

Митоз (др.-греч. μίτος — нить) — непрямое деление клетки, наиболее распространенный способ репродукции эукариотических клеток. Биологическое значение митоза состоит в строго одинаковом распределениихромосом между дочерними ядрами, что обеспечивает образование генетически идентичных дочерних клеток и сохраняет преемственность в ряду клеточных поколений[1].

Митоз — один из фундаментальных процессов онтогенеза. Митотическое деление обеспечивает рост многоклеточных эукариот за счёт увеличения популяций клеток тканей. В результате митотического деления клеток меристем увеличивается количество клеток тканей растений. Дробление оплодотворённого яйца и рост большинства тканей у животных также происходит путём митотических делений[2].
На основании морфологических особенностей митоз условно подразделяется на стадии: профазу, прометафазу, метафазу, анафазу, телофазу. Первые описания фаз митоза и установление их последовательности были предприняты в 70—80-х годах XIX века. В конце 1870-х — начале 1880-х годов немецкий гистолог Вальтер Флемминг для обозначения процесса непрямого деления клетки ввёл термин «митоз»[3].
Продолжительность митоза в среднем составляет 1—2 часа[1][4]. Митоз клеток животных, как правило, длится 30—60 минут, а растений — 2—3 часа.[5]. За 70 лет в теле человека суммарно осуществляется порядка 1014 клеточных делений[6].
История исследования
Первые неполные описания, касающиеся поведения и изменения ядер в делящихся клетках, встречаются в работах учёных начала 1870-х годов. В работе русского ботаника Э. Руссова, датируемой 1872 годом, отчётливо описаны и изображены метафазные и анафазные пластинки, состоящие из отдельных хромосом.[7] Годом позже немецкий зоолог А. Шнейдер ещё более отчётливо и последовательно, но, конечно, не совсем полно описал митотическое деление на примере дробящихся яиц прямокишечной турбеллярии Mesostomum. В его работе, в сущности, описаны и проиллюстрированы в правильной последовательности основные фазы митоза: профаза, метафаза, анафаза (ранняя и поздняя). В 1874 году московский ботаник И. Д. Чистяков также наблюдал отдельные фазы клеточного деления в спорах плаунов и хвощей. Несмотря на первые успехи ни Руссову, ни Шнейдеру, ни Чистякову не удалось дать чёткое и последовательное описание митотического деления.[8]
В 1875 году вышли работы, содержащие более детальные описания митозов. О. Бючли дал описание цитологических картин в дробящихся яйцах круглых червей и моллюсков и в сперматогенных клетках насекомых. Э. Страсбургерисследовал митотическое деление в клетках зелёной водоросли спирогиры, в материнских клетках пыльцы лука и в материнских споровых клетках плауна. Ссылаясь на работу О. Бючли и основываясь на собственных исследованиях, Э. Страсбургер обратил внимание на единство процессов клеточного деления в растительных и животных клетках.[9]
К концу 1878 — началу 1879 года появились подробные работы В. Шлейхера (о делении хрящевых клеток амфибий), В. Флемминга (о размножении клеток в разных тканях саламандры и её личинок), П. И. Перемежко (о делении клеток в эпидермисе личинок тритона). В своей работе в 1879 году Шлейхер предложил термин «кариокинез» для обозначения сложных процессов клеточного деления, подразумевая перемещения составных частей ядра.[10] Вальтер Флемминг впервые для обозначения непрямого деления клетки ввёл термин «митоз», который впоследствии стал общепринятым.[3] Также Флеммингу принадлежит окончательная формулировка определения митоза как циклического процесса, завершающегося разделением хромосом между дочерними клетками.[11]

Деление клеток по Э. Руссову (1872)
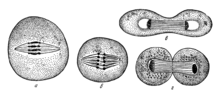
Деление клеток по Э. Страсбургеру (1875)

Деление клеток по В. Флеммингу (1882)
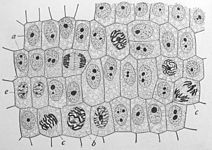
Деление клеток по Э. Б. Уилсону (1900)
В 1880 году О. В. Баранецкий установил спиральное строение хромосом. В ходе дальнейших исследований были развиты представления о спирализации и деспирализации хромосом во время митотического цикла.[11] В начале 1900-х годов хромосомы были идентифицированы в качестве носителей наследственной информации, что в дальнейшем дало объяснение биологической роли митоза, заключающейся в образовании генетически идентичных дочерних клеток.
В 1970-х годах началась расшифровка и детальное изучение регуляторов митотического деления,[12] благодаря серии экспериментов по слиянию клеток, находящихся на разных этапах клеточного цикла. В тех опытах, когда клетку в М-фазе объединяли с клеткой, находящейся в любой из стадий интерфазы (G1, S или G2), интерфазные клетки переходили в митотическое состояние (начиналась конденсация хромосом и распадалась ядерная оболочка).[13] В итоге был сделан вывод, что в цитоплазме митотической клетки присутствует фактор (или факторы), стимулирующий митоз,[14] или, иначе, М-стимулирующий фактор (МСФ, от англ. M-phase-promoting factor, MPF).[15]
Впервые «фактор стимуляции митоза» был открыт в зрелых неоплодотворенных яйцах шпорцевой лягушки, находящихся в М-фазе клеточного цикла. Цитоплазма такого яйца, инъецированная в ооцит, приводила к преждевременному переходу в М-фазу и к началу созревания ооцита (первоначально сокращение MPF означало Maturation Promoting Factor, что переводится как «фактор, способствующий созреванию»). В ходе дальнейших экспериментов были установлены универсальное значение и вместе с тем высокая степень консервативности «фактора стимуляции митоза»: экстракты, приготовленные из митотических клеток весьма разнообразных организмов (млекопитающих, морских ежей, моллюсков, дрожжей), при введении в ооциты шпорцевой лягушки переводили их в М-фазу.[16]
В ходе последующих исследований выяснилось, что фактор, стимулирующий митоз, представляет собой гетеродимерный комплекс, состоящий из белка циклина и зависимой от циклина протеинкиназы. Циклин является регуляторным белком и обнаруживается у всехэукариот. Его концентрация периодически возрастает в течение клеточного цикла, достигая максимума в метафазе митоза. С началом анафазы наблюдается резкое сокращение концентрации циклина, вследствие его расщепления с помощью сложных белковыхпротеолитических комплексов — протеосом. Зависимая от циклина протеинкиназа представляет собой фермент (фосфорилазу), модифицирующий белки за счёт переноса фосфатной группы от АТФ на аминокислоты серин и треонин. Таким образом с установления роли и структуры основного регулятора митотического деления начались исследования тонких регуляторных механизмов митоза, которые продолжаются до настоящего времени.
Типы митоза
Выработка единой типологии и классификации митозов осложняется целым спектром признаков,[~ 1] которые в различных комбинациях создают разнообразие и неоднородность картин митотического деления. При этом отдельные варианты классификации, разработанные применительно к одним таксонам, являются неприемлемыми в отношении других, поскольку не учитывают специфики их митозов. Например, отдельные варианты классификации митозов, свойственных животным или растительным организмам, оказываются неприемлемыми для водорослей.[17]
Одним из ключевых признаков, лежащих в основе различных типологий и классификаций митотического деления, является поведение ядерной оболочки. Если образование веретена и само митотическое деление протекает внутри ядра без разрушения ядерной оболочки, то такой тип митоза называют закрытым. Митоз с распадом ядерной оболочки, соответственно, называется открытым, а митоз с распадом оболочки только на полюсах веретена, с образованием «полярных окон» — полузакрытым.[18][17]
Ещё одним характерным признаком является тип симметрии митотического веретена. При плевромитозе веретено деления билатерально симметрично либо асимметрично и состоит, как правило, из двух полуверетён, располагающихся в метафазе-анафазе под углом друг к другу. Для категории ортомитозов характерна биполярная симметрия веретена деления, а в метафазе зачастую наблюдается различимая экваториальная пластинка.[18]
В рамках обозначенных признаков наиболее многочисленным является типичный открытый ортомитоз, на примере которого ниже рассматриваются принципы и стадии митотического деления. Данный тип митоза характерен для животных, высших растений и некоторых простейших.[19]
Варианты классификации митозов
7 типов митоза простейших[18]:
|
6 типов митоза водорослей[17]:
|
Основные типы митоза у водорослей: 1. Закрытый центрический; 2. Закрытый ацентрический; 3. Полузакрытый центрический; 4. Полузакрытый ацентрический; 5. Открытый центрический; 6. Открытый ацентрический |

Происхождение и эволюция митоза

Возможные пути эволюции типов митоза у простейших. В квадратных скобках изображена гипотетическая промежуточная стадия. Условные обозначения: 1. Закрытый эвгленоидный митоз; 2. Закрытый внутриядерный плевромитоз; 3. Закрытый внеядерный плевромитоз; 4. Полузакрытый плевромитоз; 5. Закрытый внутриядерный ортомитоз; 6. Полузакрытый ортомитоз; 7. Открытый ортомитоз;
Предполагается, что сложный митотический процесс высших организмов развивался постепенно из механизмов деления прокариот.[20] Это предположение подтверждается тем, что прокариоты появились приблизительно на миллиард лет раньше первых эукариот. Кроме того, в митозе эукариот и бинарном делении прокариот принимают участие схожие белки.
Возможные промежуточные стадии между бинарным делением и митозом можно проследить у одноклеточных эукариот, у которых в ходе деления не разрушается ядерная оболочка. У большинства же других эукариот, в том числе растений и животных, веретено деления формируется вне ядра, а ядерная оболочка разрушается в течение митоза. Хотя митоз у одноклеточных эукариот ещё недостаточно изучен, можно предположить, что он произошёл от бинарного деления и в конечном счёте достиг того уровня сложности, который имеется у многоклеточных организмов[21].
У многих простейших эукариот митоз также остался процессом, связанным с мембраной, однако теперь уже не плазматической, а ядерной.[22] Возможно, в связи с увеличением размера и числа хромосом, структура типамезосомы разделилась на два элемента: ЦОМТ на ядерной оболочке и кинетохор на хромосоме. Для соединения данных структур между собой в процессе эволюции развилась промежуточная система микротрубочек. В рамках данного представления, наиболее древним и примитивным считается закрытый внутриядерный плевромитоз. Сегрегация хромосом при этом происходит путём расхождения ЦОМТ, к которым хромосомы крепятся посредством микротрубочек. В свою очередь ЦОМТ закреплены на ядерной оболочке и расходятся за счёт роста ядерной мембраны между ними.[23]
От разных вариантов закрытого внутриядерного плевромитоза, вероятно, берут своё начало несколько параллельных эволюционных линий.[23] В качестве эволюционно прогрессивных признаков при этом рассматриваются: распад ядерной оболочки во время митоза; переход ЦОМТ из ядра в цитоплазму; образование биполярного веретена; усиление спирализации хромосом; формирование экваториальной пластинки в метафазе. Таким образом, эволюция митотического деления идёт в направлении от закрытого внутриядерного плевромитоза к открытому ортомитозу.[24]
Регуляция митоза
Основными регулято́рными механизмами митоза являются процессы фосфорилирования и протеолиза[25]. Обратимые реакции фосфорилирования и дефосфорилирования обеспечивают протекание обратимых событий митоза, таких как сборка/распад веретена деления или распад/восстановление ядерной оболочки. Протеолиз лежит в основе необратимых событий митоза, таких как разделение сестринских хроматид в анафазе или разрушение митотических циклинов на поздних стадиях митоза.