

А) класифікація за характером приквітників.В суцвіттях квітки не роз’єднані справжніми листками, а можуть мати лише приквітники. За їх характером суцвіття поділяються на два типи:
– якщо приквітники у вигляді зелених пластинок (фіалка триколірна, купена, вербозілля), суцвіття називається фрондозним:
– у вигляді лускоподібних листочків (брактеї) (конвалія, грушанка) – брактеозне.
Між цими двома типами є перехідні форми: фрондульозні (зірчатка, дзвоник кропиволистий). Якщо немає приквітників зовсім – голі, або ебрактеозні (дика редька).
б) класифікація за характером поведінки апікальних меристем. Перехід пагона до цвітіння супроводжується прискореним ростом і розвитком. При цьому верхівкова меристема пагона змінює свою форму та розчеплюється, формуючи зачатки квітів. У багатьох рослин суцвіття виникають всередині бруньок як єдине ціле.
Характер поведінки апікальних меристем є одна із морфологічних ознак, за якими класифікують суцвіття. У одних рослин верхівкова меристема повністю витрачається на формування верхівкових квітів. Такі суцвіття називаються визначеними, іноді закритими верхньоквітковими, обмеженими (дзвоник розлогий, гадючник оголений). В обмежених суцвіть верхівкова квітка випереджає у розвитку бокові і називається суцвіття верхньоквіткове.
У інших рослин ці меристеми завжди залишаються у вегетативному стані і називаються необмежені, невизначені. Є свої різновиди.
В одних випадках у цих невизначених суцвіттях верхівкова меристема продовжує активну діяльність протягом вегетаційного періоду (хрестоцвіті), у інших вона завмирає, перетворюючись на невеличкий додаток (конвалія, гіацинт). І, накінець, випадок, коли від апікальної меристеми залишається невеличкий горбочок, а суцвіття закінчується верхньою пазушною квіткою. Вона може зміститися на верхівку, але за природою ця квітка є бічною. Її називають псевдотермінальною.
Відрізнити від справжніх верхівкових квіток її можна, спостерігаючи за послідовністю розпускання квітів. Справжні верхівкові квіти випереджають у розвитку нижні бічні, а псевдоверхівкові розпускаються пізніше бічних. Це є акро- і базипетальний порядок наростання. У невизначених суцвіттях наростання, таким чином, акропетальний порядок.
в) тип галуження :
– Моноподіальний, коли на кінчику пагона апікальна меристема зберігається і входить до складу верхівкової зимової бруньки. Ця меристема із року в рік нарощує пагін, який є результатом діяльності верхівкової меристеми. Так і бічні, але ріст їх обмежений.
– Симподіальний. Брунька, що формується на кінці пагона за своїм походженням є бічною, а верхівкова – відмирає. Таким чином, наростання іде у підлеглому порядку. Це є пагін, який складений осями підлеглих порядків і являє собою результат діяльності бічних меристем.
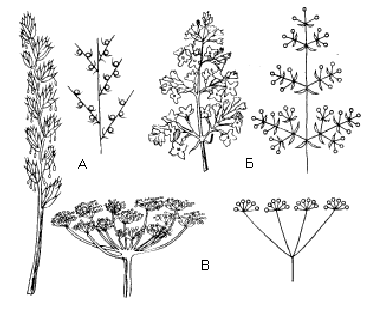
Мал. Моноподіальні суцвіття та їх схеми:
А – складний колос (пирій),
Б – волоть (бузок),
В – складний зонтик (кріп).
За способом галуження поділяються на групи: моноподіальні – ботричні, симподіальні – цимозні.
Ботричні (рацемозні) суцвіття характеризуються моноподіальним типом галуження (назва). Головна вісь закінчується не квіткою, а конусом наростання, тому суцвіття продовжує рости невизначено довго. Для цієї групи суцвіть характерне неоднакове розпускання квітів. Спочатку дозрівають нижні, потім – верхні. Такий порядок розпускання квітів називається акропетальний.
г) класифікація суцвіть за ступінню галуження. Поділяються на прості і складні. У простих ступінь галуження не більше 2, тобто на бічних осях ( ІІ порядку) сидять квітки. Прості ботричні суцвіття:
Простий колос. На стрижні розміщуються сидячі квітки (подорожник).
Китиця. На основі стрижня прикріплені окремі квіти, які сидять на однакової довжини ніжках (конвалія).
3)проста сережка – це, власне, пухкий колос, має ніжний стрижень, направлений донизу ( тополя, верба, береза).
Простий зонтик. Квітки сидять на довгих квітконіжках, які виходять майже із одного місця на верхівці пагона. Довжина квітконіжок однакова (вишня, цибуля).
Щиток. Подібне до китиці, але нижні квітконіжки довгі за верхні і квітки розміщуються в одній площині (горобина, груша, калина, глід)
Головка має вкорочену і потовщену вісь, до якої з усіх сторін прикріплені сидячі квіти (конюшина).
6) Кошик. Має дуже розширену і плоску основну вісь (дещо вигнуту), на якій розміщуються сидячі квіти розміщуються сидячі квіти.
М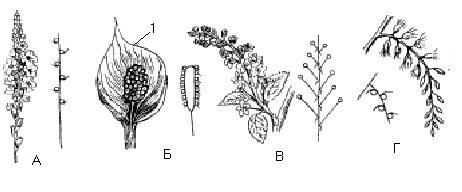 ал.
Прості суцвіття з видовженою віссю
та їх схеми:
ал.
Прості суцвіття з видовженою віссю
та їх схеми:
А – колос – вербена,
Б – початок – кремена,
В – китиця – черемха,
Г – сережка – тополя;
1 – покривало.

Мал. Прості суцвіття з укороченою віссю та їх схеми:
А – зонтик – цибуля,
Б – головка – конюшина,
В – кошик – календула;
1 – квітколоже,
2 – обгортка
Складні ботричні суцвіття
1) Складний колос. Головний стрижень не прямий, а у вигляді сходинок і на кожній сходинці сидить не один, а групка квітів у вигляді простого суцвіття – колоска (пшениця, жито, ячмінь).
2) Складний зонтик. На квітконіжках не окремі квіти, а прості суцвіття. Мають листкову обгортку (кріп, петрушка).
3) Волоть. Центральний стрижень галузиться і на кінцях сидять прості колоски (овес, сорго), або поодинокі квіти (бузок).
Цимозні, симподіальні, визначені.
Мають симподіальне галуження. Головна і бічні гілки закінчуються квіткою. Після утворення квітки на пагоні ріст припиняється і послідуючі гілки переростають попередню. Верхня квітка розпускається раніше, а нижні – пізніше – базипетальне наростання.
а) Монохазій (звивина і закрутка).
У звивини розвивається тільки одна бічна вісь, яка переростає головну і закінчується квіткою. Бокові гілки відходять то вправо, то вліво (гладіолус, ірис, гравілат).
Закрутка подібна, тільки бічні вісі відходять на одну сторону (незабудка).
б) Дихазій (розвилка). Під квіткою головної осі утворюються 2 супротивно розміщені гілки, вони теж закінчуються квіткою і т.п. (гвоздика).
в) Плейохазій. Псевдозонтик.
Нижче квітки головної осі утворюється декілька бокових, переростаючи головну вісь і розміщується мутовчасто (молочай, гречка).
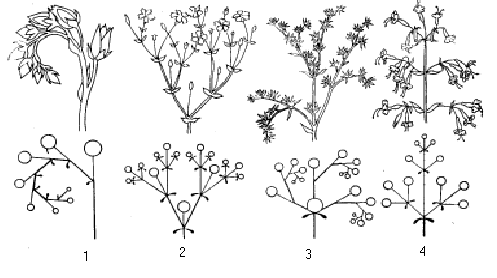
Мал. Симподіальні суцвіття та їх схеми:
1 – монохазій – закрутка – ехеверія,
2 – дихазій – роговик;
3 – плейохазій – очиток,
4 – тирс – смілка.
Отже, до морфологічних ознак суцвіття належать: характер приквітників, ступінь галуження (прості, складні), спосіб галуження (ботричні, цимозні), послідовність розпускання квітів (акропетальна, безопетальна), поведінка апікальних меристем (коли апікальна меристема витрачається на формування верхівкових квітів – визначені суцвіття (дзвоник розлогий). У інших меристема залишається у вегетативному стані (губоцвіті); продовжує діяльність (хрестоцвіті).
Вказані чотири групи морфологічних ознак не залежать одне від одного у більшості випадків, але деякі чітко взаємопов’язані. Ботричні і невизначені суцвіття завжди моноподіальні, цимозні завжди визначені.
Еволюція суцвіть йшла по направленню збільшення кількості квітів в суцвітті, зменшення їх розмірів і об’єднання їх у компактні групи, які краще помітні для комах-запилювачів.
3. Якщо розглядати всю надземну частину рослини як суцільну пагонову систему, то і всі суцвіття, які розміщуються на ній теж можна розглядати як єдине суцвіття, як квітконосні зони загонових систем, що розвиваються із бруньок поновлення. Це структурна єдність має назву синфлоресценція. Кількість, розмір і розміщення суцвіть залежить від моделі пагоноутворення даного виду рослини. Це генетично спадкова ознака.
Однорічники формують одну синфлоресценцію, може охоплювати значну частину пагонової системи, а якщо квітки і пагони збагачення розвиваються на всіх вузлах та сім’ядольний вузол, тоді вся рослина є суцільною синфлоресценцією.
У багаторічних трав, які наростають симподіально (це можуть бути напівкущі), синфлоресценцію охоплюють верхні ділянки пагонів збагачення. І їх кількість буде залежити від кількості бруньок, які пішли у ріст в цьому році. Наприклад, у слабенько кущистих кореневищних ( купена) – 1 синфлоресценція, у трав і напівкущів із багатьма бруньками (полин гіркий, волошка, свербіжник).
У моноподіальних трав’янистих багаторічників формуються різні
Якщо генеративні пагони розвиваються із бруньок, що перезимували, то кожен такий пагін є окрема синфлоресценція. Їх ділення (гравілат).
Якщо генеративні пагони розвиваються із бруньок. Що не мали стану спокою (подорожник), виникає одна синфлоресценція. Головна її вісь захоплює ділянку вегетативного пагона, встроюючись у багаторічну скелетну основу, тобто інтеркалярно, вставним ростом (вероніка дубравна, луговий чай).
Подібні вставні синфлоресценції властиві повзучим моноподіальним довгопагоновим травам. Синфлоресценцію у них охоплюють якусь невелику відносно частину вегетативного пагона і чергуються із вегетативними зонами:
Між цими варіантами є перехідні форми, що розвиваються як із бруньок, які перезимували, так із бруньок, що не мали періоду спокою. У подібних випадках нижні і верхні генеративні пагони морфологічно дуже відрізняються. Перші добре розвинені, другі – редуковані. В цих перехідних варіантах вся пагонова система – суцільна синфлоресценція. Приклад, фіалка прекрасна.
Основа будь-якої синфлоресценції утворює пагін І порядку, на якому з’являються пагони збагачення, що в свою чергу галузяться. У межах синфлоресценції порядок галуження дуже різниться від 2 до 20-80.
У випадках дуже сильного галуження спостерігається так звана ієрархія суцвіть як сукупність взаємопов’язаних і взаємопідпорядкованих пагонових систем.
Розглянемо на прикладі буркуна лікарського.
В нижній частині пагона поновлення заключені бруньки, які дадуть наростання надземних органів. Це зона поновлення. Вище – зона гальмування, тобто, нерозгалужена ділянка стебла, де пазушні бруньки недорозвинені. Вище бічні бруньки формують бічні пагони збагачення.
Головний пагін закінчується головним суцвіттям, яке відокремлено від зони збагачення основним міжвузлям. Воно у буркуна досить коротеньке, що свідчить про зміну швидкості і ритму морфогенезу пагона при переході до цвітіння.
Головне суцвіття у буркуна – складна китиця (волоть подвійна). Пазушні китиці (із пазух трійчастих листків) – це приватні, парціальні, елементарні суцвіття.
У своїй будові бічні пагони збагачення ніби повторюють будову головного. Їх називають паракладіями. Вони галузяться. Утворюють пагони І порядку, ІІ, ІІІ і т.д.
Таким чином, вегетативна ділянка пагона поновлення представлена зонами гальмування і поновлення, а генеративна частина – складна ієрархічна синфлоресценція у вигляді складної волоті.

Мал. Суцвіття буркуна лікарського:
1 –фрондозна складна китиця головного суцвіття;
2 – суцільне суцвіття (синфлоресценція);
а – головне суцвіття;
б – суцвіття пара кладіїв;
в – ебрактеозні прості китиці.
4. Типологічна класифікація синфлоресценцій базується на ознаці – тривалість існування апікальних меристем головної осі та паракладіїв. Якщо ці меристеми формують верхівкові (термінальні) квіти, синфлоресценції називають монотелічними, монотелями.
Якщо верхівкова меристема за у вегетативному стані, синфлоресценції називають політелічними, політелями.
Тей чи інший тип синфлоресценцій характеризує крупні таксономічні одиниці: родини, підродини.
Так, рослини родин жовтецевих, гвоздичних, геранієвих мають монотелічні синфлоресценції, а родини хрестоцвітих, губоцвітих, бобових, складноцвітих, ранникових – політелічні.
Основні морфологічні відмінності монотеліїв і політеліїв:
У монотеліїв синфлоресценцій всі вісі завершуються квіткою, у політеліїв – верхівки пагонів І, ІІ, ІІІ порядків залишаються вегетативними.
Політелії на верхній частині мають головне суцвіття, у монотеліїв воно відсутнє.
Тільки політелії мають основні міжвузля, у монотеліїв – це квітконіжка останньої термінальної квітки.
У політеліїв синфлоресценцій головна вісь несе як елементарні суцвіття, так і пара кладії (або загальні); у монотеліїв – всі бічні гілки, що відходять від головної осі – є паракладії, а елементарні суцвіття – відсутні.
5. Екологічна класифікація синфлоресценцій базується на пристосуваннях до успішного запилення. Так, антодіями називають будь-яке суцвіття, яке зовні нагадує поодиноку квітку (всі складноцвіті; рослини, які мають щиткоподібну волоть, крайові квітки у суцвіть збільшені – калина, молочай).
Сережка – будь-яке суцвіття із пониклою віссю і дрібними квітками. Серед сережок зустрічаються як прості суцвіття-китиці (тополя, верба), так і складні.
У ряді випадків виникають складні суцвіття, в яких об’єднуються риси ботричних і цимозних суцвіть. Якщо елементарні цимозії суцвіть зібрані на осі, які мають моноподіальний характер, то говорять про тирс, тирсоїдне суцвіття.
Сережки берези, вільхи, граба, горіха ліщини, дуба, каштана – це тирсоїдні суцвіття у вигляді сережки. Не всі тирсоїдні суцвіття є сережками, наприклад суцвіття живокісту, синяка. Сережками вважаються тільки ті, що після цвітіння опадають.
Запилення – це перенесення пилку з тичинок на приймочку маточки. Відбувається в період цвітіння, коли пильники відкриваються, а приймочки функціонують як органи, що сприймають пилок. Найдавнішим, скоріше всього, є випадок самозапилення у рослин із двостатевими квітками.
Запилення пилком однієї квітки приймочки іншої- перехресне запилення. Розрізняють два типи: гейтоногамію ( від гр. „гейтон” – сусід) , коли пилок однієї квітки запилює приймочку другої, але тієї ж рослини; ксеногамію („ксенос”- чужий), коли запилюється приймочка іншої рослини.
Біологічні переваги перехресного запилення – аутбридінгу - зрозумілі. Це, перш за все, рекомбінація генетичного матеріалу, що сприяє внутрішньовидовому розмаїттю. Але в житті рослин інбридинг важливий як засіб розмноження і розселення генетично однорідних популяцій. Тому в житті перехреснозапильних в багатьох випадках можливе і самозапилення.
Якщо взяти деякі види рослин, які мешкають у різних кліматичних умовах
( притаманні і північним і помірним зонам ),спостерігається така закономірність: тундрові представники утворюють плоди виключно внаслідок самозапилення, а для „ лісовиків” характерне перехресне запилення.
Самозапилювачами є дуже багато видів культурних рослин та бур’янів: більшість видів пшениці, ячменю, вівсу, проса, гороху, бобів, льону, бавовнику. Чому? Адже зернові є вітрозапильними рослинами, бобові – комахозапильні.
Історія становлення цих видів в умовах вирощування їх в культурі заглиблюється на тисячоліття. Вірогідно, багатовіковий несвідомий відбір корисних ознак для людини закріплював процес самозапилення. А бур’яни як супутники культурних рослин теж перейшли до самозапилення.
Що ж сприяє самозапиленню? Які пристосування повинні бути, щоб попередити, з одного боку, навіть випадкове потрапляння „ чужорідного „ пилку на приймочку, а, з другого, забезпечити потрапляння дозрілого пилку на власну, теж готову приймочку?
Самий надійний шлях – зовсім не розкрити квітки при повному дозріванні власного андро- та гінецею. Це і є клейстогамія ( від гр. „клейстос” – закритий) – запилення у закритій квітці. Вона може бути повною, коли квітка втратила механізм відкривання. Прикладом можуть бути деякі види фіалок та кислиць. У них поряд із нормальними, хазмогамними („хазма” – тріщина, зів), утворюються закриті квітки, які розміщуються біля поверхні ґрунту. У кислиці звичайної (тайга) клейстогамні квіти дрібні (3 мм) з редукованими пелюстками та короткими стовпчиками. Пильники не розкриваються, а пилок проростає всередині їх і трубки проходять через стінку пиляка по направленню до приймочки. Є і хазмогамні квіти, але в них не може відбуватися самозапилення через те, що приймочки на 1/3 вище за пиляки. А от комахи сприяють самозапиленню.
В разі, якщо відбудеться випадкове перехресне, насіння утвориться. Чому ж клейстогамія переважає у видів північних зон? Це є пристосуванням до умов затіненої тайги. Сильне затінення та підвищена волога викликає збільшення клейстогамних квітів і зменшення хазмогамних.
Фіалка польова- вид високогір’я Анд. Дуже красиві і складно збудовані весняні квіти залишаються безплідними через відсутність потрібних запилювачів. Зате чисельне насіння у багатьох видів фіалки утворюють дрібні та невиразні, схожі на бутони квіти. Вони утворюються пізніше на прямостоячих стебельцях або на повзучих пагонах, іноді на кореневій порості, занурюючись у ґрунт.
Клейстогамія може бути і неповною, тобто квітка відкривається після запилення. На рослинах бальзаміну поряд із яскраво забарвленими хазмогамними квітками з’являються і напіврозкриті із недорозвиненими пелюстками , а також повні клейстогамні. Більше їх утворюється, якщо рослина виросла у незвичайних умовах: на піску, щебні, торф’яниках. Але, якщо насіння з цих квітів висіяти у добрий лісовий ґрунт, розвиваються рослини із жовтими розкритими квітками.
Механізмами самозапилення є: контактна автогамія внаслідок дотику пиляків до власної приймочки (веснівка, одинарник, під’ялинник); внаслідок руху квітконіжки (грушанка); внаслідок проникнення квітки в кінці цвітіння (дзвоники, цикорій, соняшник).
Алогамія – перехресне запилення. Гейтоногамію не можна вважати „чистою„ алогамією, тому що рекомбінації генетичного матеріалу не відбувається- це один і той же організм.
В рослинному світі є багато пристосувань, що попереджають автогамію. Адже в природі великий процент рослин має двостатеві квітки і теоретично власний пилок може запилювати власну приймочку.
До морфологічних пристосувань належать механічна перешкода попадання пилку на приймочку, так звана гетеростилія : одні екземпляри мають квітки із довгими стовпчиками, інші – з короткими, іноді ще стовпчики проміжної форми.
Класичним прикладом є диморфна гетеростилія ( диморфізм ) у первоцвіту весняного. Явище перехресного запилення у нього було детально вивчене Ч. Дарвіном у 1862 і 1877 рр. Він спостерігав, що коли джміль в пошуках нектару занурює свій хоботок у трубку довго стовпчикової квітки, пило на його хоботку опиняється точно на такому рівні, на якому знаходиться приймочка у коротко стовпчикової квітки.
Якщо комаха відвідає саме таку квітку, то пилок потрапить на її приймочку. Таким же чином відбувається запилення від коротко стовпчикової квітки до довго стовпчикової форми. Але, як помітив Дарвін, комахи можуть переносити пилок і до однокової форми квіток і навіть часто сприяти самозапиленню. Таким чином, у гетеро стильних первоцвітів можливі три варіанти запилення.
Шляхом ретельних експериментів із різними гетеростильними первоцвітами Дарвін встановив, що найбільш успішним для рослини є перехресне запилення між різними формами, тому що в цьому випадку утворюється більш життєздатне насіння і його значно більше, ніж у разі інших варіантів запилення. Перший був названий Дарвіном легітимним, два інших –іллегітимним ( від лат. „legitimus” –законний; „illegitimus” – незаконний).
Більш рідкісне явище- триморфізм, яке властиве кисличним та плакунам
(плакун верболистий). Запилення у таких рослин буде ефективним тільки у тих випадках, коли на приймочку кожної форми потрапить пилок від двох інших форм.
Цікавий спосіб пристосування до перехресного запилення і одночасно запобіганню самозапилення у рослини монохорії списовидної. Цій рослині також властивий диморфізм квітів, але він супроводжується не гетеростилією. Диморфізм обумовлений відгином великої фертильної тичинки у праву або ліву сторону і відгином стовпчика, відповідно, у протилежному напрямку. Квітки монохорії мають 6 тичинок. З них 5 - жовті, маленькі, стерильні ; 1- голуба, велика, з виростами- шпорцями, фертильна. Відгин тичинки відувається завжди у сторону, де знаходиться шорець ( якщо їх два,то до більшого ). Ці два типи квіток – тичинка відігнута вправо, приймочка – вліво – знаходяться у одному суцвітті. Але вікриваються вони неодночасно. Якщо комаха-бджола відвідала квітку, то верхівка пиляка та приймочка торкаються різних частин тіла комахи. І при відвідуванні такої ж форми квітки запилення не відбудеться, але відбудеться, якщо відвідає іншу форму. До речі у цих рослин після цвітіння суцвіття занурюється у воду і подальше дозрівання насіння відбувається у воді.
Запобіганням самозапиленню є спосіб розкривання пиляків. Наприклад, у квітках складноцвітих пилок потрапляє спочатку всередину тичинкової трубки, а потім складеними лопатями приймочки, зокрема їх несприймальною частиною до пилку, виноситься назовні, коли приймочка розгорнеться, пилок буде під нею, звідки і буде збиратися комахами.
Зрозуміло, що і роздільностатевість та дводомність виключають автогамію.
В природі є ще один спосіб, який виключає автогамію – це неодночасне дозрівання пиляків та приймочок в двостатевих квітках – дихогамія. В одних випадках квітка пилить, коли в ній ще не дозріла приймочка - явище протерандрії, протандрії, в інших – приймочка готова до сприйняття, коли ще пиляки закриті – явище протерогінії, протогінії.
Протандричні квіти більшості бобових, злаків, осок, зонтичних гвоздичних. Протогінічні – у ранникових, деяких злакових, у гречки, гладіолуса.
У багатьох рослин, що мають двостатеві квітки, зустрічається часткова функціональна роздільностатевість. При цьому більшість квітів є двостатевими, друга частина квітів на цій же рослині, або на іншій – функціонально одностатеві через недорозвиненість андро- ,або гінецею.
Варіанти: чоловіча однодомність, коли на одній рослині є двостатеві і чоловічі квітки – андромоноеція ; жіноча однодомність, коли на одній особині є двостатеві та жіночі – гінемононодіеція; чоловіча дводомність, коли двостатеві квітки та чоловічі знаходяться на різних екземплярах – андродіеція; жіноча дводомність, коли двостатеві та жіночі квіти знаходяться на різних особинах – гінедіеція.
4. Загальні риси вітрозапильних рослин: дрібні розміри квітів з недорозвиненими оцвітинами; тичинки та приймочки далеко виходять за межі покриву квітки; сприймаюча поверхня приймочки сильно збільшена; квітки зібрані у суцвіття за межами листкової маси.
Як пристосування до рознесення пилку вітром може бути вибухоподібне швидке розкривання квітів. Так, у кропиви дводомної у чоловічих квітках тичинкові нитки згорнуті у щільну спіральку і при розгортанні із силою викидають хмаринку пилку. Іноді пилок висипається у човникоподібні при- квітники і поступово видувається звідти вітром ( обліпиха ).
Досить складне вітрозапилення у ліщини та злаків.
Ліщина, лісовий горішник – одна із самих ранньоквітучих рослин нашої флори. Квітки роздільностатеві, а рослина однодомна. Чоловічі суцвіття – сережки, зُявляються в середині літа як пазушні пагони на приростах цього року; в кінці літа вони достигають 0,2-0,5 см довжини, мають квітки із зачатками тичинок, до часу листопаду у них вже є сформовані пилкові зерна. Чоловіча квітка має 8 тичинок і основою приростає до напівзрослих лусочок (криючий листок та приквітник). Вони разом утворюють щось на вигляд даху (козирок), змикаються із іншими такими ж лусочками сережки. Це захист від зимового висушування „голих” квіток. Групки жіночих квіток (дихарії) знаходяться у пазухах листкових зачатків зимуючих бруньок, які розкриваються навесні. Восени жіночі квітки ще не сформовані.
У лютому починається вітягування чоловічих сережок, в кінці березня – початок квітня лусочки розходяться і пилок розноситься вітром. В цей час із бруньок, що містять жіночі квітки (їх зачатки), висуваються червоні приймочки, але зав’язі ще немає; дозрівання насіннєвих зачатків відбувається лише через 2–2,5 місяця після запилення. Таким чином, запилення відбувається навесні при низьких температурах, а запліднення і формування плодів відкладається на теплий літній час.
Комахами запилюються, мабуть, 70% всіх покритонасінних. Квіти їх надзвичайно різноманітні за формою, забарвленням, будовою. Екологічним фактором цього є перш за все поведінка запилювача.
Комахи відвідують квіти заради їжі (пилок, нектар), у пошуках пари, для виведення потомства, притулку. Всі ці фактори приваблення до квітки запилювача називається первинними атратантами.
Вторинні атратанти – це сигнали про наявність перших. Це і специфічна форма, запах, забарвлення.
Одна із самих оригінальних рослин світу – пасіфлора (російська назва-страстоцвет) з великими незвичної будови квітами. Малюнки її були привезені в Європу із Нового Світу у 1610 році Джакомо Боччі.
Всередині яскраво забарвленої оцвітини виділяється ще більш яскрава корона з довгих прямих або хвилеподібних ниток. В центрі квітки піднята на гінофорі піднімається зав’язь з трьома хрестоподібно складеними приймочками. Навколо 5 тичинок із продовгуватими пиляками.
Іспанські місіонери, що прийшли за Колумбом, побачили в окремих частинах квітки „знаряддя пристрастей господніх”: корона – терновий вінок; тичинки та приймочка – знаряддя катувань.
За розкішність цю рослину назвали „кавалерською зіркою”. Стилізовані квіти пасіфлори стали основою для чудової чавунної огорожі Михайлівського замку у Петербурзі.
Запилюються комахами та колібрі. Мініатюрні пташки, не сідаючи не квітку, в польоті занурюють свої дзьобики в глибину квітколожа і дістають нектар і маленьких комах, що теж їм ласують. Вони (комахи) потрапляють у лопушку – внутрішню корону із шипами, що закриває вихід.
Перехресному запиленню сприяє узгоджені рухи пиляків та приймочок. Як тільки квітка розпуститься, вертикально розміщені пиляки лопаються, поздовжньо на внутрішній стороні. Потім вони перевертаються, спочатку на 90о, стаючи горизонтально, а потім навколо осі – і звисають вниз, вже відкритою стороною назовні. Дозрілий пилок потрапляє на спинку птаха. Як тільки пиляки звільняться від пилку – дозріває приймочка (протандрія). Вона починає відгинатися донизу і під кінець розміщується в одній площині із пиляками. Таким чином пилок із тільця птаха потрапляє на дозрівши приймочку. Так і комахи – опушені джмелі.
Одним із чудових пристосувань є здатність пасіфлор до самозапилення в тому випадку, якщо не відбулося перехресне. Пасіфлора – узгоджені рухи назустріч одни одному пиляків та приймочок.
Є серед пасіфлор і справжні хіроптерофіти, чиї квітки пристосувалися до запилення летючими мишами.
Дуже своєрідний і цікавий механізм запилення у юки. Квітки відкриваються вночі і запилюються маленьким білим метеликом з роду Пронуба. Самки збирають пилок у грудочку і переносять її у другу квітку. Метелик вчавлює пилок у приймочку, а потім відкладає яйця у зав’язь цієї квітки. Личинки ж потім живляться насінням. Їх так багато, що вистачає і личинкам і для розмноження рослини.