетклар арасында корреляциялық байланыстардың жойылуы. Қартайған каллустың клеткаларының мөлшері өте үлкен шамаға дейін жетеді/500-1000мкм/, жас клеткалардың мөлшері айтарлықтай кіші/15-30мкм/. Ортада кинетин болмаған жағдайда көбінесе клеткалардың мөлшері үлкейеді. Клетканың пішіні мен мөлшерінің және ядроларының өзгеруі, ол плоидтылығының көбеюіне байланысты. Клеткаларды жаңа ортаға қайталап отырғызып өсіру /пассаж/ саны көбейген санын плоидтылығы артатыны дәледденген.
Физиологиялық әр түрлілік деген ол популяциядағы клекалардың әртүрлі физиялогиялық жағдайында болуы, яғни олар бөлінеді, өседі, картайып, өледі. Сейтіп ортада өскен клеткалар асинхронды келеді /бөлінуі бір уақытқа сәйкес келмеуі/. Жоғары сатыдағы өсімдіктер популяциясындағы клеткаларды клетка циклі фазаларын бір кезде өтуге мәжбүр ету, яғни синхрондау мүмкіндігі жок деуге болады. Себебі, бір мезгілде бөлінуге қабілетті клеткалардың бөлігі 2-4 құрайды. Қолайсыз жағдайлар төменгі температура, қоректік ортада негізгі компоненттердің жетіспеуі клеткалар бөлінуін тежейді және бөлінуге дайын клеткалар санының көбеюіне себепші бола алады. Клетканы бөлуге дайындық кездерінің бірін тежейтін химиялық заттарды қолданған тиімді. Клекалардың бөлінуінің синхрондалуының жоғары деңгейі 10-30%-ке жетуі мүмкін, бұл өте жақсы жағдай, бірақ келесі бөлінулер кезінде популяция өзінің бір мезгілде бөліну қасиетін тез жояды.
Көңіл аударатын жай, ол сұйық қоректік ортада /суспензияда/ өскен клеткалардың физиологиялық өзгергіштіктері, агарланған қатты ортада өскен каллустың ұлпаларымен салыстырғанда төмен. Ол клеткалардың қоректену мен аэрациялау жағдайларының бір келкілігіне және клеткалар бөліп шығаратын зиянды зат алмасу өнімдерінен, сұйық араласып тұрған ортада оңай құтылуына байланысты.
Жасанды қоректік ортада өскен клеткаларға цитогенетикалық тұрақсыздық та тән. Бір ыдыста өсірілген, бір ұлпа клеткаларыньң хромасомалар бір-бірінен айтарлықтай айырмашылығы /диплоидтар, полиплоидтар, анеуплоидтар/ болуы мүмкін. Генетикалық өзгергіштіктің себебі әр алуан. 1. Өсімдіктен бастапқы эксплантты бөлген кезде үйлесімді /коррелятивтік/ байланыстардың бұзылуынан, 2. қоректік орта компоненттерінің әсерінен, 3. коректік ортаға бөлінген метаболизм өнімдерінің әсерінен, 4. сегізгі материалдың гетерогенділігі және бөлгіш клеткалар типтерінің сұрыпталуы.
Жасанды жағдайда өсірілген клеткалардың цитогенетикалык гетерогенділік механизмі жалпы түрде анықталған. Оларға жататындар: 1/ клеткалық сұрыптау процессі; 2/ клеткалардың бөлінуінде эндомитоз бен эндоредупликация құбылыстары; 3/ митоз кезінде клеткада жүретін жағымсыз өзгерістер. Жоғарыда келтірілген үш процесстің әр қайсысы бір-бірінен тәуелсіз жүре алады. Клеткалық сұрыптау, ол популяциядағы клеткалардың белгілі типінің көбейіп басым болуы. Ондай басым болу клеткалардың бір бөлігінің айрықша бөлінуінен немесе басқа клеткалардың тез жойылуынан болуы мүмкін. Эндомитоз және эндоредупликация /басқа атаулары эндоциклдер/ тек қана клетканың жаңадан түзілуіне, яғни пролиферацияға байланысты болады. Бұл екеуі де митоздың формалары, олардың ерекшелігі ол карио және цитокинездің өтпейтіндігі, және ядродағы ДНҚ-ның мөлшері геометриялық прогрессия бойынша көбейетіндігі. Қалыпты митоздың бұзылуы мен хромосомалардың дұрыс бөлінбеуі, полиплоидтық және анеуплоидтық клеткалардың түзілуіне әкеліп соғады.
Ең негізгі себептердің бірі, шамасы, ол бүтін организмнен бақылау болмауына байланысты болу керек, яғни корреляциялық байланыстардың бұзылуы себебінен клеткалар өзгереді. Тіршілікке қабілетті полиплоидтық клеткаларды сұрыптау арқылы, клеткалар популяциясы жаңа жағдайға бейімделеді. Жасанды қоректік ортада өсіру жағдайы өзгергенде, сұрыптаудың бағыты да өзгереді, бұл ескеретін жағдай. Мысалы, ортадағы 2,4-Д және юшетиннің жоғары концентрациялары клеткалардың полиплоидтану мумкіндігін арттыратыны анықталған.
Жасанды ортада өсіру жағдайынын цитогенетикалық көп түрліліктің /гетерогенділігін/ пайда болуына әсер ететіндігін гаплопаппус өсімдігі ұлпасымен жасалған тәжірибеден анық көруге болады. Осы өсімдіктің меристемалық клеткалары Р.Г.Бутенконың лабораториясында екі жыл бойы үзбей әрбір ай сайын жаңа ортаға отырғызу арқылы өсірілген. Сондықтан да, негізгі өсіруге алынған диплоидты клеткалардың 95%-і жоғары плоидтылықка айналған. Осы ұлпамен жұмыс жасаған швед зерттеушісі Эриксон, әрбір екі күн сайын жаңа қоректік ортаға отырғызып отырған. Бұндай жағдайда штамм бір қалыпты диплоидты күйін сақтап калған. Дегенмен, өсімдіктерді өсіру тәсілдері клеткалар популяциясының генетикалык тұрақтылығының толық кепілі бола алмайды, себебі негізгі материаддың екінші генетикалық әртүрлілігі, болуы әбден мүмкін. Көптеген өсімдіктердің дифференциалданған ұлпалары клеткаларының диплоидтылығы әр түрлі. Маманданған клеткалар, мысалы, жарық сәулесін сіңіретін жасыл жапырак паренхимасы, тамырдың етженді қорлық ұлпалары, түйнектер клеткалары көбінесе полиплоидтык, Тек белсенді түрде бөлініп жатқан клеткалар әр уақытта диплоидтылығын сақтап қалады.
Егер негізгі материал ретінде диплоидтық меристемалық клеткалар алынған болса және оларды үзілмейтін ағын коректік ортада өсірсе, сонда ғана олардың генетикалық тұрақтылығын сақтауға кепілдік бар. Экономикалық жағынан тиімді, қажеттігі зор, түрлі заттар алу мақсатымен ұлпалардың өте үлкен массасын өсіргенде, клонның генетикалық тұрақтылығын сақтаудың маңызы зор. Клеткаларды жасанды түрде өсіру барысында, өсімдіктің негізгі түрімен генетикалық пара-парлықты сақтау, бірегей, генетикалық жағынан бағалы өсімдіктер формаларын көбейткенде қажет екендігі белгілі. Сонымен қатар, клеткаларды модель ретінде физиологиялық және биохимиялық процестерді зерттеу үшін қолданғанда да олардың генетикалық тұрақтылығының маңызы зор. Демек, клеткаларды қоректік ортаға бірнеше қайталап отырғызып, өсіріп тіршілігін сақтағанда, оларда морфологиялық, физиологиялық, цитогенетикалық әртүрлілік пайда болады. Кайталап отырғызбай клеткалар штаммдарын өзгеріссіз сақтау жолының бірі мұздатып алып өте төмен температурада /-196°С /, яғни криогенді әдіспен сақтау.
Қалай дегенмен, жасанды ортада өсірілген ұлпалардың -генетикалық гетерогендігінің пайдалы жақтары да бар, ол клеткалардың адаптациялық мүмкіншілігін арттыратындығы. Клеткаларды жасанды қоректік ортада өсіргенде, полиплоидтық клеткалардың өздігінен сұрыпталуы /автосұрыптау/, яғни полиплоидтық клеткалардың спонтанды /табиғи/ сұрыпталуы жүреді. Ондай сұрыпталған клеткалар тіршіліктің жаңа жағдайларына тезірек бейімделе алады. Бұдан басқа, табиғи немесе қандай да бір факторлар әсері арқылы индукцияланған, өсімдіктердің түрліше варианттары бар формаларының пайда болуын, казіргі уакытта қолданып жүрген ауылшаруашылық дақылдарының сапасын жақсарту үшін колдануға болатыны. Қоректік ортада өсіріліп алынған клеткалар линияларының немесе регенерант өсімдіктердің көп түрлілігі сомаклондық өзгергіштік нәтижесінде пайда болады, оларды сомаклондык варианттар деп атайды. Сомаклондық өзгергіштік -өсімдік клеткаларының ядролық және органиодтық геномдарының тұрақты еместігінен болатын фенотиптік өзгергіштік.
Агарланған және де сұйық жасанды қоректік орталарда өсірілетін өсімдік клеткаларының негізгі типі, ол көбеюге қабілеті бар каллус ұлпалары. Кез келген негізгі ұлпалардан пайда болған каллус ұлпаларын өзіне лайықты қоректік ортада ұзақ уақыт бойы, тіпті шексіз өсіруге болады. Үзбей өсіру үшін әрбір 3-4 апта өткеннен кейін каллустарды жаңа қоректік ортаға отырғызып /пассаж жасап/ бағады. Дүниежүзінің көптеген лабораторияларында ондай каллустар өсірілуде. Мысалы, 1938 жылы сәбіздің тамыр жемістерінен Р.Готре алған каллус күні бүгінге дейін өсірілуде. Ескертетін жағдай, өсімдіктердің жеке клеткалары олардың кішкене топтарын /агрегаттарын/ өсіргенде, өсуге қажет қоректік заттарға қоятын талабы қатты ортадағы клеткаларымен салыстырғанда жоғары болады, өйткені агардағы клеткалар өзі тәріздес клеткалармен қоршалған. Жеке клеткалар араластырылып тұрған сұйық қоректік ортада бөлінуі үшін қажет, қандай да бір заттарын жоғалтады, сондықтан сұйық қоректік ортада өсірілетін клеткалар үшін орталардың құрамы өте құнарлы болу керек. Агарланған ортаға отырғызылған жеке клеткалар бөлінуге өту үшін, көмек ретінде "бағушы" ұлпаларды немесе "асыраушы қабаты" болуын қажет етеді және ортаға отырғызылатын клеткалар тығыздығы белгілі бір мөлшерден төмен болмау керек. Сұйық қоректік ортада өсетін бірен-саран клеткалар басқа жақсы өсіп жатқан клеткаларының ортасын қосқан соң ғана бөлініс бастайды.
Бүтін организмдер сияқты жасанды ортада клеткалардың өсуі 3 -тәрізді қисық сызықпен бейнеленеді және ол мынандай фазаларды қамтиды:
I/ латенттік немесе лаг-фаза, бұл фаза кезінде клеткалардың көрінетіндей өсуі болмайды, бірақ клетка белсенді түрде суды және коректік заттарды сіңіреді де бөлінуге дайындалады. 2/ үдеу фазасы, бұл кезде клеткалар бөлініп созылып өсе бастайды; 3/ логарифмдік немесе экспоненциялдық фазасы, түзу сызыкпен бейнеленетін өте қысқа фаза, бұнда өсу жылдамдығы уакыт өткен сайын екі еселеніп үдейді; 4/ Клетка өсуінің төмендеу фазасы, бұл фазада клетка өсуінің салыстырмалы жылдамдығы бәсеңдейді; 5/ стационарлық немесе бір сарынды фазасы, бұл кезде клетканың өсуі жылдам болмаса да тұрақты жүреді; 6/ клеткалардың біртіндеп жойылып құру фазасы.
Клеткалардың бір фазадан басқа фазаға ауысуы ішкі және сыртқы факторлардың катысуымен бақыланады. Ішкі факторларға жататындар: пролиферациялық қоры; клетканың созылу ұзақтығы; циклға енген клетканың жалпы күйі; жеке клеткалардың бөліну саны. Пролиферациялық қоры деп бөлініп жатқан клеткалар санының жалпы клеткалар санына қатынасын атайды. Бұл ара қатынасты процентпен белгілесе, онда ол митоздық индекс деп аталады.
Сыртқы факторларға жататындар : қоректік ортаның құрамы, рН-тың деңгейі, оттегінің мөлшері, температура, отырғызылған клеткалар мен ұлпалардың тығыздығы, т.б. факторлар.
Клеткалар мен ұлпаларды жасанды ортада өсіргеңде барлық клеткалар бірдей бөлінбейді, сондықтан пролиферациялық қоры төмендейді. Оны мынадай себептермен түсіндіруге болады:
1) клеткалардың қайтымсыз дифференциялануы;
2) клеткалардың С кезеңіне өтуі (циклден тыс немесе тыныштық кезеңі);
3) клеткалардың кұруы. Клеткалардың бөлінуі арқылы өсуі мен бір бөлігінің өліп жойылуы теңелгенде, жасанды ортада өсрілген клеткалар стационарлық (өсуі бір қалыпты өзгермейтін) күйде болады.
Өсу процессін түрлі факторлардың қатысуымен реттеуге болады және мынадай көрсеткіштер арқылы бақылауға болады: өлшем, көлем, масса, клеткалар саны, белоктың және ДНҚ-ның мөлшері. Тәжірибенің мақсатына байланысты, келтірілген көрсеткіштердін, біреуі қолданылады. Клеткалардың өсуін зерттеп білу үшін, белок пен ДНҚ мелшерін белгілі бір әдістермен анықтаған ыңғайлы және оны клеткалар санын анықтағанда да қолдануға болады.
Бақылау сұрақтары
1 Қоректік ортада өсірілген клеткалар биологиясы.
2. Дедиффереңциялану және каллустың түзілуі
3. Жасанды ортада өсірілген клеткалардың әр түрлілігі.
4. Клеткалардың жасанды ортада өсуі.
Дәріс №11,12
Гендердің дифференциялды активтелінуі және белок синтезін реттеу деңгейлері.
Жоспар: 1. Гендердің экспрессиялануының (активтенуінің) реттелу механизмдері.
2. Гендердің экспрессиялануының реттелу механизмдері.
3. Транскрипциялану немесе РНҚ синтезінің механизмдері
4. Белок биосинтезі.
Тұқым қуалаушылықтың материалы - ДНҚ молекуласы, ізінен ядрода шоғырланған, ал белок биосинтезі цитоплазмада-полисомада жүреді. Сонда, ДНҚ молекуласының бойына "жазылған" ақпарат қалайша цитоплазмаға жеткізіледі? ДНҚ молекула-сындағы генетикалық ақпараттың цитоплазмада белок-фермент синтезі күйінде жүзеге асуы тіршіліктің түпкілікті қасиеттерінің бірі болып саналады.
Цитоплазмада белгілі бір белоктың синтезделу акгивтігі гендердің активтігіне (экспрессиялануына) байланысты екені айдан айқын. Жасушада белок биосинтезі екі саты арқылы жүзеге асады-транокрипция және трансляция. Транскрипция дегеніміз геннің экспессиялануы (активтенуі) нәтижесінде а-РНҚ молекуласының синтезделуі, яғни ДНҚ молекуласындағы ақпараттың а-РНҚ молекуласына көшіріліп жазылуы (транскрипция-көшіріліп жазылу деген ұғымды береді), ал трансляция а-РНҚ молекуласының негізінде цитоплазмада-полисомада, полипептид молекуласының синтезделуі болып табылады.
Транскрипция өте күрделі процесс. ДНҚ бойына орналасқан гендердің бәрі бірдей, бір мезгілде транскрипцияланбайды. Ол, біріншіден жасуша тіршілігінің активтігіне және даму кезеңіне, екіншіден, гендердің экспрессиялануының реттелу механизмдеріне байланысты болады. Сондықтан да, бір мезгілде әртүрлі жасушаларда түрліше гендер транскрипцияланады және ағзаның дамуының әртүрлі кезендерінде бір жасушаның түрліше гендері экспрессияланады.
Әдетте біз, бір ген-бір белок (фермент) деген ұғым бойынша әрбір ген өз алдына жеке транскрипцияланады деп ойлаймыз. Ал, шын мәнінде бір белгіні дамытатын бірнеше гендер ДНҚ бойына қатар орналасып, бірге транскрипцияланады. Ондай гендерді кластерлі гендер деп атайды. Кластерлі гендердің бәрі бірдей транскрипцияланып ортақ полицистронды а-РНҚ түзіледі. Осының негізінде бір белгінің дамуына кажет барлық ферменттер бір мезгілде синтезделінеді. Кластерлі гендердің экспрессиялануын ерекше реттеуші гендер реттеп отырады.
Гендердің экспрессиялануының реттелу механизмдерін зерттеу үшін прокариотгар өте қолайлы объект болып саналады, себебі олардың геномдары небәрі бірнеше гендерден құралған және олар өте тез көбее алады. Сонымен қатар, гендердің экспрессиялануының реттелу механизмдері прокариоттарда да, эукриоттарда да ұқсас жүретіндігі аныкталды.
Гендердің экспрессиялануының реттелу механизмін зерттеуде ішек бактериясы - Езсһегісһіа соіі өте қолайлы объект болды. Е.соіі тіршілігі үшін қалыпты энергия көзі болып глюкоза саналады. Егер де тіршілік ортасында глюкоза болмаса ол лактозаны пайдалануға көшеді. Осы кезде жасушада лактозаны ыдырататын (3-галак-тозидаза ферменті синтезделуі қажет. р -галактозидаза ферменті дисахарид лактозаны галактозаға және глюкозаға ыдыратады.
Е. соіі жасушасында р-галактозидаза ферментінің синтезделуі қоректік ортада лактоза болған жағдайда индукцияланады, ал оның мөлшері азайса не мүлдем болмаса бұл ферментгің синтезделу карқыны да азаяды не тоқталады. р-галактозидаза ферменті синтезделу үшін Е. соіі ДНҚ-сындағы Ьас-2 деген ген транскрипцияланып оның а-РНҚ-сы түзілуі қажет. Р-галактозидаза ферментінің синтезделу қарқыны иңдукцияланғаннан кейін 1000 есеге дейін көбейеді және ол қоректік ортада индуктор-лактоза болса бір деңгейде ұзақ уақыт сакталынады. р-галактозидаза ферментінің лаісгозадан басқа иңдукторы ретінде оның ыдырауында пайда болатын аралық зат-аллолактоза да саналады. Ортада индуктордың (лактоза не аллолак-тоза) азаюы не жойылуы р-галактозидаза а-РНҚ-сының нуклеоидтерге ыдырап жойылуына алып келеді. А-РНҚ-ның тіршілік ұзақтығы бірнеше минутқа ғана тең, сондықтан да оның бір деңгейде синтезделіп тұруы үшін және жасушада үнемі р-галактозида ферменті синтезделуі үшін ол үнемі иңдукцияланып тұруы қажет, Р и ортада лактоза не аллалактоза болуы қажет. Сонда ғана синтезделуі теңесіп жасушада р-галакто-ғ;Гь2ерМент^ ^ір қалыпты синтезделінеді.
гендерінің экспрессиялануының реттелуін зерттеуде 1 және Ж.Моно еңбектерінің маңызы өте зор. Олар өздерінің тәжірибелерінің нәтижесінде 1961 ж. гендердің экспрессиялануы-ның реттелуінің оперондық теориясын ұсынды.
Е. соіі жасушасында лактозаны толық ыдырату үшін бір-бірімен тығыз байланысқан екі геннің өнімі қажет. Олар-р-галактозидаза ферментін анықтайтын Ьас-2+ гені және пермеаза ферментін анықтайтын-Ьас-У+ гені. р-галактозидаза ферментінің кызметі белгілі, ал пермеаза ферменті Е. соіі жасушасына лактозаны актив тасымалдау қызметін аткарады. Осы екі генмен катар орналасқан үшінші ген де болады ол Ьас-А+ гені. Ьас-А+ гені тиогалактозид трансацетилаза ферментін анықтайды. Ол жоғарыда келтірілген 2 генмен Ьас-2", Ьас-У+, тығыз байланысып кластерлік гендер кешенін кұрайды, бірге реттелінеді, бірге транскрипцияланады, бірақ лактозанын, ыдырауына қатынаспайды. Бұл үш ген мынадай тәртіппен орналасады 2-У-А. Осы үш геннің бірге индукциялануының нәтижесінде бір полицистронды а-РНҚ синтезделінеді.
Ф. Жакоб және Ж. Моно өздерінің тәжірибелері нәтижесінде мынадай болжам жасады: 2+,У+,А+ кластерлі гендердің транскрипциялануы оператор (О+) деп аталатын геннің бақылауында болады, ал оператордың қызметі, өз кезінде, ретфессор деп аталатын (І+) ген арқылы ретгелінеді. Репрессор екі түрлі қызмет аткдрады: 1)егер ортада индуктор (лактоза, аллалактоза) болмаса оның өнімі -репрессор молекулалары, операторды (О+) "тығыңцдап" әрі карай 2+, У+, А+ гендеріне ақпаратты өткізбей, олардың транскрипциялануын болдырмайды; 2)ал егер ортада индутсгор (лакгоза, аллалактоза) болатын боса, онда индуктор молекулалары (лактоза, не аллалакто-за) репрессор молекулаларына жабысып, қосылыл оларды активсіздендірді, сондықтан активсізденген репрессор молекуклалары операторды (О+) "тығындап" жаба алмай, акпарат әрі қарай 2+,У*,А+ гендеріне өтіп олардың транскрипциялануына "рұқсат" беріледі. Осы гендердін транскрипциялануы промотор учаскесінен басталады.
Ф.Жакоб, Ж.Моно- 2+,У+,А+ гендерін және оператор, промотор, репрессор учаскелерін оперон деп атаған.
Сонымен, гендердің экспрессиялануының реттелуіне 3 түрлі реттеуші элементтер катынасады.
1) реттеуші белоктар-транскрипцияның инициациялануында не тежелуінде РНҚ полимераза ферментінің активтігіне әсер ететін белоктар (репрессор);
2) эффекторлар-үсақ, белок емес заттар, олар ретгеушІ белок молекулаларымен қосылып олардың активтігіне әсер етеді (индук-торлар-лактоза, аллалактоза);
3) реттеуші нуклеотидтізбектері-ДНҚмолекуласында болатын кейбір реттеуші нуклеотид тізбектері (промотор, терминатор). Ретгеуші белоктардың осы учаскелерге тигізетін әсеріиің нәтижесінде а-РНҚ синтезінің активтігі реггелінеді.
Ф.Жакоб және Ж.Моно өздерінің оперондық гипотезасын тео-рия күйінде болжамдаған болатын. Ал қазіргі кезде оперон теория-сының шындығы көптеген тәжірибелер арқылы дәлелденген. Мы-салы, таза күйінде репрессорды бөліп алып зерттеп оның белок екендігі дәлелденді; репрессор молекуласындағы аминқышқылда-рының реті анықталып оның І+ геніндегі нуклеотидтер тізбегінің кодына колинеарлы болатындығы аныкталды. Сонымен қатар, реттеуші белоктардың ДНҚ молекуласының реттеуші аймақтарымен нақтылы байланысатындығы анықталды. Мысалы, 3,2 хЮ6 нуклеотид жүбынан түратын Е. соіі ДНҚ-сының ішінен Ьас-реп~ рессор таңцап-таңдап тек кдна белгілі бір нуклеотидтер тізбегімен (оператор) байланысады, ал оның үзындығы небәрі 24 нуклеотид ^Үбына тең.
Прокариотгар негізінде зерттелініп анықгалған гендердің эксп-рессиялануының ретгелуінің оперондық механизмі эукариоттарға да тән. Тек, эукариотгардың айырмашылығы, көп жасушалы ағзалар-да әртүрлі жасушалар бірдей және түрліше белоктарды синтездейді. Яғни эукариоттардың гендері екі топқа бөлінеді: 1)олардың бірі жасушаның түпкілікті, әмбебапты тіршілік қызметін қамтамасыз ететін, яғни барлықжасушалар типінде кездесетін, геңдер. Оларды "тұрмыстық" гендер деп атайды, оларсыз кез - келген жасушаның тіршілігі мүмкін емес; 2) жасушаның ерекше кьізметгерін камтама-сыз ететін және тек кейбір жасушаларда ғана актив болатын ген-дер. Оларды "молшылық" геңдері деп атайды.
Эукариоттар гендерінің экспрессиялануының реггелу жобасын 1972 ж. Г.П.Георгиев жасаған. Көпжасушалы ағзалардың ДНҚ молекуласының ұзындығы бір жасушалыларға қарағанда әлдеқайда ұзын, бірақ сол ДНҚ молеку-ласының бәрі бірдей а - РНҚ - ға айналмайды, себебі алғаш про-а-РНҚ (Гя-РНҚ) түзіліп, оныңтек азғана бөлігі-10% ғана, а-РНҚ-ға айналады. Адам геномы 2,9 хЮ6 нж түрады десек, одан пайда болған а-РНҚ мөлшері 2 хЮ* нж тең. А-РНҚ гендерінің орташа үзындығы 1800 нж тең десек адам геномында 110000-ға жуық струк-туралық гендер кездеседі (соңғы кездері адам геномындағы гендердің саны 30000 деп айтылуда).
Эукариоттар гендерінің экспрессиялануының реттелу механизмі прокариотгарға ұқсас. Дегенмен, оның кейбір ерекшеліктері де белгілі. Эукриоттар гендері экзон-интрон учаскелерінен тұрады. А-РНҚпісіп жетілуінде (процессинг, сплайсинг) интрондар үзіліп түсіп, экзондар бір-бірімен жалғанады. Сол сияқты, эукриоттар жасушасында ДНҚ молекуласы гистонды белоктармен өте тығыз байланыскан, ал бүл көп жағдайларда РНҚ-полимераза ферментінің әсеріне беріле бермейдіде а-РНҚсинтезделмейді.
Эукариотгар гендерінің транскрипциялануының иницияциясы прокариоттар сияқгы промотордан басталады. Промотор учаскесіне жақын жерлерде А-Т нуклеотидтері жиі кездесетін ген орналасқан, оның ұзындығы 20-30 нж тең. Бүл аймакды Гольдберг-Хогнесс боксы немесе *ТАТА...* тізбегі деп атайды. *ТАТА...* тізбегі РНҚ-полимераза ферментімен байланысып транскрипцияның басталу нүктесін аныктайды.
ДНҚ молекуласының транскрипциялану механизмі прокариоттар жасушасында жаксы зерттелінген.
Транскрипция журу үшін ДНҚ молекуласының жіпшелсрі бір-бірінен кем дегенде 2 өрім бойына ажырасуы қажет. Транскрипция промоторлық учаскеден басталып терминаторлық учаскемен аяқталады. РНҚ синтезін жүргізуші, яғни рибонуклеотидтерді бір-біріне жалғастырушы ферментті - РНҚ- полимераза ферменті деп атайды. РНҚ - полимераза молекуласының негізін 4 компаненттен (а, а (3 р,) тұратын корфермент құрайды. Корферментке бір белок -сигма (б) белок, жалғанса ол РНҚ-полимеразаның холоферментіне яғни актив күйіне айналады.
Сигма белок ДНҚ молекуласының промоторлық учаскесін дәл табады және бір-бірінен ажырасқан ДНҚ жіпшелерінің қайсысы матрица (калып) қызметін атқаратынын анықтайды. РНҚ синтезі басталғанны кейін сигма (б) белоктың кқжеті жоқ, сондықтан ол холоферменттен ажырап түсіп қалады, ал оның орнына басқа бір белтың - А генінің өнімі келіп қосылады. І Чиз-А генінің өнімінің қызметі ДНҚ молекуласының терминацялық учаскесін танып транскрипция процессін тоқтату. Синтезделінген РНҚ молекуласының ДНҚ спиральдерінен ажырап бөлініп шығуы үшін тағы бір белок қажет ол ро (р) фактор деп аталынады.
Эукриоттар ДНҚ-сының транскрипциясы, жалпы алғанда, про-кариоттарға ұқсас, дегенмен біршама ерекшеліктер де кездеседі. Мысалы, прокариоттарда РНҚ-ның 3 түрі де (а-РНҚ, т-РНҚ, р-РНҚ) бір РНҚ полимераза ферменті арқылы синтезделінеді, ал эукриоттарда олардың әрқайсысы тек өздеріне ғана сәкес РНҚ-полимеразалар арқылы синтезделінеді: р-РНҚ-РНҚ-полимераза -I арқылы; а-РНҚ-РНҚ-полимераза-ІІ-арқылы; т-РНҚ-РНҚ-по-лимераза- Ш-арқылы.
Прокариотгар гендерінің құрылысы қарапайым, ол тек мағыналы нуклеотид тізбектерінен тұрады және түгел транскрипцияланып бірден цитоплазмаға өтіп трансляция үшін матрица (қалып) бола алады. Ал, эукариоттардың гендерінің құрылысы күрделі, одан мағыналы және мағынасыз учаскелерден тұрады. Мағыналы учаскелерін экзондар, мағынасыз учаскелерін интровдар деп атай-ды.
Транскрипция процессінде гендердің экзондық және интрондық учаскелерінің бәрі де бірдей синтезделінеді оны про-а-РНҚ (Гя-РНҚ) деп атайды. Бірақ ол цитоплазмаға өтіп белок синтезі үшін матрица (қалып) қызметін атқара алмайды, себебі ол пісіп жетілуі қажет, яғни нағыз а-РНҚ-ға айналуы қажет. Про-а-РНҚ немесе Гя-РНҚ дан нағыз а-РНҚ пайда болуы үшін оның интрондары қиылып алынып тасталып экзондары бір бірімен жалғануы қажет. Бұл процессті процессинг және сплайсинг деп атайды. Пісіп жетілген а-РНҚ ядродан цитоплазмаға өтіп белок синтезі үшін матрица (қалып) қызметін атқарады.
Белок биосинтезі тіршіліктің ең маңызды қасиеттерінің бірі, себебі тіршілік процессінде белок молекулалары әртүрлі биохимиялық реакцияларға түсіп ыдырап жойылып отырады, ал олардың орнын толтыру тек жаңа белок молекулаларының синтезделуі нәтижесінде жүзеге асады.
РНҚ молекуласының нуклеотидтер тізбегінде жазылған ақпа-раттың колинеарлы полипептид молекуласының аминқышқылда-ры ретіне берілуін, яғни белок счнтезін, трансляция деп атаймыз.
Трансляция процессі полипептидтің N ұшынан басталып С ұшына қарай жүреді және ол метионин аминқышқылынан басталады. Яғни, кез - келген а-РНҚ молекуласының бас жағында метионин аминкышқылына сәйкес келетін (колинеарлы) код-инициаторлық код болады (АУГ). Полипептид синтезінің инициялануы ерекше т-РНҚ-инициаторлықт-РНҚ-ның қатынасуымен жүзеге асады. Бұл процесске 3 инициациялаушы факторлар - ІҒР ІҒ2, ІҒ3 қатынасады. Белок биосинтезінің жалғасуы, немесе элонгация сатысы, 2 фактордын қатысуымен жүреді - БҒ-1, ЕҒ-2, ал белок синтезінің терминациялануы кодон-терминаторларға және терминациялық факторларға-КҒ-1, К.Ғ-2-ге байланысты. Кодон терминаторлардың 3 түрі белгілі УАА, УАГ, УГА.
Трансляция процессіне 3 РНҚ-үшеуіде қатынасады. А-РНҚ-белок синтезі үшін матрица (кдлып) қызметін атқарса, рибосомалық РНҚ рибосомалардың құрамына кіреді. Рибосомалар цитоп-лазманың ең ұсақ субмикроскопиялық органеллалары болып табылады. Белок синтезі жүруі үшін рибосомалар а-РНҚ-мен байланысады. Рибосома 2 бөлшектен тұрады: рибосоманың кіші бөлшегі-305, рибосоманың үлкен бөлшегі-505. Әдетте, белок синтезі жүрмеген кезде, рибосоманың екі бөлшегі бір - бірінен бөлек кездеседі, олардың бір - бірімен қосылып біртұтас органоид - рибосома түзілуі белок биосинтезі басталған кезде жүзеге асады. А-РНҚ алғаш рибосоманың кіші бөлшегімен қосылады, содан кейін оларға инидияция факторлары ІҒр ІҒ,, ІҒ3 және т-РНҚ - метионин қосылып инициаторлық кешен түзіледі.
Инициаторлық кешен түзілгеннен кейін оған рибосоманың үлкен бөлшегі келіп қосылады. Рибосоманың үлкен бөлшегінде 2 учаске болады: пептидил учаске-П учаске, аминацил учаске-Аучаске. Ри-босоманың үлкен бөлшегінің П-учаскесіне формилденген метио-нинмен инициаторлықт-РНҚ орналасады, ал А-учаскеге а-РНҚ-ньң екінші кодонына сәйкес келетін антикодоны бар т-РНҚ өзінің аминкышқылымен орналасады.
Трансляция процессінде а-РНҚ кодондары аминқышкылдарымен тікелей әрекеттесе алмайды. Олардың арасындағы байланыс тасымалдаушы РНҚ (т-РНҚ) арқылы жүзеге асады. Әрбір аминқышқылының тек өзіне ғана сәйкес т-РНҚ-сы болады, сонымен кем дегенде 21 т-РНҚ белгілі-20-аминқышқылының 20 т-РНҚ асы және инициаторлық Т-РНҚ. Т-РНҚ-лар - т-РНҚфгн, т-РНҚд-' т-рНҚс<^ т-РНҚИ:і(>л, т-РНҚАсп т.с.с деп аталынады. Барлық прокариоттардың және эукариоттардың – РНҚ молекулалары 80- жуық нуклеотидтерден құралған және олардың соңғы құрылысы Ф рще ұқсас болып келеді. Олардың молекулаларының соңғы құрылысы беденің 3 құлақты жапырағына ұқсас. Т-РНҚ молеку-ласының 2 ерекше қасиеті белгілі: 1) т-РНҚ-ның 3'-ұшында ерек-ше рЦ рІД рА-ОН-тан тұратын бос нуклеотид тізбегі болады, осы ұшымен т-РНҚ-ға сәйкес келетін аминқышкылы ковалентті бай-ланысады; 2) т-РНҚ-ның қарама-қарсы ұшында 3 нуклеотидтен тұратын антикодон болады.
Т-РНҚ-ның негізгі қызметі-өзіне сәйкес аминқышқылын *танып* 3'-ОН ұшына қондыру және а-РНҚ-ның антикодонға ком-плиментарлы кодонын *танып* өзі алып келген аминқышқылын полипептид тізбегіне орналастыру болып саналады.
Аминқышқылының өзіне сәйкес-РНҚ молекуласыныңЗ^-ОН ұшына ковалентті байланысуы ерекше аминоацил т-РНҚ-синтета-за ферментінің қатысуы арқылы жүреді.
20 аминкышқылына сәйкес 20 аминоцил т-РНҚ синтетаза ферменттері болады. Аминоацил т-РНҚ синтетаза ферменттері өздеріне сәйкес келетін аминкышқылдарымен т-РНҚ-ның бір-бірін *танып*, аминқышкылының т-РНҚ молекуласының 3'-ОН ұшына коваленттік байланысуын қамтамасыз етеді. Бұл процесс 2 сатыдан тұрады. Алғашқы сатыда-аминкышқылдары активтенеді: Мысалы, Серилацил т-РНҚ-синтетаза+серин+АТФа(серилацил-АМФ) Се-рил-т-РНҚ-синтетаза+Ррі; Екінші сатыда активтенген аминқышқылы өзіне сәйкес т-РНҚ-ның 3'-ОН ұшымен байланысады.
(Серилацил-АМФ) серил-т-РНҚ-синтетаза+т-РНҚ а серил-т-РНҚСер+серил-т-РНҚ-синтетаза+АМФ.
Аминоацил-т-РНҚ-синтетаза ферменттері трансляция процесінің қателіксіз жүруін, яғни а-РНК,кодына колинеарлы (сәйкес) поли-пептид молекуласының синтезделуін, қамтамасыз етеді. Демек, трансляцияның қателіксіз жүруі аминоацил-т-РНҚ-синтетаза ферменттерінің өздеріңе сәйкес аминқышқылдарын *танып* оларды тиісті т-РНК,-ға қондыруына байланысты болады. Сонымен қатар, аминоацил~т-РНҚ-синтетаа ферменттері бақылаушылық қызметін де атқарады, Мысалы, изолейцил-т-РНҚ-синтетаза ферменті өзіне сәйкес т-РНҚ ормына басқа-*жат* т-РНҚ, айталық т-РНҚ**", *таныды* делік. Осының нәтижесінде изолейцин-т- изолейцил-т-РНҚфен кешені пайда болар еді. Осы Кателіктің нәтижесінде синтезделуші белок молекуласында фенилаланин аминқышқылының орнына изолейцин аминқышқылы орналасар еді. Бірак, фенилаленил-т-РНҚ-синтетаза ферменті өзінің т-РНҚФЕН - сымен *жат* аминқышкылының қосылғанын сезіп қалып қате кешенді - изолейцин аминқышкылына және т-РНҚ*"1-ге ыдыратады.
Аминоацил-т-РНҚ-синтетаза ферменттерінің екінші бақылау-шылық қызметі аминқышқылдардың активтену сатысында байқа-лады. Айталық, изолейцил-т-РНҚ-синтетаза ферменті изолейцин аминқышқылының орнына валин аминқышқылын активтендірді делік.
Изолейцил-т-РНҚ-синтетаза+валин+АТФа(валилацил-АМФ) изолейцил-т-РНҚ-синтетаза+Ррі. Соңда, келесі сатыда түзілген кешен т-РНҚИті мен әрекеттескенде валил-т-РНҚИюп пайда болуының орньша валил-АМФ, еркін валин аминқышқылына және АМФ-ге гидролизденеді.
(Валилацил-АМФ) изолейцил-т-РНҚ-синтетаза+т-РНҚИіола валин +АМФ+т-РНҚИтоі+изолейцил-т-РНҚ-синтетаза.
Сонымен, аминоацил-т-РНҚ-синтетаза ферменттері нақтылы аминқышкылдарын өздеріне тиісті антикодондармен байланыстырып генетикалық ақпараттың трансляциялану процессінде маңызды роль атқарады. Сонымен қатар, бұл ферменттердің қос қабат бақылаушылык кызметі нәтижесінде трансляция процессінің қателіксіз жүру денгейі едәуір жоғарылайды. Мысалы, аминқышқылдардың *қате* активтенуі шамамен 100 изолейцин молекуласынан 1 валинға тең болса, активтенген аминқышқылының *жат* т-РНК, мен байланысуы 1/180 тең, сонда трансляцияның жалпы қателігі 1/100 х1/180=1/18000-нан аспайды.
Аминоацил-т-РНҚ кешені пайда болғаннан кейін олар рет-реттерімен тізіліл орналасып, аминқышқылдар арасында пептидтік байланыс пайда болады. Оны рибосоманың үлкен бөлшегінде кездесетін ерекше фермент — пептидилтрансфераза ферменті қалыптастырады. Екі аминқышқылдары арасында пептидтік байланыс пайда болғаннан кейін рибосома бір кодонға солға қарай жылжиды, осының нәтижесінде инициаторлық кешен рибосомадан сыртқа шығады, А учаскедегі кешен П учаскеге өтеді, ал А учаскесіне а-РНҚ-ның үшінші кодонына сәйкес келетін т-РНҚ өзінің аминқышқылымен келіп орналасады, содан кейін екінші және үшінші аминқышқылдары арасында пептидтік байланыс пайда болады. Бұл жерде элонгация факторлары-ЕҒ-1, ЕҒ-2 қатынасады. Содан кейін рибосома тағы бір кодонға солға жылжып А учаске босайды. Осылайша белок синтезі А учаскеге а-РНҚ-ның терминаторлық кодондары келіп жеткенге дейін жалғаса береді. Терминаторлық кодондарды (УАА, УАГ, УГА) арнайы терминация факторлары-КР-1, ЯҒ-2 *таниды*. Терминаторлық кодондарға сәйкес келетін аминқышқылдары болмағандықтан А-учаске бос қалып, пептидил трансфераза ферменті пептидтік байланыс жасай алмайды, сондықтан ри-босома 2 бөлшекке ыдырайды, синтезделген полипептид бөлініп шығады. Осылайша белок синтсзі аяқталады.
Бақылау сұрақтары
1. Гендердің экспрессиялануының (активтенуінің) реттелу механизмдері.
2. Гендердің экспрессиялануының реттелу механизмдері.
3. Транскрипциялану немесе РНҚ синтезінің механизмдері
4. Белок биосинтезі
Дәріс № 13,14
Клеткалардың іn vіtro жағдайында өсуі және морфогенездің жолдары. Органогенез
Жоспар:
Клеткалардың іп vitro жағдайында дифференциялануы морфогенезі
Жасушалардың регенерациялану ерекшеліктері.
Жасанды ортада регенерант өсімдігі пайда болу құбылысы
Өзіне тән бір қалыпты өсуімен және көбеюімен сипатталынатын ретсіз өсетін каллус клеткалары қайта дифференциялану процестері арқасында, ұлпалар (гистогенез), мүшелер (органогенез) және ұрық тәріздес құрылымдар-эмбриондар (эмбриогенез, немесе сомалық эмбриогенез) түзе алады.
Бастапқы кезде каллус өсіп, массасы қандай да бір ауыспалы кезеңге жеткеннен кейін морфогенезге ауысады. Бірақ, тек бірлі жарым санаулы клеткалар ғана, өзінің даму бағдарламасын өзгертеді. Бұл, мына жағдайды көрсетеді, ретсіз өсетін каллус клеткалары морфогенезге өту үшін , сыртқы және ішкі факторлардың біріккен әрі үйлескен әрекеті қажет. Сөйтіп, бірен-саран комлементті клеткалар сыртқы факторлардың әсерін (түрлі индукторлар) қабылдап, белгілі бір даму жолына түсуге дайын болып тұрады, яғни детерминацияға өтеді. Ал содан кейін олар өзінің тұқым қуалау бағдарламасын жаңа бағытпен жүзеге асыра бастайды, ақырында каллус клеткалары редифференцияланады. Редифференциялану деген дифференцияланған клеткалардың бір маманданған жағдайынан басқа жағдайға ауысуы. Ол өте күрделі процесс, сыртқы және ішкі факторлардың катысуымен реттеледі.
Ретсіз өсетін каллус ұлпасынан ұйымдасқан құрылымдарға, атап айтқанда өскінге, тамырға, эмбриондарға бастама болатын меристемалық аймақтардың пайда болуына әсер ететін факторлардың мәні туралы анық түсінік әлі жоқ.
Мүмкін, клеткалардың дифференциялануын қозғайтын және олардың гистогенезге өтуін, одан кейін мүшелер түзетін құрылымдардың түзілуінің негізгі себептерінің бірі, фотогормондардың әсері деуге болады. Дегенмен морфогенездің қандай да бір типін анықтайтын негізгі факторлардың бірі ол қоректік ортадағы ауксиндер мен цитокининдердің ара қатынасы. Бұл заңдылықты алғаш рет Скуг пен Миллер 1957 жылы ашқан, одан кейінгі жылдары көптеген тәжірибелер арқылы дәлелдетілген.
Ауксин мен цитокининнің ара қатынасы орташа болғанда темекі өсімдігінің каллус ұлпасында морфогенез процессі байқалмайды, бірақ клеткалардың бөліну арқылы көбеюі (пролиферациясы) жүреді. Ортада ауксиннің цитокининге ара қатынасы жоғары болса, тамыр түзілуін (ризогенезді) қоздырады, ал ортада ауксиннің цитокининге ара қатысы төмен болған жағдайда бүршіктердің пайда болуын (геммогенез) ынталандырады. Сонымен бір каллустың өзінде әр түрлі морфогендік құрылымдар түзіле алады. Бұл жағдайлар кактустық ұлпаларда фитогормондардың кондентрациялық градиенті (концентрацияның үздіксіз өзгеруі) түзілетіндігінің дәлелі бола алады. Былайша айтқанда, каллустың әр түрлі бөліктеріндегі фитогормондар мөлшері әр шамада болғандықтан, ол морфогенездің қандай да бір типінің басталуына себеп болады.
Сөйтіп пайда болатын меристема типі ортадағы ауксин мен цитокининнің ара қатынасына байланысты екені белгілі, бірақ морфогенездің түрлі этаптарында қайсысының қатынасатындығы туралы анық дерек жок, Алайда, бірқатар өсімдіктер каллустарынан ортадағы фитогормондар балансын қанша өзгерткенмен бүршік немесе тамыр өсіріп алу мүмкін емес. Бүршіктің немесе тамырдың дифференциалануы гормондар әрекеттесуден реттеледі десек те, оған басқа да факторлар, мысалы: қоректік ортадағы қанттар мен фосфаттардың мөлшері, азоттың көзі, пуриндер т.б. ортаның құрамына кіретін компонеттер әсер етуі мүмкін.
Өткізгіш жүйенің элементтері пайда болуына тек қана гормондар әсер етіп қоймайды, сонымен бірге қанттар да ықпал етеді. Каллус ұлпаларына ауксин - қант ерітінділерін инъекция жасағанда өткізгіш элементтері түзілген. Ол ауксин концентрациясына да және қант концентрациясына да тәуелді болады. Сахарозаның төмен концентрациясын (2 %) енгізгенде көбінесе ксилема элементтері, ал жоғары концентрациясында (8 %)-флоэма элементтері пайда болады. Сахарозаның орнына глюкозаны колданғанда гистогенез жүрмеген. Скуг және Миллер ашқан факт, ол темекінің каллус ұлпаларынан жасанды ортада бүршіктер мен тамырдың өсуін экзогендік фитогормондар балансы арқылы реттелетіні, морфогенезді іп vitro жағдайында зерттеуде маңызды роль атқарды. Дегенмен, бұл құбылыстың, клеткалық және молекулалық деңгейдегі механизмі әзірше анық емес.
Фитогормондардың морфогенезге әсерінің механизмдері туралы нақтылы білім жок, сондықтан жаңа өсімдіктерді жасанды түрде қолдан өсіргенде қайтадан гормондардың үйлесімді арақатынасын және құрамын іріктеп алу керек. Астық тұқымдастар және кейбір бұршақ тұқымдас өсімдіктердің клеткалары мен протопластарын іп vitro жағдайында өсіргенде, оларда морфогенез процессінің басталуына әсер ететін, оңай әрі регенеранттарды алу үшін гормондардың қажетті концентрациялары белгісіз. Осындай фактілер және де көптеген әдебиет деректеріне карағанда, клетканың дифференциалануын және морфогенезді реттеуде фитогормондардың ерекшелігі жоқ деген түсінік туындайды да, морфогенездің гормондық теориясына сенімсіздік пайда болады. Т. Мурасигенің көрсетуі бойынша Скуг пен Миллердің концепциясы органогенез процессі жөніндегі түсінікті жеңілдетеді. Даму бірнеше этаптардан тұратын, өте күрделі процесс, этаптардың әрқайсысында гормондарға, жарыққа, температураға қойылатын талаптар да өзгеріп отырады. Мурасигеннің пікірі бойынша, өсімдіктердің жасанды ортада өсуіне әсер ететін басқа да заттарды ескеру қажет, ол заттарды ортаға клеткалар мен ұлпалардың өздері бөледі, олар улы болғандықтан, морфогенезді тежеуі мүмкін. Сөйтіп, іп жағдайында өсетін клеткалардың дифференциалануы мен морфогенезінің барлық реттелу бағыттарын, жалғыз гормондық концепциямен түсіндіру мүмкін емес. Коректік ортадағы өскен ұлпалар мен клеткалардың морфогенезге өту қабілеті негізгі өсімдіктің генотипіне, оның физиологиялық күйіне, эксплант алынған мүшеге, жыл мерзіміне тағы басқа химиялық және физикалық факторларға тәуелді.
Р. Г. Бугенко өз лабораториясының осы бағыттағы жұмыстарының нәтижелерін және әдебиеттердегі бар деректерлі тұжырымдай келіп, каллустардағы меристемалық аймақтардың пайда болуын төмендегідей баяндайды. Индуктордан келетін снгналды қабылдап алуға қабілеті бар клеткалар көп емес, олардың ерекше бір даму жолына түсуге дайындылығы ертеден белгіленеді. Бұл клеткалар детерминация кезеңінде бөлінуін токтатады, қалыңдау клетка қабығын түзіп, өзін қоршап тұрған каллустық клеткалардан оңашаланады. Бұл кезеңде клеткалар кейінгі өзінің тез бөлінуіне кажет субстрат және энергия жинақтайды. Паренхима тәрізді каллус клеткалары массасынан пайда болған бастама клеткалар, өзін қоршап тұрған клеткаларға қарағанда морфология жағынан өзгеше болады. Олардың ядросы үлкейеді және сол сияқты ядролық - плазмалық арақатысы да, рибосоманың, митохондрияның саны көбейеді, пластидтерінде крахмал жинақталады, Сөйтіп, өсімдіктердің бастапқы маманданған клеткалары каллусқа айналған кездегідей /дедифференцияланғанда/, каллустың клеткасы меристемалық клеткаға айналғанда толығымен өзінің ішкі құрылымын қайта құрайды. Біраз уақыт өткеннен кейін, бұл клеткалар бөлшектену типімен тез бөліне бастайды да, меристема типті ұсақ клеткалардың сфералық массасын түзеді. Одан әрі қарай бұл клеткалар массасы өскін немесе тамыр бастамалары түзілетін меристемалық аймаққа айналады немесе биполярлық глобулярлы проэмбрио / ұрық бастамасы / түзіледі де, одан эмбриоинды - ұрық тәрізді құрылым өсіп жетіледі. Дегенмен әрі қарай дамудың өзгешелігін полярлық / қарама-карсылық / 'белгілейді. Жасанды ортада өскен каллустар клеткалардың бөлінуі ретсіз, әрі әр түрлі бағыттарда жүреді де, ұлпалардың бытыраңқы массасы пайда болады, яғни каллустарда белгілі полярлық өсі жоқ. Ал, өскін-тамыр меристемаларының, клеткаларының бөлінуі қатаң түрде бір ретпен жүретіндігімен
іп vitro жағдайында өсіретін клеткалар өсімдіктер биотехнологиясында екі бағытта қолданылады: біріншісі- клеткалар массасы, яғни каллус; екіншісі- регенерант өсімдіктері. Сөйтіп, өсімдік клеткаларын жасанды қоректік ортада өсіргенде регенерация процессі негізгі іргелі процесс, себебі онсыз өсімдіктер биотехнологиясындағы көптеген салалары жүзеге аса алмайды.
Жасанды ортада регенерант өсімдігі пайда болу құбылысы дәлелденгендей, маманданған өсімдік клеткалары барлық генетикалық қасиеттерін сақтап қалады, яғни тотипотентті болады. Тәжірибе жүргізуде талай өсімдіктер түрінің жеке клеткаларының тотипотенттілігі анық көрсетілгенімен, зерттеушілерідің бәрі бірдей, тотипотенттілік қасиет өсімдіктің кез келген клеткаларына тән деп есептемейді. Оған мысал ретінде тотипотенттік қасиетін көрсетуі қиын объектілер мағынасында, көптеген өсімдіктердің жасыл жапырақ табақшасы клеткаларын, астыктұқымдастарда-оқшауланған протопласттарды келтіруге болады. Бұдан басқа қоректік ортада ұзақ уақыт өсірілген штаммдар да морфогенезге деген қабілетін жояды, астыктұқымдастар ұлпалары жасанды ортада өсіру кезеңінде сол қабілетін өте тез жоғалтады. Қоректік ортада ұзақ уақыт өсіру кезінде, ядролық және цитоплазмалық гендері қайта құрылған клеткалар популяциясы қалыптасып ,ал олар клеткалардың қайта дифференциялануына және морфогенезге өтуіне тиым салып, жол бермеуі мүмкін. Тәжірибе жүзіндегі морфогенездің, өз алдына қоятын мақсаты, ол дифференциялану және регенерация процестерін іп vitro жағдайында басқару мәселелері.
Бақылау сұрақтары:
Клеткалардың іп vitro жағдайында дифференциялануы морфогенезі
Жасушалардың регенерациялану ерекшеліктері.
Жасанды ортада регенерант өсімдігі пайда болу құбылысы.
Органогенез
Дәріс №15,16
Өсімдіктердің экономикалық маңызы зор заттарын өндірудің клеткалық технологиялары
Жоспар:
1. Экономика жағынан маңызды заттар алудын клеткалык технологиясы
2. Сұйық қоректік ортада өсетін клеткалар және олардың ерекшеліктері.
3. Клеткаларды қоректік ортада өсіріп қосымша метаболизмде түзілетін заттар алу жұмыстарының этаптары.
Өсімдіктер құрамында орасан зор көп әр түрлі қосымша түзілетін заттар бар. Қосымша заттар деп аталғанмен, олар өсімдіктегі заттардың алмасуында маңызды роль атқарады. Олардың көпшілігі медицинада және техникада кеңінен қолданылады, тамақ және парфюмерия өндірісінде пайдаланылады.
Ұлпаларды қоректік ортада өсіргенде, зерттеушілер назары осы заттардың арасында, бірінші болып алкалоидтарға түспек, шыгу тегі әр түрлі өсімдіктердің /қызғылт қабыршөп, жыланды раувольфия, қара мендуана, сасық мендуана т. б./ каллус ұллпаларын зерттегенде олардың кұрамында алкалоидтар бар екендігі анықталған. Көптеген ғалымдардың ерен еңбектерінің арқасында, каллус ұлпаларының басқа биологиялық активті заттарды синтездеуге қабілетті екені туралы біршама құнды деректер жиналды. Жасанды қоректік ортада өскен өсімдік клеткалары, өсімдіктің негізгі түріне тән, қосымша метаболизмде түзілетін заттардың кең спектрін синтездеу қабілетін сақтайды. Сол заттарға жататындар:
1/ алкалоидтар;
2/ терпендер мен терпеноидтар;
3/ гликозидтар;
4/ полифенолдар;
5/ пслисахаридтер;
6/хош иісті майлар;
7/ ерекше пептидтер;
8/ табиғи бояулар;
9/ стероидтар;
10/ дәмділік заттар;
11/ инсектицидтер;
12/ балауыздар;
13/ витаминдер т.б.
Экономикалық маңызды заттар алу үшін, өнім беретін клеткалар мен ұлпаларды өндіріс жағдайында өсіру-биотехнологиясының жаңа бағыты. Егер дәстүрлі биотехнологияда құнды биологиялық активті заттар алу үшін тұтас организмдерді: микроорганизмдер, өсімдіктер, жануарлар пайдаланылып келсе, ал казіргі биотехнология- клеткалық биотехнологаяға бағытталған, тап осы жағдайда өсімдіктердің ерікті және орнықтырылған /иммобильденген/ клеткаларын жасанды ортада өсіруге негізделген. Қоректік ортада өскен клеткаларды биосинтездік өнеркәсіпте қолданудың үш бағыты суретте көрсетілген.
-
суспензия
Орнықтырылған клеткалар
Экономикалык маңызы бар дәстүрлі өсімдіктер өнімдерін синтездеу |
|
БИОТРАНС-ФОРМАЦИЯ |
|
негізінде жаңа өнімдер синтездеу |
-
Ферменттер әсерінен кейінгі процесстер
-
қажет өнімдер
Қоректік ортада өскен өсімдік клеткаларын пайдаланып маңызды өлшемдер алу жолдары.
Қоректік ортада өскен клеткалардың ферменттері қатысуымен арзан және коры мол бастапқы заттардан биологиялық активті косылыстарды синтездеуге болады, бұл процессті биотрансформация деп атайды.
Қоректік орта кұрамын және баскада өсіру жағдайларын өзгерту алынатын өнімнің мөлшерін өзгертіп қана қоймайды, сонымен бірге сапалық құрамы да өзгереді, нәтижесінде басқаша әсер ететін мүлде жаңа қосылыстар алынуы мумкін. Жапондық зерттеушілер іп vitro жағдайында мынадай жаңа биологиялық активті заттар: ерекше пентиндтер, канцерогендерге қарсы колданылатын қосылыстар, убихинон - 10, агар - агар т, б. алуда осылай табыска жетті. Клеткалық биотехнологияның келесі қадамы, иммобилъденген, яғни орнықшрШЕгаи өсімдік клеткаларын пайдалаку.
Қоректік ортада өсірілген клеткалар өндіріс жағдайында пайдаланатын объектілерге айналу үшін, олар табиғи жане модеңп, дәрілік, тағамдық өсімдіктердің, сонымен бірге микробиологиялық өндірістің бәсекегінен тыс болуы керек. Дәстүрлі өсімдіктер шикізатын алуымен салыстырғанда, қоректік ортада өсетін клеткалрдың бірнеше артықшылықтары бар: 1/ өнімді алу климатқа, жыл мерзіміне, ауа-райына, топырақ жағдайына байланысты емес; 2/жасанды ортада өсірудің ең жақсы жағдайларын стандарттауға болады; 3/ барлық процесстерді толық автоматтандырып компьютерлік бақылауға алуға мүмкіндік. Экономикалық маңызы бар заттардың негізгі көзі өсімдіктер екенін ескерсек, ал өсімдік шикізаттарының қорлары түгел планета бойынша азаюда, сондықтан клеткалық технологиялардың болашағы алда екенін көз алдымызға елестету қиын емес. Жасанды ортада өскен ұлпалар мен клеткаларды пайдаланып, ғылыми түрде лабораторияда жасалған тәжірибелерді өндіріске енгізу жұмыстары жаңадан басталуда. Жоғары рентабельдік технологиялар жасауға қол байлау болып отырған өсімдік клеткасындағы қосымша метаболизмнің генетикалық, биологиялық реттелуін анықтайтын іргелі білімнін, жетіспеушілігі.
Экономика жағынан маңызды заттарды өсімдіктерден алу үшін, микробиологиялық өндірісте қабылданған арнайы технологиялар мен аппараттар қолданылады, сондықтан сұйық қоректік ортада өскен клеткалар пайдаланылады. Сұйық қоректік ортада өскен клеткалардың қоректену жағдайларының біркелкі болуынан, таза ауамен канықтыру және зиянды метаболиттерді клеткадан кашықтатуға байланысты клеткалардың өзгергіштігі де аз болады. Оның маңызы, бағалы өнім беретін клеткаларды өндіріс жағдайында өсіргенде, штаммның негізгі қасиетін сақтау керек болғанда өте зор.
Жоғарғы сатыдағы өсімдіктердің өз алдына тіршілік ететін ретсіз өсіп жатқан клеткалар популяциясы ерекше жүйе, ол өзінің биологиясы жағынан өсімдік ұлпалары клеткаларынан да, микроорганизмдерден де өзгеше. Клеткаларды сұйық қоректік ортада әр түрлі жағдайларда өсірілген, популляцияның өсу заңдылыктарын зерттеу теориялық және практикалық мәселелерді шешуде маңызды рол атқарады.
Жеке клеткаларды өсіру екі этаптан тұрады: алдымен өсімдік ұлпаларынан, негізінде каллус ұлпасынан, зақымдалмаған тіршілікке икемді клеткаларын оқшаулап алады, одан кейін клеткалардың өсуіне және бөлінуіне қолайлы жағдайлар жасалады. Сұйық ортада өсетін клеткалар алудың үш әдісі бар: 1/ Сұйык қоректік ортаға борпылдақ дпфференцияланбаған каллусты ауыстыру ең колайлы әдістердің бірі, айналмалы шайкағышты сұйық ортада өсіргенде каллус оңай ыдырап кетеді. Ондай каллустарды қоректік ортадағы гормондардың арақатысын реттей отырып өсіріп алуға болады. Мысалы: ортада ауксиондер / жақсысы 2,4 -Д / мөлшері көп, ал цитокининдер және кальций аз мөлшерде / жоқ болса тіпті жақсы /, болса қолайлы жағдай. 2/ Бастапқы экспланттарды, мысалы, тозаңқаптарды сұйық қоректік ортаға салып каллусогенез процессінің өзін сол ортада жүргізеді. 3/ қатты тығыз каллусты кесіп, бөлшектеп, біріккен клеткаларды ажыратқаннан кейін сұйық қоректік ортада өсетін клеткалардың көзі болу мүмкіндігі бар. Жеке клеткалардан және ұсак, агрегаттардан / біріккен клеткалар/ тұратын суспензияны алу үшін клеткаларды саңылаулары белгілі елек арқылы сүзіп алады, не болмаса суспензиядағы ірі агрегаттарды минут сайын тұндыру жолымен бөліп алады. Ортадағы жекеленген клеткалар мен ұсақ агрегаттардың мөлшерін көбейту үшін, суспензияның жеңіл фракцйяларынан алынатын инокулюмді жиі-жиі жаңа ортаға ауыстырып отыру керек. Инокулюм - жаңа қоректік ортаға отырғызу үшін пайдаланылатын суспензияның бір бөлігі. Клеткалардың диссоциациялану дәрежесіне қоректік ортаның құрамы, ауамен қанықтыру, суспензияның араластырылуы әсер етеді. Қанша күш салғанмен біркелкі жеке клеткалардан тұратын суспензияны алу мүмкін емес. Ең жақсы жағдай жасалғанның өзінде жеке клеткалар саны 60% аспайды, ал қалған бөлігі 2 ден 10 клеткаға дейінгі топтар және көп клеткалы агрегаттар үлесіне тиеді. Жеке клеткаларға карағанда топтасқан клеткаларда клеткалардың бөлінуі жиі байқалады. Суспензияның жоғары диссоциацияланған фракциясын өсіру процессі кезінде ол өзінің өнімділігін төмендетеді. Осыған қарай жекеленген және агрегаттанған клеткаларды аз көрсеткіштері бойынша айырмашылықтары мол субпопуляция деп қарауға болады. Суспензиядан клеткалардың әртүрлілігі тек морфологиясы жағынан емес, сонымен бірге биохимиялық қасиеттері, физиологиялық күйі және генетикасы жағынан да гетерогенділігі. Өндірістік мақсаттар үшін, бірнеше қайталап отырғызып өсіру кезінде клеткалардың эпигенетикалық көрсеткіштерін өзгеріссіз бір деңгейде сақтайтын клеткалар бағалы. Болашақтағы клеткалық технологиялардың объектісі гомоплоидтық, өнімділігі бір қалыпты, бөлінуі синхронданған жеке клеткалар болуы керек.
Өсімдік клеткаларының қатты целлюлоза- пектин қабығы және үлкен вакуолі болғандықтан, қабығы сынғыш, механикалық өсерге өте сезімтал, сондықтан суспензияны ұқыптап араластыру керек.Араластыру және ауамен қанықтыру үшін айналмалы шайқағышты қолдану немесе зарарсызданған ауаны коректік ортаға үрлеу арқылы жүргізіледі.
Жүзгін клеткаларды үлкен көлемді сұйық коректік ортада өсіруге арналған аппараттарды ғалымдар жыл сайын жетілдіріп жатқанын айта кету керек. Егер алғашқы жұмыстар жүргізілген кезде клеткаларды сұйык қоректік ортада өсіру үшін қарапайым роллерлі /дөңгелегіш/ қондырғылар және қуаты аз шайкағыштар қолданылған болса, қазір көлемі үлкен микробиологиялық ферменттерлер қолданылуда. Клеткаларды сұйық қоректік ортада өсірудің бірнеше әдістері бар.
Клеткалардың бір бөлігін /инокулюмді/ жабық ыдыстарда көлемі бір қалыпты сұйық қоректік ортада өсіруді жиналмалы немесе мерзімді өсіру деп атайды. Бұл жағдайда клеткалар өсетін жүйе газдар мен клеткалық метаболизмде ортаға бөлінетін үшқыш заттардан басқаға жабық. Сұйык коректік ортадағы клеткалардың өсуі бәсеңдегенде, ортадағы клеткалар тығыздығы бастапқы деңгейге жеткенше жаңа дайындалған коректік орта қосылады да клеткаларды өсіру циклі кайталанады. Инокулюмді жаңа коректік ортаға отырғызғаннан бастап келесі қайталап жаңа коректік ортаға отырғызуға дейінгі уақыт аралығын клеткаларды өсіру циклі деп атайды, ал өсу циклі деп, қоректік ортадағы клеткалардың өсуінің стационарлық фазаға шығуына дейінгі мерзімді атайды. Клеткаларды қоректік ортаның ағынында өсіру - үзіліссіз өсіру деп аталады, оның үш түрі болады: 1/ ағынды жабык, жүйеде өсіру- клеткалар үздіксіз жаңа қоректік ортамен қамтамасыз етіліп тұрады, клеткаларға түсетін орта көлемі мен пайдаланылған ортаның ағып шыққан көлемі бірдей болады, ал клеткалар сол ортада кала береді; 2/ ағынды-ағынсыз жүйеде өсіру- сұйық қоректік ортада өскен клеткалар суспензиясының бір бөлігі әлсін-әлсін алынып тұрады, қалғанына нақ сондай мөлшерде жаңа қоректік орта құйылады; 3/ ағынды ашық жүйеде өсіру- үздіксіз ағып түсіп жатқан жаңа коректік орта мен сол ыдыстан алынған өскен клеткалар суспензиясы көлемдерінің тең болуы.
Бұдан баска клеткаларды үзіліссіз өсіру үшін турбидостат және хемостат әдістерін қолдануға болады. Турбидостат ережесімен үздіксіз, тыстан ешқандай шектеусіз өсіру аркылы клеткалар популяциясының өсу карқындылығы белгілі бір деңгейде ұсталады, өсу оптикалық тығыздығын тексеру арқылы бақылайды. Бұл әдісті клеткаларды өсірудің бастапқы кезеңінде қолданады. Клеткалар суспензиясын хемостат әдісімен өсіргенде, клеткалар популяциясының өсуі мен тығыздығы құрамында өсуді тежейтін факторы бар қоректік орта арқылы реттеледі. Хемостат принципі клеткалардың өсуінің экспоненциялдық фазасының соңғы кезеңінде қолданылады.
Р.Г.Бутенко, өсімдіктердің экономикалык жағынан тиімді және құнды заттар алу үшін, клеткалық технологияларды жан-жақты зерттеп, олардың алдағы уақыттағы мүмкіндігін, болашақта қол жеткізуге болатын табыстарын төмендегідей деп тұжырымдайды:
Қазіргі технологиялар: |
|
Келешектегі технологиялар: |
|
|
|
Жабайы типті клеткалардың штамдары
|
|
Мутант және қолдан кұрастырылған клеткалардың штаммдары |
Клеткаларды жиналмалы (мерзімді) өсіру
|
|
Клеткаларды ортанын үзілмейтін ағынында өсіру
|
Оттегі бірқалыпты |
|
Оттегі мөлшерін көбейту. |
Клеткаларды сұйық ортада өсіру (суспензия)
|
|
Иммобильденген клеткаларды өсіру. |
Әдеттегі бақылау. |
|
Процесстерді компьютер арқылы бақылау және басқару. |
1. Экономикалык жағынан тиімді өсімдікті таңдап алу жасанды қоректік ортада өсіру үшін таңдап алынған өсімдіктердің құрамында айтарлықтай мөлшерде құнды, экономика жағынан тиімді қосымша заттар болу керек. Бұл келтірілген қағиданың әсіресе сирек немесе жойылып бара жатқан өсімдіктерге қатысы бар.
2. Ұлпаларды жасанды қоректік ортада өсіруге алғаш рет енгізу. Бұл максат үшін, құрамында ең көп мөлшерде ізделіп отырған заты бар жеке өсімдіктер таңдалынып алынады. Осының мысалы ретінде өз қызметкерлерімен бірге Ценк жасаған жұмысты келтіруге болады. Олар құрамында индол тектес алкалоидтар серпентин мен аймалициңді көп мөлшерде ұстайтын қызғылт қабыр шөпті /Саіһагапіиз гозеиз - барвишок розовый/ жасанды қоректік ортада өсіру үшін әр түрлі аймактарда өскен өсімдіктерден 184 тұқым үлгісін жинаған. Осы жиналып тұқымдардан өсіп шықкан өскіндерден тек кейбіреулері, құрамы жағынан алкалоидтарға байлары / кұрғак салмакка шаққанда 0,7 % кем емес/ ғана іріктелініп алынған, олардан каллус ұлпалары өсіріліп алынған. Әдетте, бастапқы каллус ұлпаларын қатты қоректік ортада өсіріп алады.
3. Алынған биомассаның сандық және сапалық құрамын химиялық әдістер арқылы зерттеу. Өсімдіктердің ретсіз өсіп көбейетін каллустық клеткалары косымша түзілетін қосылыстарды тек бір ерекше жағдайда болмаса, тұтас өсімдіктердің кор жинайтын мүшелеріне қарағанда кем ұстайды. Бұндай жағдай бүтін организмде қосымша түзілетін заттардың синтезі цитодифференцировканың бакылауында болатындығын көрсетеді. іп vitro өсетін клеткалардың дифференциялануының басынуы олардағы қосымша заттардың биосинтезделу мүмкіндігін төмендетеді. Клеткалардың генетикалық мүмкіндіктерін толық жүзеге асыра алмауы басқа себептерге байланысты болуы мүмкін. Мысалы, қоректенудің гетеротрофтык типіне кезінде, автотрофтық қоректенудің бұзылуынан. Бұдан басқа да факторлардың әсерінен өнімнің сапалық құрамы өзгеруі мүмкін. Кейбір өсімдіктерде қосымша заттардың биосинтезі олардағы өсу процессі бәсеңдегенде басталады, сондықтан оларды екі сатылы коректік ортада өсіру тәртібі қолданылады, бастапқы кезінде клетка массасының карқынды түрде өсуіне, одан кейін қосымша заттардың биосинтезделуіне жағдай жасалады.
4. Үйлесімді қоректік орта мен өсіру жағдайларын іріктеу.Қосымша заттар алмасуына қатысы бар генетикалық информацияларды толық жүзеге асыру үшін, арнайы жағдайлар жасалуы керек. Пайдаланылатын коректік орталар клеткаларға үйлесімді екеніне толық сенім болмағандықтан, әрбір нақты жағдайда коректік ортаны т.и. факторларды басқа зерттеушілердің тәжірибелеріне сүйене отырып таңдап алады. Бұл жұмыс іп vitro өскен клеткаларды сұйык ортаға ауыстырғанда бұрынғыдан да күрделене түседі, оның себебі сұйык, ортада аэрациялау және шайқап араластыру т.б. факторлардың әсерін ескеру керек.
Қоректік ортаның құрамына кіретін косындылардың ішінде қосымша метаболизмге әсер ететін негізгілері фитогормондарға жүктеледі. Бірақ фитогормондардың әсер етуі олардың түрше, түзетін заттың табиғатына, штаммға т.б. тәуелді, сондықтан фитогормондарды қолдану үшін дайын ереже болмайды. Фитогормондардың әсері жанама тілде клеткалардың дифференциялануы арқылы жүруі мүмкін, дегенмен іп vitro жағдайында өсу процесстері мен қосымша заттар түзілуінде арасында кандай да бір байланыстың болуы барлық уақытта байқала бермейді. Трофикалық факторлары ішінде қосымша түзілетін заттар биосинтезіне анық әсер ететін сахароза және сол қосылыстардың биосинтез жолындағы кейбір алғы заттары. Ұлпаларды ұзақ уақыт қоректік ортаның шектелген бір көлемінде өсіргенде, клеткалардың органикалық заттар синтездеу қабілеті томендейді, сол уакытта қоректік ортаға қосымша синтезделетін қосылыстардың алғы заттарын қосса, олар қосымша заттардың биосинтезіне әсер етеді деп, болжауға болады. Алғы заттар ретінде амин қышкылдарын / глутамин, фенилаланин, лейцин, орнитин т.б. / жеке органикалық қышқылдарды /ацетат, сукцинат, шиюім т.б./ қолданcа болады, Бірақ, каллустық ұлпалар олардың бәрін әр уақытта өз қажетіне жарата бермейді. Кейбір жағдайда қоректік ортаға алғы заттарды қосканмен одан жақсы нәтиже алына бермейді. Оны мынадай себептермен түсіндіруге болады: оларды клеткалар сіңірмейді, себебі, клеткалардың өзі синтездеуі мүмкін немесе қоректік ортаны дайындау, зарарсыздандыру барысында ыдырап кетуі, болмаса физиологиялық активтілігі жойып басқа формаға айналуы әбден мүмкін.
Әдебиеттерде қоректік ортаның минералды кошюненттеріпін және орта рН- ның кейбір жеке қосымша түзілетін өнімдердің мөлшеріне әсер ететіні туралы деректер бар. Физикалық факторлардың ішінен жарыкты, температураны, оттегінің, көмірқышқыл газының парциялдық қысымын келтіруге болады. Жарық әсерінің түрге ерекшелігі бар. Жасанды ортада өскен ұлпалардың кейбіреуінде бірқатар қосымша заттардын /алкалоидтар, гликозидтер, эфир майлары, флавоноидтар/ синтезі жарыкта ұлғайса, ал кейбіреулерінде төмендейді. Сонымен қатар, жарық соңғы өнімнің сапалық кұрамын да өзгерте алады. Жарықта және температурада қосымша түзілетін метаболиттердің кейбір ферменттеріне әсер ететін болу керек.
Жасанды ортада өскен ұлпаларда қосымша қосылыстардың синтезделу карқындылығын арттыру үшін, қолайсыз факторлардың әсерін қолдануға болады, Мысалы, аноксия, жоғары температура, ультракүлгін сәулесі т.б. Стресс факторлармен әсер етіп, косымша заттардың синтезделуінің реттелу механизмі жете зерттелмесе де, тәжірибе жүзінде қолданғанда іп vitro өскен клеткалардан алынатын өнімдердің мөлшерін арттыруға болады. Оіозкогеа йеішісіеа клеткаларын қалыпты жағдайда өсіргенде 22-26 күнде 70-80 мг/л сапогеншілер түзеді. Стресстік фактор /аноксия/ қолданғанда сапониндердің сондай мөлшері жасанды ортада өскен ұлпаларда 15-16 күнде пайда болды. Бүтін өсімдіктерде қосымша метаболизмнің реттелуі туралы іргелі білім болмағандықтан, іп vitro жағдайында қосымша түзілетін заттардың алмасуын реттеу қиын екені белгілі.
Rauvolfia serpentina мутант клеткалары және сапогенин қатарынан диасгенинді беретін Dioskorea deltoidea мутантты клеткалар линиясы фармацевтік өндіріс үшін өте құнды қосылыс диасгенинді мөлшері жағынан түтас өсімдіктің тамырындай шығарған.
5. Клеткалардың өнімді штаммдарын алу. Құрамыңда қосымша заттардың қандай болса да біреуі мол біркелкі клеткалар популяциясын шығару қиын және күрделі міндет, Себебі, клеткалар популяциясын ұзақ уақыт жасанды ортада өсіргенде, генетикалық өзгергіштікке ұшырайды, тіпті басында жалғыз жоғары өнімді клетканы клондау арқылы алынған болса да біраз уақыттан кейін, жеке клеткалар құнды қосылыстар синтездеу кабілетін жоюы да мүмкін. Клеткалардың түріне тән ерекше биосинтездерге қабілетін жоғары деңгейде ұстап тұру үшін, клеткаларды жасанды ортада өсіру жағдайларын үйлесімді жағдайға жеткізумен қатар, тағы баска жашна да күш салу керек, мысалы, генетикалық әрекеттер мен клеткалық сұрыптау жұмыстарын жүргізу керек. іп vitro өскен клеткаларды мутагендермен немесе кері метаболиттермен өңдегенде, биохимиялық мутанттар алыну мүмкіндігі бар. Мутагенезді және клеткалық деңгейдегі сұрыптауды колданып, Ресейдің өсімдіктер физиологиясы институтының ұлпаларды жасанды ортада өсіру және морфогенез лабораториясында кұрамында аритмияға қарсы қолданылатын алкалоид аймалинды 10 есе көп ұстайтын Каи^оі/Іа ьегрепііпа мутантты клеткалары және сапогенин қатарынан диасгенинді беретін Оіозсогеа йеііоійеа мутантты клеткалар линиясы іріктелініп алынған. Диоскорея клеткалары фармаценттік өндіріс үшін өте кұнды қосылыс диасгенинді мөлшері жағынан тұтас өсімдіктің тамырындай шығарған. Осы жұмыстарын әрі қарай жалғастырып, тәжірибелік мутагенезді және клондау әдістерін колданғанда, диоскореяның штаммы бұрынғыдан да өнімді көп беретін мутант клеткалар линияларын берген.
Қоректік ортада өскен клеткаларды метаболизмге зиянды заттармен, яғни антиметаболиттермен өндегенде, мысалы, ароматтық амин қышкылдарымен, клеткалардың көп бөлігі тіршілігін жояды. Осындай жағдайда тіршілігін сақтап қалған аз ғана клеткалар, метаболиттердің артық мөлшерін синтездеуге қабілетті болады. Осы жолмен сәбіз және картоп клеткаларының триптофанды өте көп мөлшерде синтездеуге қабілетті клондары, ал ақ үйеңкі /клеи белый/ клеткаларының фенилаланин мен тирозин өнімі көп беретін клондары алынған.
Іп vitro жағдайында өсірген клеткаларының неганде рентабельді кең масштабтары өндірісті ұйымдастыру үшін, гормондарға прототрофтық /қажет етпейтін/, қоректік орталарға талғамсыз, сонымен катар осмостық және механикалық стресстерге түраклы мутанттар қажет. Қазір клеткаларды жасанды орталарда өсіріп, мутанттар және олардың варианттарын алу кеңінен таралуда. Дегенмен өнімділігі мол штаммдар алудың ең тиімді жолы, клеткалық және гендік инженерия әдістерін қолданып жасанды түрде клеткаларды қолдан кұрастыру, оның болашағы өте зор.
6. Ең жақсы өнімді штаммдарды сұйық қоректік ортада өсіру. Бастапқыда қатты қоректік ортада алынған каллустық клеткалар сұйық қоректік ортаға ауыстырылады. Клеткаларды өсіру тәсілінің өзгеруі, штаммдардың ерекше қасиеттерінің өзгеруіне әкелмеу керек, яғни штаммдар жасанды орталарда өсіру кезінде болатын стресстерге тұракты болуы керек. Әсіресе бұл мәселеге, кішкене колбаларда лабораториялық тәжірибеде өсірген клеткаларды көлемі үлкен ферменттер ауыстырған кезде, ерекше көңіл аударған жөн.
Биомассаға тиімді химиялық бақылау жасау үшін, клеткалардың жағдайын бақылайтын экспресс әдістер қажет.
7. Өнімді және тұрақты штаммдарды өндіріс жағдайында ферментерлардың көлемін біртіндеп үлкейте отырып өсіру.
Егер, осындай жартылай өндіріс жағдайында өсірілетін клеткалар жоғары деңгейдегі өсу жылдамдығын дәне ізделіп отырған затты биосинтездеу қабілетін сақтап, оны жинақтап және қоршаған ортаға көп мөлшерде бөлетін болса, онда бұндай штамм ірі масштабта өндіріс ұйымдастыру үшін жарамды болады. Одан басқа, штаммның сапалығының маңызды көрсеткіштерінің бірі болып генетикалық тұрақтылығы саналады.
8. Клеткалық биомассаны алу және оны бағалаудың техникалық регламентін (іс тәртібін) жасау. Ол үшін аппаратура қажет, ондай аппаратура Жапония т.б. мемлекеттерде бар. Мысалы, Жапонияда ірі масштабта торғайшөптің клеткаларын жасанды ортада өсіріп , одан шиконин өндіру жолға қойылған. 20000 литрлік ферменттерде убихинон - 10 өнімін беретін темекі клеткалары ортаның үзілмейтін ағында 3 ай бойы өсірілген. Убихинон - 10-ның бір күндік өнімділігі 1 л. суспензияға шақканда 5,582 г. болған.
Қазіргі уақытта дүние жүзінің әр түрлі елдерінде экономика жағынан маңызды заттар өндіруге бағытталған биосинтездік өнеркәсібінде елуден астам өсімдіктер түрлерінің клеткалары қолдануда, олардың ішінде жень-шень, рауволъфия, оймак, гүл және дельта тәрізді диоскорея, торғайшөп, итжедек, алка, сасық мендуана, меруерттүл, агава, апиын т.б.
Бақылау сұрақтары
1. Экономика жағынан маңызды заттар алудын клеткалык технологиясы
2. Сұйық қоректік ортада өсетін клеткалар және олардың ерекшеліктері.
3. Клеткаларды қоректік ортада өсіріп қосымша метаболизмде түзілетін заттар алу жұмыстарының этаптары.
4. Қазіргі технологиялар.
5. Келешектегі технологиялар
Дәріс №17,18
Иммобильденген клеткалар, олардың артықшылығы және иммобильдеу тəсілдері
Жоспар:
1. Иммобилизация растительных клеток: необходимость, основные методы
2. Физиологические основы преимущества иммобилизованных растительных клеток перед традиционными способами культивирования
3. Системы культивирования иммобилизованных клеток
Культуры клеток и тканей растений считаются потенциальным источником специфических вторичных метаболитов, к которым относятся такие соединения, как алкалоиды, стероиды, масла и пигменты. Многие из этих веществ все еще получают путем экстракции из растений. Не ко всем видам растений в настоящее время применимы методы микробиологической промышленности. За исключением некоторых видов растений, суспензионные и каллусные культуры клеток синтезируют вторичные метаболиты в меньших количествах, чем целые растения. При этом рост биомассы в ферментере может быть значительным.
Новым подходом, направленным на увеличение выхода вторичных метаболитов, является иммобилизация клеток и тканей растений. Первая удачная попытка зафиксировать целые клетки была осуществлена в 1966 г. Мосбахом. Он зафиксировал клетки лишайника Umbilicaria pustulata в полиакриламидном геле. На следующий год ван Вецель выращивал клетки эмбрионов животных, иммобилизованных на микрошариках ДЭАЭ (диэтиламиноэтил сефадекса, на основе декстрана). После этого клетки были иммобилизованы на разных субстратах. В основном это были клетки микроорганизмов.
Методы иммобилизации клеток делят на 4 категории:
1. Иммобилизация клеток или субклеточных органелл в инертном субстрате. Например, rktnrb Catharanthus roseus, Digitalis lanata в альгинатных, агарозных шариках, в желатине и т.д. Метод предполагает обволакивание клеток одной из различных цементирующих сред – альгинат, агар, коллаген, полиакриламид.
2. Адсорбция клеток на инертном субстрате. Клетки прилипают к заряженным шарикам из альгината, полистирола, полиакриламида. Метод применялся в экспериментах с животными клетками, а также клетками Saccharomyces uvarum, S. cerevisiae, Candida tropicalis, E. coli.
3. Адсорбция клеток на инертном субстрате с помощью биологических макромолекул (таких, как лектин). Применяется редко, есть сведения об экспериментах с различными линиями клеток человека, эритроцитами крови барана, адсорбированными на покрытой белком агарозе.
4. Ковалентное связывание с другим инертным носителем типа КМЦ. Очень редко применяется, известна удачная иммобилизация для Micrococcus luteus. В основном проводились эксперименты по иммобилизации клеток животных и микроорганизмов. В последнее время интерес к иммобилизации клеток растений значительно возрос, это связано с тем, что иммобилизованные клетки имеют определенные преимущества перед каллусными и суспензионными культурами при использовании их для получения вторичных метаболитов.
В литературе имеются многочисленные данные о том, что существует положительная корреляция между накоплением вторичных метаболитов и степенью дифференцировки в культуре клеток. Кроме того, лигнин, например, откладывается в трахеидах и сосудистых элементах ксилемы только после завершения процессов дифференцировки, что было показано в экспериментах как in vivo, так и in vitro. Полученные данные свидетельствуют о том, что дифференциация и накопление вторичных продуктов обмена веществ происходит в конце клеточного цикла. При снижении роста процессы дифференциации ускоряются.
Изучение содержания алкалоидов, накапливаемых многими растениями in vitro, показало, что компактные, медленно растущие культуры клеток содержат алкалоиды в больших количествах, чем рыхлые, быстро растущие культуры. Организация клеток необходима для их нормального метаболизма. Наличие организованности в ткани и ее последующее действие на различные физические и химические градиенты – четкие показатели, по которым различаются высоко- и низкопродуктивные культуры. Очевидно, что иммобилизация клеток обеспечивает условия, приводящие к дифференциации, упорядочивает организацию клеток и способствует тем самым высокому выходу вторичных метаболитов.
Иммобилизованные клетки имеют ряд преимуществ: