

54.Особенности высших растений как результат приспособления к жизни на суше.
Обезвоживание. По целому ряду причин для жизни необходима вода, а воздух – это среда, кот. способствует высыханию. Поэтому наземным растениям нужно было выработать какие-то приспособления для добывания и запасания воды.
Размножение. Нежные половые клетки должны быть защищены, а мужские гаметы (сперматозоиды) могут встретиться с женскими гаметами только в воде.
Опора. В отличие от воды воздух никак не поддерживает растение.
Питание. Для ф/с растениям необходим свет и СО2, поэтому хотя бы часть растения должна возвышаться над землей. А минеральные соли и вода находятся в земле или на ее пов-сти, и, чтобы эффективно использовать эти в-ва, часть растения должна находится в земле и расти в темноте.
Газообмен. Для ф/с и дыхания нужно, чтобы обмен СО2 и О2 происходил не с окружающим раствором, а с атмосферой.
Факторы окружающей среды. Вода, особенно когда ее так много, как, скажем в озере или в океане, обеспечивает большее постоянство условий окружающей среду. Сухопутная же среда обитания в гораздо большей степени подвержена воздействие таких важных переменчивых факторов, как температура, интенсивность освещения, концентрация ионов в среде и рН.
Адаптационные приспособления голосеменных и покрытосеменных растений к жизни на суше.
Основные преимущества этих растений над всеми остальными, конечно же, связано с их способом размножения. Здесь можно выделить три главных аспекта:
1. Гаметофитное поколение очень редуцировано. Гаметофит полностью зависит от спорофита и всегда находится под его защитой. А у мхов и печеночников, у кот. преобладает гаметофит, и у папоротников, у кот. есть свободноживущий заросток, гаметофит не защищен и очень легко высыхает.
2. В отличие от всех др. растений, у кот. сперматозоиды подплывают к я/кл, покрытосеменным для оплодотворения не нужна вода. Мужские гаметы семенных растений неподвижны и переносятся ветром или насекомыми вместе с пыльцевыми зернами. На конечном этапе опыления мужские гаметы проникают к я/кл ч/з пыльцевую трубку, а сами я/кл при этом заключены внутри семязачатков.
3. Из всех современных растений только у семенных есть особые структуры – семена. Возникновение семени стало возможным благодаря тому, что семязачатки вместе со всем своим содержимым остаются на родительском спорофите.
а) У всех сосудистых растений ткани ксилемы и склеренхимы лигнифицированы и дают внутреннюю опору. У многих семенных растений наблюдается вторичный рост и отложение большого кол-ва древесины (вторичной ксилемы). К таким растениям относятся кустарники и деревья.
б) Настоящие корни, кот. тоже характерны для сосудистых растений, позволяют эффективно извлекать влагу из почвы.
в) От высыхания эти растения защищены эпидермисом и нерастворимой в воде кутикулой или же пробкой, образующейся при вторичном утолщении.
г) Эпидермис наземных органов, и в особенности эпидермис листьев, пронизан устьицами, что способствует лучшему газообмену м/у растением и атмосферой. У голосеменных устьица глубоко погружены в ткань листа – сохранение воды. Листья в виде иголок – уменьшение площади пов-сти и потери воды.
+появление цветков вместо шишек у покрытосеменных, что позволило привлекать насекомых, птиц и летучих мышей для опыления.
55. Ксенобиотики в экосистемах
 5
5




57. Биотрансформация ксенобиотиков






58. Биоаккумулирование ксенобиотиков





59. Мембранотропное действие




60. Водообмен растений, его характеристика.
В клетке вода существует как в связанном, так и в свободном состоянии. В клеточной стенке часть воды находится в адсорбированном состоянии, связанном с гемицеллюлозой. В порах есть свободная вода. В цитоплазме есть свободная и коллоидная. Интенсивность осмотических процессов зависит от свободной воды, но есть корреляция между связанной водой и выживаемостью растений.
Вода поступает в клетку по трем механизмам: 1. Осмотический – зависит от разности концентраций веществ по обе стороны клеточной стенки. Система, содержащая полупроницаемые мембраны и растворы разных концентраций называется осмотической. Диффузия воды через мембрану называется осмосом; концентрация растворенных веществ в вакуоли служит мерой максимальной способности клетки поглощать воду. В состоянии насыщения осмотический потенциал одинаков по абсолютной величине тургорному давлению: Р = Т. Таким образом, в клетку может поступать вода только в том случае, если осмотический потенциал превышает тургорное давление. Сила, с которой вода поступает в вакуоль и была названа сосущей: S = Р – Т. Если клетка полностью насыщена водой (тургенесцентна) S = 0, тогда Р = Т (правая сторона рисунка). Когда подача воды уменьшается (ветер, недостаток влажности в почве и т. д.), то сначала возникает водный дефицит в клеточных оболочках, водный потенциал в них уменьшается (становиться ниже, чем в вакуолях) и вода начинает передвигаться в клеточные оболочки. Отток воды из вакуоли снижает тургорное давление, и, таким образом, увеличивается сосущая сила. При длительном недостатке влаги большинство клеток теряет тургор. В этих условиях Т = 0, S = Р 2. Коллоидно-химический механизм (набухание). В клетку вода может поступать и в результате набухания. Набуханием называют поглощение жидкости или пара высокомолекулярным веществом (набухающим телом), сопровождаемое увеличением объема. Состояние набухания протоплазмы имеет решающее значение для интенсивности всего обмена веществ. Вода в протоплазме является важнейшей средой для биохимических реакций и диффузии, а гидратация протоплазматических белков необходима для поддержания ультраструктуры и функциональной активности органоидов. У некоторых частей растения поступление воды происходит исключительно путем набухания, например у семян и у многочисленных приспособлений эпифитов для поглощения воды и водяного пара (всасывающие чешуйки и др.). Вода проникает в набухающее тело путем диффузии. Удерживаемые набухающим телом молекулы воды теряют часть своей кинетической энергии, которая превращается в тепло. 3. Электо-осмотический – вода может поступать в клетку в виде диполей за счет разности потенциалов на мембране.
Транспирация – это физиологический процесс испарения воды растениями. Основным органом транспирации является лист. Следовательно, основную роль в испарении воды играют следующие виды транспирации: устьичная (испарение воды через устьица); кутикулярная (испарение воды с поверхности листа, покрытого кутикулой);перидермальная (через чечевички, стебель, почки).
Говоря о перемещении воды по растению выделяют транспорт в тканях одного органа, который называют ближним (радиальный), и транспорт между отдельными органами, который называют дальним. Между ними есть существенная разница. Ближний транспорт идет по неспециализированным тканям, а для дальнего в растениях имеются специальные проводящие ткани. вода, попавшая в корневой волосок или другую клетку корня с помощью одного из механизмов, о которых мы говорили, движется к сосудам ксилемы, т. е. происходит радиальное перемещение воды по тканям корня .Для поступления воды в корень необходимо чтобы водный потенциал ризодермиса был меньше, чем водный потенциал почвенного раствора; в этом случае вода начнет поступать в клетки. Дальний транспорт воды. Путь, который проходит вода от корневого волоска до испаряющей клетки листа, распадается на две части: различные по протяженности, строению и физиологическим признакам. Первая часть состоит из живых клеток и имеет малые размеры (миллиметры или доли миллиметра). Это два коротких участка – один – в корне, от его поверхности с корневыми волосками до сосудов, которые находятся в его центральном цилиндре; второй – в листе, от сосудов, входящих в состав проводящего пучка, и испаряющей воду в межклетники хлоренхимы. Вторая часть пути – это сосуды, трахеиды, которые представляют собой мертвые трубки. У травянистых растений их длина составляет несколько сантиметров, а у деревьев достигает нескольких метров и даже десятков метров. Силу, которая подымает пасоку вверх по сосудам, назвали корневым давлением. Механизм корневого давления, как считают, основывается на действии сократительных белков, функцию которых выполняют микрофибриллы Ф-белков. Чем интенсивней клетки листьев испаряют воду, тем быстрее она будет поступать в клетки корней и быстрей транспортироваться вверх по растению.
61. Дыхание – разрушение органических веществ при участии кислорода воздуха, в результате образуется СО2 и Н2О. Энергия используется на образование АТФ. То что дыхание проходит в 2 стадии определил Паладин.
1. С6Н12О6 + 6Н2О + 12R → 6 СО2 + 12RH2 , 12 R – дыхательный пигмент, 12 RН2 – дыхательный хромоген.
2. 12RH2 + 6О2 → 12R + 12 Н2О
Итого С6Н12О6 + 6О2 → 6 Н2О + 6СО2
Первая фаза – анаэробная, а вторая – аэробная.
В дыхание участвуют вода и пигменты
Окисление в результате дегидрирования
Часть энергии запасается в связях АТФ, а часть восстанавливает коферменты (доноры Н+). Восстановление коферментов идет с образованием воды и одновременным синтезом АТФ. Энергия должна освобождаться малыми дозами и использоваться эффективно (иначе она рассеется в виде тепла).
Отличительные особенности дыхания растений.
Окисляется экзогенный НАДН (НАДФН), который поступает из цитоплазмы с помощью НАД(Ф)Н дегидрогеназы.
На внутренней мембране наряду с цитохромным переносом электронов по ЭТЦ, функционирует цианидный путь (устойчивый к действию цианидов), т.к. есть устойчивая цианидоксидаза в растительных стенках.
Митохондрии растительных клеток содержат малит-энзим, который катализирует окислительное декарбоксилирование малата с образованием пирувата, далее малат окисляется в цикле Кребса.
Гликолиз или путь Эмбдена – Мейергофа – Парнаса
Этот процесс подразделяют на две стадии: 1) активирование и расщепление глюкозы; 2) окисление, восстановление и образование АТФ и пирувата.
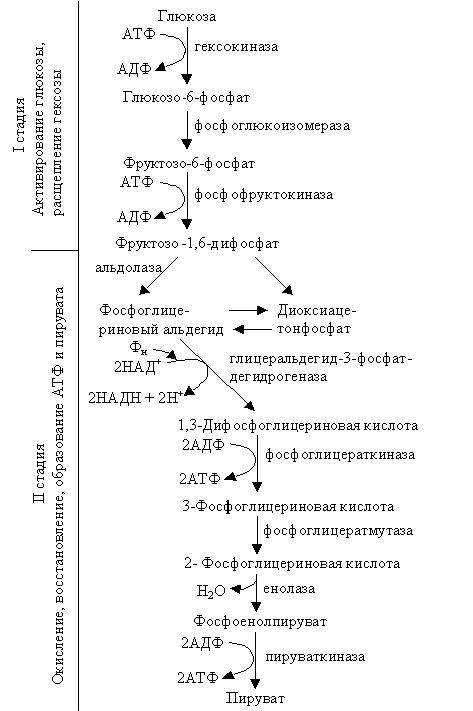
|
Все реакции, происходящие при гликолизе, можно представить в следующем виде: С6Н12О6 + 2НАД+ + 2АДФ + 2Н3РО4 → 2СН3СОСООН + 2НАДН + 2Н+ + 2АТФ
Во время гликолиза – первой фазы дыхания при распаде молекулы глюкозы на две молекулы пирувата (ПВК) образуется 4 молекулы АТФ. Однако 2 молекулы АТФ используются вначале процесса для активации глюкозы, таким образом, в клетке запасаются только 2 молекулы АТФ. Одновременно на этой фазе дыхания при активации ФГА до ФГК образуются две молекулы НАДН. Гликолиз происходит в гиалоплазме и ядре. Для него не нужен О2. Это анаэробная стадия дыхания.
Превращение пирувата. В аэробных условиях главный путь превращения образовавшейся при гликолизе ПВК – окислительное декарбоксилирование при помощи пируватдегидрогеназного мультиферментного комплекса, составной частью которого является коэнзим А (СоА или СоА-SH). В результате образуется ацетил-СоА. Окисление пирувата и образование ацетил-СоА (активированной уксусной кислоты) – очень сложный процесс, рассматриваемый подробно в курсе биохимии. Ограничимся суммарным выражением этого процесса: СН3СОСООН + НАД+ + СоА-SH → СН3СО ~ S-СоА + НАДН + Н+ + СО2
В анаэробных условиях пируват вступает в реакции брожжения. При брожжении, в основном спиртовом, пируват декарбоксилируется с образованием уксусного альдегида при участии карбоксилазы, а затем восстанавливается до этилового спирта ферментом алкагольдегидрогеназой: СН3СОСООН → СН3СОН + СО2
СН3СОН + НАДН + Н+ → СН3СН2ОН + НАД+.