

1.Суть и значение цтк.
В аэробных условиях образовавшийся ацетил-СоА вступает в цикл Кребса. В цикле Кребса после реакций отнятия и присоединения воды, декарбоксилирования и дегидрирования ацетильный остаток, поступивший в цикл в виде ацетил-СоА, полностью расщепляется. Суммарная реакция записывается в следующем виде: СН3СО ~ S-СоА + 3Н2О + АДФ + Н3РО4 → НS-СоА + 2СО2 + 4[Н2] + АТФ. Цикл Кребса проходит одинаково у животных и растений. Первая реакция цикла – перенос ацетильного остатка от ацетил-СоА на щавелево-уксусную кислоту (ЩУК) с образованием лимонной кислоты (цитрат).
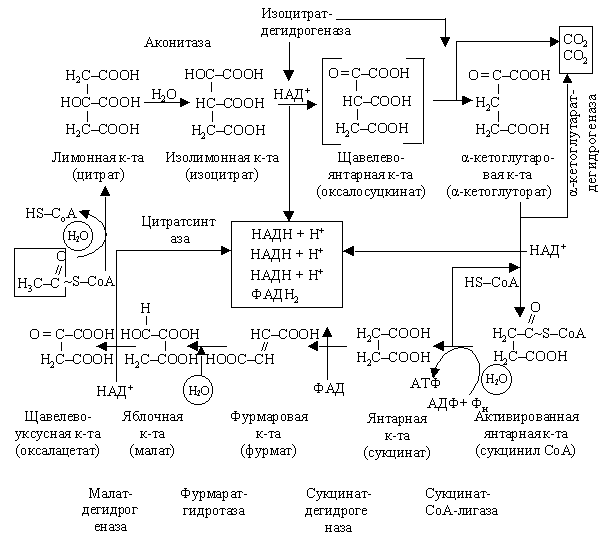
В ходе реакции, катализируемой цитратсинтазой, растрачивается макроэргическая связь ацетил-СоА, т. е. та энергия, какая была занесена в процессе окисления пирувата перед началом цикла. Это значит, как и гликолиз, цикл Кребса начинается не с запасания энергии в клетке, а с расходования. Цепь преобразований, образующих этот цикл и направленных, в конечном счете, на разрушение углеродного состава ряда кислот, начинается с их увеличения: двухуглеродный фрагмент (уксусная кислота) присоединяется к четырехугольному фрагменту ЩУК с образованием шестиуглеродной трикарбоновой кислоты цитрата, которая может запасаться в клетках в больших количествах.
Таким образом, цикл Кребса – процесс каталитический и начинается не с катаболизма (разрушения), а с синтеза цитрата. Цитратсинтетаза, катализирующая эту реакцию, относится к регуляторным ферментам: она ингибируется НАДН и АТФ. НАДН – конечный продукт, в форме которого запасается энергия, освобождаемая в процессе дыхания. Чем активней цитратсинтетаза, тем быстрей пойдут и другие реакции цикла, быстрей пойдет дегидрирование веществ с образованием НАДН. Однако увеличение количества последнего вызывает ингибирование фермента, и цикл затормозится. Это пример реакции по принципу обратной связи. Следующая серия реакций – преобразование цитрата в активную изолимонную кислоту (изоцитрат). Она протекает при участии воды и по сути сводится до внутримолекулярного преобразования лимонной кислоты. Промежуточным продуктом этого преобразования является цис-аконитовая кислота:

Катализируются обе реакции аконитатгидротазой. Затем изоцитрат дегидрируется с участием изоцитратдегидрогеназы, коферментом которой является НАД+. В результате окисления образуется щавелево-янтарная кислота (оксалосукцинат). Последняя кислота декарбоксилируется. Отсоединяющийся СО2 принадлежит ацетильному остатку, вступившему в цикл в виде ацетил-СоА. В результате декарбоксилирования образуется очень активная α-кетоглутаровая кислота (кетоглутарат).
α-Кетоглутарат, в свою очередь, подвергается тем же изменением, которые происходят перед началом цикла с пируватом: одновременное окисление и декарбоксилирование.
В реакции принимает участие α-кетоглутарат дегидрогеназный комплекс.
Освободившийся СО2 является другой частицей, которая отщепляется от ацетильного остатка. Образовавшийся в результате этих сложных преобразованием янтарная кислота (сукцинат) вновь дегидрируется, и образуется фумаровая кислота (фумарат). Реакция происходит с помощью сукцинатдегидрогеназы. Фумарат после присоединения молекулы воды легко преобразуется в яблочную кислоту (малат). В реакции принимает участие фумаратгидротаза. Яблочная кислота, окисляясь, преобразуется в ЩУК при участии НАД+ – специфической малатдегидрогеназы. Напомним, что ЩУК конечный продукт цикла Кребса – образуется и при фотосинтезе С4-растений (цикл Хетча – Слека) при карбоксилировании ФЕП на свету, и в темноте у растений типа САМ. Таким образом, цикл Кребса заканчивается и может начинаться сначала. Одно условие – подача новых молекул ацетил-СоА. Главное значение цикл Кребса состоит в запасании энергии, которая освобождается в результате разрушения пирувата, в макроэргических связях АТФ. Поставляя в клетку АТФ, цикл Кребса может являться регулятором других процессов, идущих с затратой энергии, таких как транспорт воды и солей, синтез и транспорт органических веществ. Чем быстрей проходит преобразование веществ в цикле, тем больше может синтезироваться АТФ, тем быстрей пойдут указанные процессы.
Промежуточные вещества, образуемые в цикле, могут использоваться на синтез белков, жиров, углеводов. Например, ацетил-СоА – необходимый продукт для синтеза жирных кислот, кетоглутарат может в результате восстановительного аминирования преобразовываться в глутаминовую, а фумарат или ЩУК – в аспарагиновую кислоты.
Суммарный результат цикла Кребса сводится, таким образом, к тому, что каждая ацетильная группа (двухуглеродный фрагмент), которая образуется из пирувата (трехуглеродный фрагмент) расщепляется до СО2. Во время этого процесса восстанавливается НАД+, ФАД+ и синтезируется АТФ.
В регуляции цикла ди- и трикарбоновых кислот важное значение имеет соотношение между НАДН и НАД+, а также концентрация АТФ. Высокое содержание АТФ и НАДН угнетает активность таких ферментов цикла Кребса, как пируватдегидрогеназа, цитратсинтетаза, изоцитратдегидрогеназа, малатдегидрогеназа. Повышение концентрации оксалоацетата угнетает ферменты, активность которых связана с его синтезом, – сукцинатдегидрогеназы и малатдегидрогеназы. Окисление 2-оксиглутаровой кислоты ускоряется аденилатами, а сукцината – АТФ, АДФ и убихиноном. В цикле Кребса имеется и ряд других пунктов регулирования.