

4. Статевий процес.
Статеве розмноження характерне для всіх еукаріотів (ядерних). Відсутність у деяких – це вторинне явище, вони позбулися статевого процесу за певних умов. У прокаріотів його ніколи не було, що пояснюється відсутністю ядра.
Статеве розмноження є найдосконалішим, тому що має ряд переваг:
При статевому розмноженні утворюється більша кількість зачатків нових особин;
Набагато більша можливість розселення на значні відстані, а це означає освоєння більших територій;
Поява нового, як правило, більш досконалого матеріалу для природного добору.
Статеве розмноження забезпечує поєднання більш менш різних батьківських і материнських задатків, потомство стає більш різноманітним завдяки їх комбінаціям.
Це означає, що нове покоління має більшу можливість пристосовуватись до постійно змінюю чого зовнішнього середовища, а отже і виживання.
Статевий процес – це злиття двох статевих клітин–гамет і утворення зиготи.
Обов’язковим при утворенні статевого процесу є злиття не тільки цитоплазми, а й обов’язковим є злиття ядер. Утворюється диплоїдний набір хромосом (від гр. „diploos” – подвійний ); ядра ж гамет мають вдвічі менший набір – гаплоїдний (від гр. „haploos” – одинарний, простий).
Розглянемо типи статевого процесу за мірою їх ускладнення і удосконалення.
Найпростіші типи цього процесу будуть відповідать більш примітивним рослинним організмам. Типи будуть залежати від типів гамет:
1) Зовні копулюючі гамети (копуляція - злиття) не відрізняються між собою. Прикладом може бути вищезгадана водорость дунаніела солоноводна , синьо – зелена. Називається процес хологамія.
2) Кон’югація. (нитчаста водорость спірогіра). Відділ рослин, яким при таманна кон’югація і називаються Зчіплянки або Кон’югати.
Нитки водорості складаються з гаплоїдних клітин. Восени, на двох поряд розміщених нитках спірогіри утворюють копуляційні вирости, які сполучаються між собою і вміст клітин через калюс зливається. Вміст однієї клітини (вважається чоловічої) переливається до другої (жіночої). Отже, кон’югація це злиття вегетативних клітин, але гаплоїдних.
Еволюція рослин передбачила і ускладнення гамет, утворенню яких передує поділ клітин (мейоз). Крім того, вони спеціалізуються: змінюються їх розмір і з’являється певних у певних рухомість. Більш простий серед них процес ізогамії.
За формою і розміром жіночі і чоловічі однакові: їх умовно називають „плюс” і „мінус”-гаметами. Такий процес притаманний хламідомонаді, улотриксу. При цьому зливаються гамети різних особин, це явище називатьєся гетероталізм. Коли відбувається злиття гамет, різних за розміром – це гетерогамний процес.
Оогамія. Процес, при якому іде злиття більшої за розміром жіночої гамети, яйцеклітини із маленькою рухомою, озброєною джгутиками чоловічою гаметою, або сперматозоїдом. Безджгутикові чоловічі гамети називаються спермії. Вони в процесі еволюції втратили джгутики (переважна більшість голонасінних і всі покритонасінні). Отже, оогамія – еволюційно найвищий тип статевого розмноження.
Переваги:
велика, або більша за розміром яйцеклітина має запас поживних речовин, який забезпечує живленням зиготу, майбутній зародок;
в свою чергу, зменшення розмірів чоловічих гамет забезпечило їм рухомість навіть у маленькій кількості вологи;
нерухомість яйцеклітини обумовлює внутрішнє запліднення і надійний захист зиготи – майбутній зародок.
У вищих рослин гамети формуються у спеціальних статевих органах – гаметангіях. У примітивних рослин гамети утворюються в вегетативних клітинах шляхом поділу.
У разі оогамії, тобто досконалого статевого процесу, чоловічі статеві органи і жіночі морфологічно відрізняються. Гаметангії, що утворюють яйцеклітину називаються оогонії , чоловічі гамети - антеридії . У вищих рослин багатоклітинні жіночі статеві органи називаються архегонії, чоловічі – антеридії. Еволюція йшла за напрямком редукції – спрощення у зв’язку із пристосуванням до наземного способу життя. Зовсім втрачені ці органи у покритонасінних.
Поняття про цикл відтворення.
Коли в рослинному організмі іде формування диплоїдних спор, він називається спорофіт. Наприклад доросла рослина папоротей із спор проростає організм, на якому формуються гамети – гаметофіт. У різних рослинних організмів в цей час іде формування спорофіту і гаметофіту. Якщо узагальнити, то можна сказати, що з кожним еволюційним кроком іде скорочення гаметофіту і збільшення у масі і об’єму спорофіту.
Зрозуміло, що перехід до гаметофіту, повинен супроводжуватись редукційним поділом, який відбувається у певний період. У більшості водоростей мейотично поділяється зигота. У зв’язку із цим все їхнє життя відбувається у гаплоїдному стані. Це рослини-гаплобіонти. Отже, у більшості спорофіт відповідає диплобіонту, а гаметофіт – гаплобіонту . Рослини із гаплоїдною кількістю хромосом у вегетативному стані називається гаплобіонтами .Рослини із диплоїдною кількістю хромосом у вегетативному стані – диплобіонти.
У процесі еволюції спостерігається редукція (спрощення) одного із поколінь, або спорофіта, або гаметофіта. Вона обумовлена зміною зовнішніх умов.
Гаплобіонтами є деякі водорості, зокрема улотрикс. Приклад напрямку розвитку, коли у циклі розвитку рослини домінує гаметофіт (Зозулин льон, мох).
Гаметофіт – є доросла рослина. Він дводольний, тобто на одній формується чоловічій статевий орган – антеридій , на других – архегонії. На верхівках чоловічих рослин формуються округлі шишечки у основі буруватих розеток листків. Це сперматогенні клітини, в яких утворюються дводжутикові сперматозоїди.
На верхівках жіночих в оточенні зелених листочків формуються архегонії. Архегоній має колбовидну форму з розширенним черевцем, яке містить яйцеклітину, а над нею черевну канальцеву клітину і звужену довгу шийку з рядом канальцевих клітин. Ці допоміжні клітини ослизнюються і у цій рідині просувається сперматозоїд.
Із зиготи розвивається спорогоній (від гр. spora – посів, honos - народження) – спорофіт.
Будова. Коробочка на ніжці, яку прикриває ковпачок. Клітини – диплоїдні. Із спори утворюється нитчаста протонема. Це зумовлює думку про походження мохів від нитчастих водоростей. На ній закладається кілька бруньок, з яких і розвивається гаметофіт.
Отже, під циклом відтворення розуміють певний відрізок життя виду, що обмежений двома одноклітинними станами від спорофіту до спорофіту, від гаметофіту до гаметофіту. У багатьох представників рослинного світу спостерігається чергування поколінь у межах одного циклу.
Чергування поколінь у рівноспорових папоротей. (чоловіча папороть, або щитник).
Спорофітом є сама рослина з кореневищем, коренем і листками-вайями. Ядра клітин відповідно мають подвійний набір хромосом.
З нижнього боку листків закладаються спорангії у вигляді скупчень – сорусів. Зовнішні його клітини утворюють покривальце, внутрішні – археспоріальну тканину, з якої в результаті мітотичних поділів утворюють тетради спор.
Розкриття спорангія і вивільнення спор відбувається за допомогою фіброзного шару, в якого клітини нерівномірно потовщені. При зневодненні і підсушуванні фіброзне кільце як пружинка розкриває. Спори, висипавшись, проростають у заростки. На них утворюються і антеридії і архегонії. Має вигляд пластинки серцеподібної (2 – 3 см діаметром).
В антеридіях формуються сперматозоїди із джгутиками. В архегоніях (колоподібної форми) –яйцеклітина. Обов’язковим для злиття є наявність води. З прониклих сперматозоїдів тільки один зливається з яйцеклітиною. Запліднена яйцеклітина перетворюється у зиготу. Яка і дає початок дорослому спорофіту.
Отже у папороті є не тільки чергування ядерних фаз, а й два різних за виглядом і функціями самостійне покоління: спорофіт і гаметофіт. Таку зміну поколінь називають гетероморфною.
У папороті чоловічі заростки – гаметофіти двостатеві, однакові за формою і фізіологічно. Серед вищих спорових рослин спостерігається утворення двох видів спор – , мікро- і макроспори і відповідно два типи зародків – чоловічі і жіночі. Приклад – сальвінія плаваюча – однорічна рослина. 5-15 см. завдовжки, живе на поверхні води.
Голонасінні .
Продовження. Різноспорові папороті.
На одній рослині, але в різних спорангіях, які називаються спорокарпії – гомологи сорусів рівноспорових, утворюються мікро- і макроспорангії.
В мікроспорангіях в результаті мейотичного поділу утворюються мікроспори, які тут же проростають у дуже редуковані чоловічі заростки. В антеридіях (2) розвивається по 4 багатоджгутикових сперматозоїда.
Мікроспорангії великі, розвивається 1 макроспора. Вона після проростання дає багатоклітинний жіночий заросток трикутної форми, в якому розвиваються архегонії. Закладка іде лише в одн.
Отже, в порівнянні із рівноспоровими:
різні за розміром спорангії і спори;
спори не висипаються, а проростають всередині спорангія;
утворення дводомних гаметофітів і редукція одного з них (чол.);
відносно вкорочений час життя гаметофіти.
У голонасінних життєвий цикл у загальному плані протікає за типом різноспорових, але має свої специфічні особливості. Перш за все вони виражені у будові насінного зачатка (пригадайте, що у папоротей зачатком розмноження була мегаспора (в мегаспорангії). У голонасінних насінний зачаток вже є видозмінений у процесі еволюції мегаспорангій. Друга особливість – утворення з насіннєвого зачатка не спори, а насіння. Воно утворюється на материнській рослині і проростає в подібну до материнської рослини.
Отже, немає морфологічно відокремлених поколінь: гамето- і спорофіту, процес цей, як і чергування ядерних фаз замаскований. Жіноча шишка невелика, червоно-бурого розвивається на верхівці пагона.
Насінний зачаток у голонасінних утворюється на жіночих шишках – мегаспорофілах. Разом із насіниною сидить відкрито, звідси і назва від. Складається насіннєвий зачаток із нуцелуса (власне мегаспор) і покривів (інтегументів). Клітини нуцелусу, є диплоїдними. Одна із його клітин стає материнською клітиною (археспоріальною). Вона редукційно ділиться і тільки одна з 4х проростає у первинний ендосперм, який гомологічний вегетативній частині жіночого гаметофіта у різноспорових (трикутна зелена пластинка).
У верхній частині первинного ендосперму розвивається 2 редуковані архегонія ( жіночий статевий орган). Отже із диплоїдної клітини нуцелусу, утворилися 4 гаплоїдних, з яких тільки одна мегаспора дозріває, а інші ідуть на її живлення.
В середині нуцелуса ця мегаспора проростає і створює багатоклітинний жіночий зародок, або гаметофіт, який знаходиться під надійним захистом. В ньому накопичується велика кількість поживних речовин – первинний ендосперм – він же і є жіночий гаметофіт.
Коли ж відбудеться запліднення. Перше – коли потрапить пилок, але за того пройдуть певні процеси підготовки в самому жіночому гаметофіті.
Вони полягають у тому, що на верхній частині ендосперму біля мікропіле утворюються два архегонії із яйцеклітинами і декілька канальцевих клітин. Чоловічі шишки, або стробіли виникають на тій же рослині (біля основи молодих пагонів). Це вісь, вкрита лусочками. Слід сказати, що у голонасінних редукований ще більш.
Мікроспорангії (їх 2 ) – називається пилковий мішок. Проростає в середині мікроспорангія. Вона одноядерна має покрив: екзину і інтину. Після поділу утворюються дві клітини, антеридіальна і спороногенна.
У різних голонасінних запліднення забезпечується дещо по-різному.
У гінкгових, саговникових – за допомогою сперматозоїдів, а пилкова трубка, що формується із сифоногенної клітини виконує роль гаусторії, за допомогою якої чоловічій заросток прикріплюється до нуцелуса. У інших голонасінних, наприклад у хвойних, чоловічій гаметофіт після розкриття мікроспорангія висипається в повітря. Двоклітинний пилок потрапляє на жіночі шишки. Відбувається запилення.
Отже, у зв’язку із переходом до надземного способу життя з’явилось пристосування і перевага – незалежність від водного середовища. Замість води як засобу пересування гамет виникло запилення - перенесення пилку різними агентами (повітряними течіями, комахами, а пилкова трубка – засіб внутрішнього пересування чоловічої гамети до яйцеклітини).
Після запилення жіночі шишки щільно закривається, зеленіють, потім лусочки дерев’яніють. Лише наступної весни пилок (який і є практично чоловічим гаметофітом) проростає. Пригадайте, що пилок на стадії двох клітин висипався: антеридіальної і сифоногенної.
Із антеридіальної під час проростання утворюються дві клітини: одна стерильна (гине) і друга – спермогенна. Одночасно внутрішня оболонка пилку інтина витягується у пилкову трубку за участю сифоногенної клітини. Туди потрапляє спермогенна клітина, вона ділиться з утворенням двох чоловічих гамет – сперміїв. Через пилкову трубку один запліднює яйцеклітину, другий – гине. Бачимо, що запліднення – внутрішній процес. Це ще одна перевага насінних над споровими.
З розвитком зародка нуцелус зовсім руйнується і перетворюється на плівочку. Зародок вкривається покривами (інтегументами) нуцелусу. Ця оболонка має має крильце подібні придатки для рознесення вітром. Отже утворюється насінина.
В насінині: зародок формується з зиготи, живиться за рахунок первинного ендосперму (жіночий гаметофіт) і має насіннєву шкірочку із покривів нуцелуса. Насінина є органом розмноження, розселення і відтворення. Це переваги насінини перед спорами.
Життєвий цикл у покритонасінних має специфічні особливості в порівнянні вже із голонасінними. По-перше, у них з’являється особливий орган насінного розмноження – квітка, головний орган якої маточка. Маточка утворилася внаслідок зростання мегаспорофілів (плодолистиків). Жіночий гаметофіт ще більше редукований і представлений зародковим мішком. Насінний зачаток і насінина розвиваються швидше, ніж у голонасінних (вегетаційний період).
Будова насінного зачатка, який формується за рахунок клітин зав’язі маточки, дещо подібний до голонасінних. Головну масу складає нуцелус, або мегаспорангій і покрив. У нуцелусі формується зародковий мішок – він же і є мегаспора. Покриви інтегументи є.
Розрізняють 5 типів насінних зачатків:
1) ортотропний, або прямий. Мікропіле знаходиться на одній осі із рубчиком ( місцем прикріплення насінного зачатку до ніжки) та ніжкою (фунікулюсом). Ніжка дуже коротенька (гречка, ароїдні);
2) антропний. Обернений на 180º. Мікропіле та рубчик знаходяться на одній лінії;
3) гемітропний. Нуцелус напівобернений на 90 º по відношенню до плаценти та ніжки;
4) кампілотропний. Спостерігається однобічне розростання нуцелуса та інтегументів (бобові);
5) амфітропний. Двостороннє ввігнутий нуцелус у вигляді підкови.
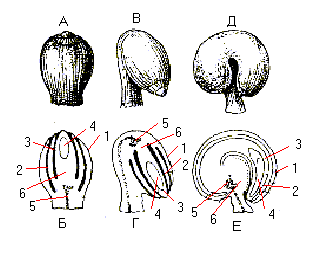
Мал. Різни типи насінних зачатків (схема):
А, Б – прямий (атропний);
В, Г – обернений (анатропний);
Д, Е – зігнутий (кампілотропний);
А, В, Д – зовнішній вигляд;
Б, Г, Е – насінні зачатки в розрізі:
1 – зовнішній інтегумент;
2 – внутрішній інтегумент;
3 – нуцелус;
4 – зародковий мішок;
5 – провідний пучок;
6 – халаза.
Формування зародкового мішка починається із проростання мегаспори з утворенням 4 гаплоїдних клітин. Одна дає початок зародковому мішку, а три– відмирає ( ідуть на підживлення) до або після запилення.
Ця проросла мегаспора, або одне гаплоїдне ядро, ділиться на дві половини. Ядра розходяться до полюсів: одне - в сторону мікропілярного полюсу, друге - в сторону халатного полюсу. На полюсах поділяється двічі і утворюється по 4 ядерні структури. З кожного ядра до екватора наближується по одному ядру. Зливаючись, вони утворюють вторинне ядро зародкового мішку. Із тих, що залишились на мікрополярному полюсі (по 3 на кожному), формується яйцеклітина і синегіди (яйцевий апарат). На другому полюсі формуються антиподи. Їх роль не з’ясована.
Таким чином, сформований і готовий до запліднення зародковий мішок складається з яйцеклітини, синергід, антиподів і вторинного ядра. Зародковий мішок 8-ядерний.
Ми власне розглянули мегаспоро – і мегагаметогенез, або розвиток зародкового мішка.
Мікроспорогенез і мікрогаметогенез.
Мікроспорогенез – це утворення мікроспор у пилкових гніздах. Мікргаметогенез – проростання мікроспор і утворення чоловічого гаметофіту – пилинки.
Формування відбувається в сукупності тичинок андроцеї (мікроспоролистках) з утворенням пилинок. В процесі утворення пилку відбуваються досить складні процеси, які можна назвати підготовчим періодом.
Спочатку у пиляка можна виділити два шари клітин: епідермальний – зовнішній і субепідермальний – внутрішній. Саме в субепідермальному шарі закладаються мікроспорангії. На місці майбутніх пилкових гнізд закладається по одній збільшеній клітині. Вона ділиться перегородкою на дві: із зовнішньої утворюється ще три шари клітин:
зовнішній або фіброзний (лат. fibra – волокно) ще називається ендотецій (від гр. „endon” – в середині, „there” - скринька).
Характеризуються його клітини тим, що стінки їх нерівномірно потовщені завдяки здерев’янінню. Коли пиляк дозріває, стінки клітин цього шару підсихають нерівномірно і це приводить до розривання. Так пилок висівається. Другий шар клітин безпосередньо під ендотецієм виконує підживлюючу роль. Він руйнується і вмістом живиться пилок. Третій шар формується до середини пиляку іназивається вистиляючий, тапетум (гр.” тапетум”– килим). Тапетум прикриває внутрішню тканину пиляка – археоспорій, з якого утворюються мікроспори.
Після мейотичного поділу з археспоріальної клітини формується тетрада гаплоїдних клітин – мікроспори. Дозріла мікроспора одноядерна і має, як у голонасінних, дві оболонки – екзину і інтину. Перенесення пилку залежить від екзини: вітрозапильні мають гладеньку і сухут екзину; комахозапильні – з горбочками, різними виростами; клейку. Форма і розмір пилкового зерна це важлива видова ознака. Кількість може бути надзвичайна, у однієї рослини до 14 – 16 млн (кукурудза).
Запліднення.
Отже, є сформований до запліднення зародковий мішок (яйцеклітина, подвійне ядро, антиподи, синергіди).
Пилок (а він у покритонасінних одноклітинний) мітотично поділяючись, утворює дві клітини: маленьку репродуктивну і велику – вегетативну, або сифоногенну.
Пилинка потрапила на приймочку маточки (пригадайте, що у голонасінних при запиленні вона потрапляє безпосередньо до насінного зачатка – характерна ознака голонасінних). Вегетативна клітина рухається до зав’язі, паралельно утворюючи пилкову трубку. Напрям росту трубки зумовлений законами позитивного хемотаксису, її ріст іде у напрямку подразника, яким є яйцеклітина з насінним зачатком. В трубку потрапляє і генеративна клітина, яка поділяється на два спермії. Так як вони не мають джгутиків, то трубка і забезпечує їхнє проходження до зародкового мішка. Пройшовши успішно через клітини нуцелуса, попередньо розчинивши їх ферментами і дійшовши до яйцеклітини, пилкова трубка розчиняється. Один із сперміїв зливається із яйцеклітиною і утворюється зигота; другий зливається із вторинним ядром це - майбутній ендосперм. Синергіди і антиподи зникають.
Ендосперм формується у різних покритонасінних по – різному. У одних ядро багаторазово поділяється. Ядра розміщуються по периферії зародкового мішка. Потім біля кожного формується цитоплазма і з’являються клітинні оболонки. Сформовані клітини зразу ж поділяються і виповнюють весь зародковий мішок Такий тип формування ендосперму називається нуклеарний або ядерний. Якщо зразу ж після поділу триплоїдного ядра утворюються клітинні оболонки, такий тип називається клітинний, або целюлярний. Є рослини, у яких зовсім не утворюється ендосперму - орхідні .
Явище подвійного запліднення, відкрите у 1898 р. рос. біологом Навашиним.
Формування зародку у покритонасінних. Зигота поділяється і утворює дві клітини. Одна (до напрямку мікропіле) багаторазово поділяється і утворює підвісок, який занурюється в і живить майбутній зародок. Друга, нижня клітина утворює кулясте тіло (тричі поділившись). Верхня його частина з часом стає плоскою. У дводольних на протилежних кінцях площини, з горбочків формуються сім’ядолі. Однодольні мають одну, видозмінену сім’ядолю - щиток. У злакових біля щитка є виріст – епібласт – рудимент другої сім’ядолі.
Покритонасінні з’явилися в кінці юрського періоду. Вони дуже швидко витіснили інші відділи і зайняли пануюче положення в рослинному світі нашої планети.
Виникає питання: „Що ж спонукало таке явище?” Різні вчені пов’язують його з різними факторами. Мабуть, кожен з цих факторів і відіграв певну роль.
-покритонасінні краще всіх пристосувалися до умов середовища, які склалися на той час. Зокрема, зменшення вологості, збільшення сонячного освітлення.
-виникненню і подальшому розвитку покритонасінних сприяла більша захищеність насінного зачатка, якій заключається у зав’язі та плоді. Крім того, плід сприяв ефективно му розповсюдженню.
- виникнення Покритонасінних співпадає із періодом масової появи класу Комах, які і забезпечували процес запилення.
Існує декілька гіпотез походження квітки: стробілярна, псевдантова і теломна.
Квітка, як особливий орган насіннєвого розмноження, виникла як результат всіх етапів розвитку рослинного світу.
Вихідна форма Покритонасінних - стробіли бенетитових вимерлих голонасінних Мезозойської ери.
За стробілярною гіпотезою квітка походить від двостатевої шишки (стробіли). На осі пагона, дещо вкороченого формуються мікроспорофіли, вище на ніжках макроспорофіли, а між ними – безплідні лусочки. Є ще і покривні лусочки. Вісь стробіла у верхній частині розширена і нагадує сучасне квітколоже.
Сьогодні квітки із спіральним розташуванням її членів, як у ефедри, магнолієвих, нагадують описане вище утворення у вимерлих голонасінних мезозойської ери.
Про те, що квітка сформувалася за рахунок пагона, частково свідчить явище, коли вісь її подовжується і утворюється звичайний пагін. Крім того, анатомічна будова деяких частин квітки теж підтверджують цю гіпотезу.
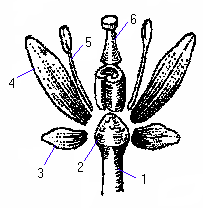
Мал. Схема квітки покрито насінної рослини:
1 – квітконіжка,
2 – квітколоже,
3 – чашолистик.
4 – пелюстка,
5 – тичинка,
6 – маточка.
Наприклад, наявність продихів.
Квітка – видозмінений вкорочений пагін з обмеженим ростом, пристосований до виконування функції розмноження.
Будова квітки: головні частини – маточка , тичинки, всі інші є допоміжними.
Бувають квітки голі, безпокривні (у верби, ясена, образків).
Квітколоже буває плоске (півонія), конічне (суниця), ввігнуте (яблуня, груша).
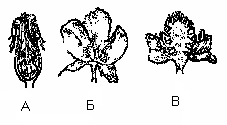
Мал. Форми квітколожа:
А – ввігнуте (шипшина),
Б – плоске (півонія),
В – опукле (жовтець).
Покрив, або оцвітина- це сукупність чашечки та віночка. Оцвітина буває простою та подвійною.
Квітки бувають: двостатеві, різностатеві і стерильні(дзвоникові, розові, жовтцеві).
Стерильні квітки зустрічаються у айстрових. Наприклад, у кошику соняшника, ромашки основну масу займають трубчасті фертильні квітки. Крайові язичкові, подібні до пелюсток –стерильні і виконують функцію приваблення.
Частини квітки розташовані по-різному. В одних випадках вони розташовані по спіралі, наприклад у адонісу. Такі квітки називають ациклічними, або спіральними. В інших оцвітина закладається колами, а тичинки і маточка (маточки) по спіралі (у калюжниці, купальниці, магнолії та ін.). Такі квітки називають гем циклічними, або напівколовими.
Чашечка складається із чашолистиків – нижня частина квітки. Функція – захист, приваблення, поширення плодів, навіть фотосинтезуюча роль. Віночок складається з пелюсток. Основна роль - приваблення запилювачів.
У разі подвійної оцвітини чашечка та віночок добре морфологічно відокремлені ( квітки яблуні мають зеленого кольору чашечку та білі пелюстки).
У разі простої оцвітини або чашечка, або віночок відсутні. Відповідно, проста оцвітина буває чашечкоподібна та віночковидна.
Ще Чарльз Дарвін виявив роль красоти у природі, показав істинні причини походження різноманітного забарвлення квітів. Адже ті рослини, які запилюються вітром не мають яскраво забарвленого віночка.
Як правило, квітки розміщуються на загальному зеленому фоні і забарвлення їх різко контрастує з ним.
Кожний вид комах-запилювачів віддає перевагу певному забарвленню. Так, бджіл приваблюють сині та фіолетові кольори. Жовтих вони майже не помічають, до зеленого – байдужі, а червоні їх відлякують. Але бджоли і джмелі іноді сідають і на пурпурні квіти. Кроваво-червоні приваблюють метеликів. Нічні метелики охоче відвідують білі та блідо-жовті.
Можуть приваблювати і чашолистики. Таке призначення і суцвіть. Але ж, коли квітка маленька її важко примітити, а зібрані у суцвіття вони є приманкою досить яскравою.
Навіть різкі контрасти кольорів у однієї квітки – це пристосування до приваблення. Приклад, королиця (білі крайові, жовті – у центрі),
Те ж можна сказати і про співвідношення кольорів. Так яскраво забарвлена квітка отримує суттєву перевагу у конкуренції з менш помітними квітками і тому із більшою вірогідністю запилюється і дає сильне потомство.
Все розмаїття форм і розміщення пелюсток квітки із правильними і неправильними формами, з пелюстками, розміщеними симетрично і асиметрично, з роздільними пелюстками і тими, що зрослися: двогубими, метеликовими, язичковими – все це більш ускладнені пристосування до перехресного запилення.
Як відомо, різні комахи не рівноцінні як запилювачі. Чи багато буде користі від жуків, які тільки з’їдають пилок? І яка буде користь від мухи, метелика, що кружляють безладно від квітки до квітки? Якщо пилок буде перенесений з огірка на яблуню, або з пшениці на півонію, запліднення не відбудеться!
Ось чому у квіток протягом еволюції виробились спеціальні пристосування, що, наприклад закривають солодкий приваблюючий нектар від некорисних комах і, навпаки, така морфологія квітки, щоби привабити тих комах, які забезпечують перехресне запилення даного виду рослини.
Нижня сторона віночка наперстянки густо покрита волосками. Дрібні комахи в них просто заплутуються, зате великі (джмелі) легко заповзають по них, як по драбинці, щоб дістатися нектару.
У багатьох квітів нектар глибоко захований у шорцях, трубках, що є виростами пелюсток, щоби мухи і інша дрібнота не поласувала їм.
Квітки польової гвоздики мають пелюстки у вигляді зручної посадочної площадки для метеликів, а нектар у них прихований вглибині віночка, дістати його може тільки довгий хоботок цих же метеликів. Коли вони будуть перелітати з квітки на квітку гвоздики, буде відбуватись запилення.
Є така рослина – льонок польовий. Він має віночок зросло пелюстковий віночок і деякі пелюстки зрослися у вигляді губи, яка щільно зімкнена. Квітки льонка опилюють тільки джмелі, вони добувають нектар з глибини шпорки (теж утвір віночка), легко розсуваючи губу віночка.
Продовжуючи тему приваблення слід зазначити,що не тільки колір, нектар, а й різноманітні запахи – це приваблення запилювачів.
Сама велика квітка в світі належить Рафлезії Арнольді (острів Суматра), її діаметр - один метр. Квітка має запах гниючого м’яса для приваблення мух та жуків.Запах виробляється спеціальними залозистими утвореннями.
Виходячи з того, що пристосування для перехресного запилення, що стосується різноманітності форм, формувалися у процесі філогенезу, встановлено, що зрослопелюстковість в еволюційному відношенні більш молода ознака, вторинне утворення. Стародавні (наприклад, магнолієві) рослини мають роздільнопелюсткові квіти.
Явище махровості – занадто збільшена кількість пелюсток, з біологічної точки зору є відхиленням. Походження різне: наприклад, у гвоздики – це розчеплені тичинки, у фуксії розчеплюються самі пелюстки.
Взагалі, вважається, що пелюстки походять від стерильних тичинок, які втратили здатність до утворення пилку.