

Хромосоми (грец. chroma - колір + soma –
тіло) – нуклеопротеїдні комплекси,
матеріальні носії генетичної інформації,
що формуються внаслідок ущільнення
хроматину
(термін і опис – В. Вальдейер, 1888)Хроматин і хромосоми
Хроматин – хімічна основа хромосоми,
ниткоподібні структури ядра, утворені
НК та білками
(термін – В. Флемінг, 1880)
перебігу процесів транскрипції,
репарації і реплікації ДНК;
рівномірного розподілу подвоєного
набору ДНК між дочірніми клітинами
під час мітозу.Забезпечення правильності компактної упаковки молекул днк є необхідною умовою для:
У ядрі кожної соматичної клітини людини:
46 молекул ДНК;
загальна кількість пар основ 6,6109
(біля 2 м);
ДНК упаковується і під час метафази
ступінь її компактизації досягає 104
разів.


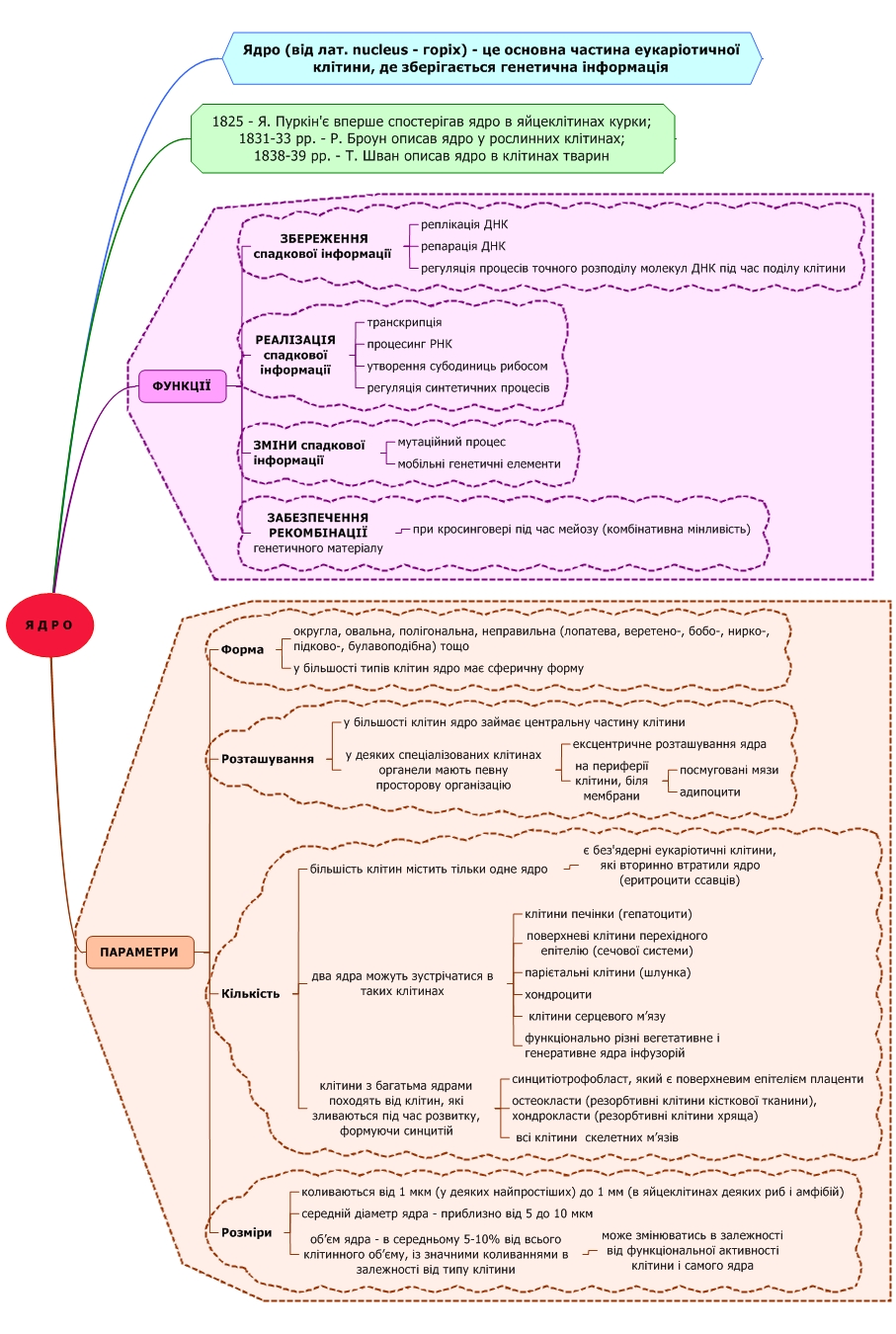
Хроматин – це нуклеопротеїни ядра, що є основою хромосом.
З а
своїми морфофункціональними особливостями
хроматин інтерфазних ядер поділяють
на 2 фракції:
а
своїми морфофункціональними особливостями
хроматин інтерфазних ядер поділяють
на 2 фракції:
1) еухроматин – деспіралізований (світлий), це функціональна частина геному, що складається з генів, відповідальних за життєдіяльність і розвиток організмів.
2) гетерохроматин – спіралізований (темний), транскрипційно неактивний). Види:
- конститутивний гетерохроматин – завжди залишається в конденсованому стані;
- факультативний гетерохроматин – має здатність переходити у стан еухроматину.
Відмінність еухроматину та факультативного гетерохроматину полягає у тому, що перший містить постійно здатну до транскрипції ДНК, а ДНК другого транскрибується лише у певний перод і у визначених клітинах. Цим зумовлюється його склад і відносна кількість у різних клітинах різних організмів, клітин різних тканин та при різному рівні диференціації клітин одного типу.
Гетерохроматин має особливе значення у життєдіяльності клітин зародкового шляху – його гени потрібні для дозрівання гамет та здійснення мейозу.
Хромосоми, відповідно, можуть знаходитися у двох станах:
- спіралізованому (мітотичні хромосоми);
- деспіралізованому (інтерфазні хромосоми)
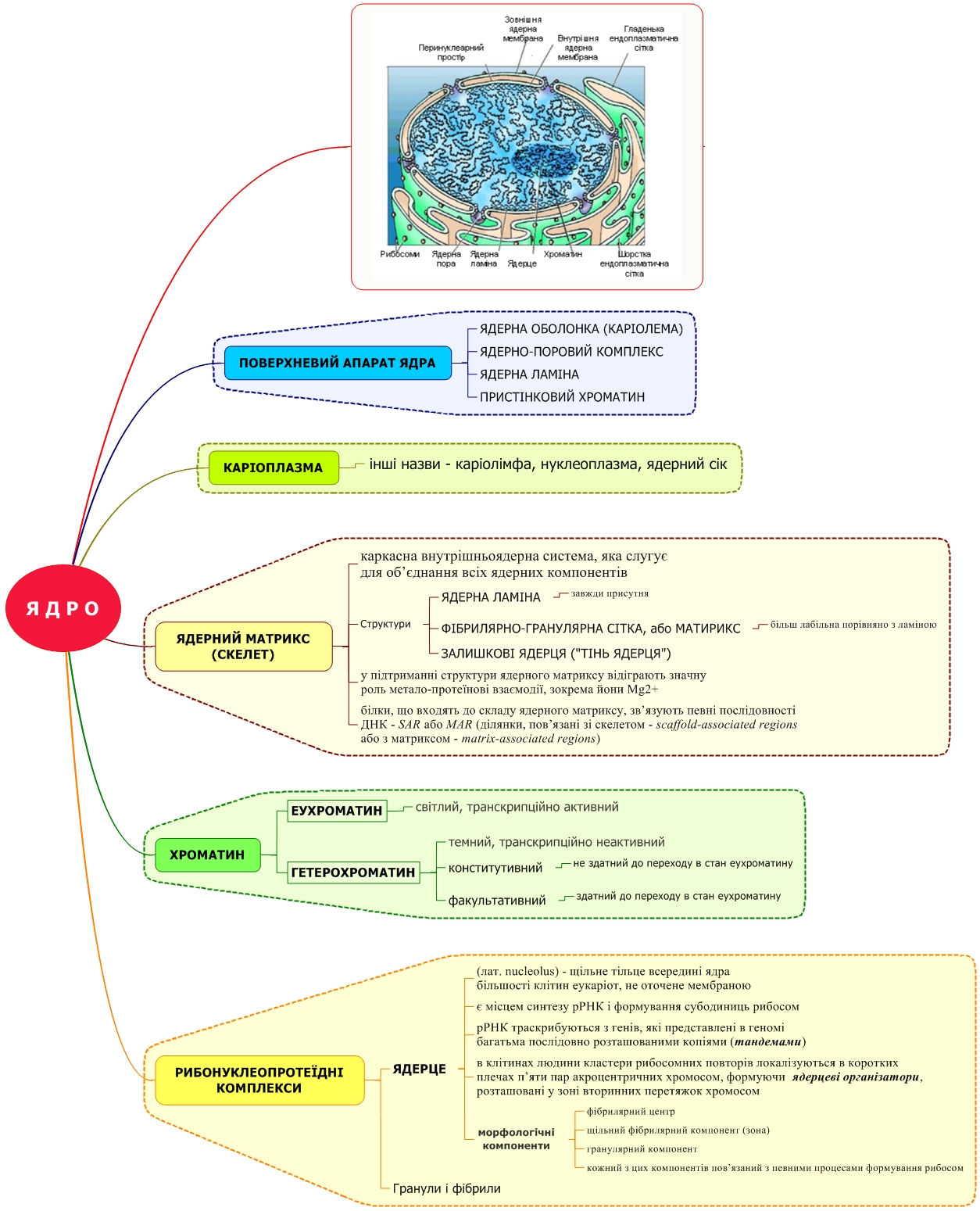
Морфологія мітотичних хромосом
У хромосомах найвищого ступеня конденсації (під час поділу) виділяють ряд важливих структурних елементів.
Ц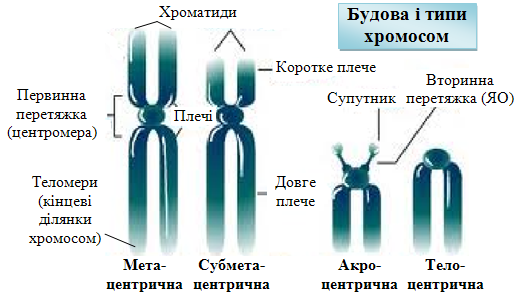 ентромери
(первинні перетяжки)
– ділянки, в яких з’єднані дві сестринські
хроматиди. Ділянки ДНК, які розташовуються
в зонах центромер, мають високоповто-рювані
послідовності.
ентромери
(первинні перетяжки)
– ділянки, в яких з’єднані дві сестринські
хроматиди. Ділянки ДНК, які розташовуються
в зонах центромер, мають високоповто-рювані
послідовності.
В залежності від розташування центромери хромосоми можуть мати плечі однакової чи різної довжини і класифікуватись на:
1) метацентричні – центромера займає центральне положення, а обидва плеча хромосоми мають майже однакову довжину;
2) субметацентричні – центромера розташовується ближче до одного з кінців, а плечі хромосоми при цьому відрізняються своїми розмірами;
3) акроцентричні – одне з її плечей є дуже коротким, а ценромера знаходиться біля кінця хромосоми;
4) телоцентричні – одне плече відсутнє, не зустрічаються в каріотипі людини
Центромера є структурою, важливою для правильного розподілу хроматид під час мітозу між дочірніми клітинами. На ній формується спеціальний білковий диск – кінетохор, до якого прикріплюються нитки веретена поділу.
Деякі хромосоми можуть мати ще й вторинну перетяжку (в цій ділянці знаходиться ДНК, яка відповідає за синтез рРНК і формування ядерця, тому її ще називають ядерцевим організатором). Ділянки хромосом, відокремлені вторинними перетяжками називаються супутниками.
Теломери локалізовані на обох кінцях кожної хроматиди всіх хромосом, складаються з множинних повторів особливих консервативних послідовностей ДНК і виконують низку дуже важливих функцій в клітині (Е. Блекберн, К. Грейдер, Дж. Шостак, Нобелівська премія, 2009):
підтримують стабільність хромосом, запобігають їх „злипанню”;
захищають кінці хромосоми від руйнування нуклеазами;
відіграють важливу роль в забезпеченні правильної реплікації кінців хромосом;
забезпечують контроль нормального клітинного росту.
Наявність теломер на кінцях хромосом – сигнал того, що хромосома нормальна. Коли в клітині з’являється пошкоджена хромосома або фрагмент хромосоми, такі структури позбавлені