

5. Физико-химические механизмы возникновения потенциала действия
Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны. При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный. Компоненты потенциала действия: 1) локальный ответ; 2) высоковольтный пиковый потенциал (спайк); 3) следовые колебания: а) отрицательный следовой потенциал; б) положительный следовой потенциал.
Локальный ответ
Пока раздражитель не достиг на начальном этапе 50–75 % от величины порога, проницаемость клеточной мембраны остается неизменой, и электрический сдвиг мембранного потенциала объясняется раздражающим агентом. Достигнув уровня 50–75 %, открываются активационные ворота (m-ворота) Na-каналов, и возникает локальный ответ. Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку. Если сила раздражения недостаточна, то локального ответа не происходит.
Высоковольтный пиковый потенциал (спайк)
Пик
потенциала действия является постоянным
компонентом потенциала действия.
Он
состоит из двух фаз:
1)
восходящей части – фазы деполяризации;
2)
нисходящей части – фазы
реполяризации.
 Лавинообразное
поступление ионов Na в клетку приводит
к изменению потенциала на клеточной
мембране. Чем больше ионов Na войдет в
клетку, тем в большей степени деполяризуется
мембрана, тем больше откроется
активационных ворот. Постепенно заряд
с мембраны снимается, а потом возникает
с противоположным знаком. Возникновение
заряда с противоположным знаком
называется инверсией потенциала
мембраны. Движение ионов Na внутрь клетки
продолжается до момента электрохимического
равновесия по иону Na. Амплитуда потенциала
действия не зависит от силы раздражителя,
она зависит от концентрации ионов Na и
от степени проницаемости мембраны к
ионам Na. Нисходящая фаза (фаза реполяризации)
возвращает заряд мембраны к исходному
знаку. При достижении электрохимического
равновесия по ионам Na происходит
инактивация активационных ворот,
снижается проницаемость к ионам Na и
возрастает проницаемость к ионам K,
натрий-калиевый насос вступает в действие
и восстанавливает заряд клеточной
мембраны. Полного восстановления
мембранного потенциала не происходит.
В
процессе восстановительных реакций на
клеточной мембране регистрируются
следовые потенциалы – положительный
и отрицательный. Следовые потенциалы
являются непостоянными компонентами
потенциала действия. Отрицательный
следовой потенциал – следовая
деполяризация в результате повышенной
проницаемости мембраны к ионам Na, что
тормозит процесс реполяризации.
Положительный следовой потенциал
возникает при гиперполяризации клеточной
мембраны в процессе восстановления
клеточного заряда за счет выхода ионов
калия и работы натрий-калиевого насоса.
Лавинообразное
поступление ионов Na в клетку приводит
к изменению потенциала на клеточной
мембране. Чем больше ионов Na войдет в
клетку, тем в большей степени деполяризуется
мембрана, тем больше откроется
активационных ворот. Постепенно заряд
с мембраны снимается, а потом возникает
с противоположным знаком. Возникновение
заряда с противоположным знаком
называется инверсией потенциала
мембраны. Движение ионов Na внутрь клетки
продолжается до момента электрохимического
равновесия по иону Na. Амплитуда потенциала
действия не зависит от силы раздражителя,
она зависит от концентрации ионов Na и
от степени проницаемости мембраны к
ионам Na. Нисходящая фаза (фаза реполяризации)
возвращает заряд мембраны к исходному
знаку. При достижении электрохимического
равновесия по ионам Na происходит
инактивация активационных ворот,
снижается проницаемость к ионам Na и
возрастает проницаемость к ионам K,
натрий-калиевый насос вступает в действие
и восстанавливает заряд клеточной
мембраны. Полного восстановления
мембранного потенциала не происходит.
В
процессе восстановительных реакций на
клеточной мембране регистрируются
следовые потенциалы – положительный
и отрицательный. Следовые потенциалы
являются непостоянными компонентами
потенциала действия. Отрицательный
следовой потенциал – следовая
деполяризация в результате повышенной
проницаемости мембраны к ионам Na, что
тормозит процесс реполяризации.
Положительный следовой потенциал
возникает при гиперполяризации клеточной
мембраны в процессе восстановления
клеточного заряда за счет выхода ионов
калия и работы натрий-калиевого насоса.
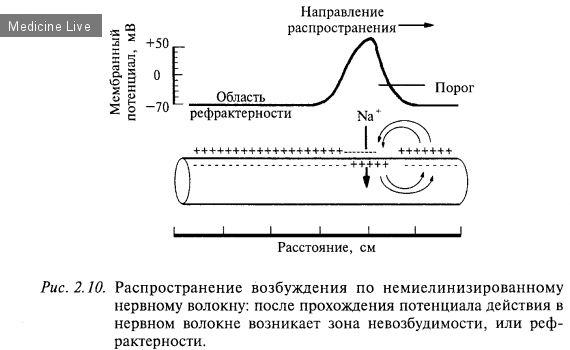
3.Фазы возбудимости.
Возбудимость – это способность отвечать возбуждением (генерацией потенциала действия) в ответ на действие раздражителей любой природы.
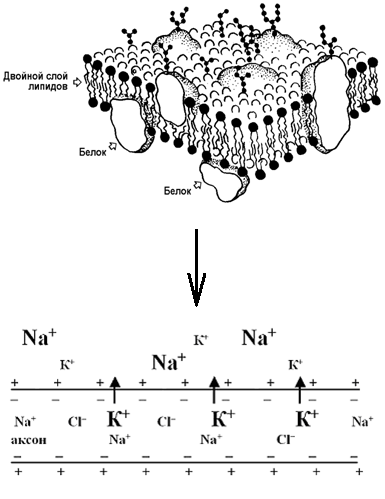
Рисунок 9 – Модель биологической мембраны и механизм возникновения потенциала покоя
Клетки сердечной мышцы, как и любой другой возбудимой ткани, поляризованы. Мембрана кардио-миоцитов снаружи заряжена поло-жительно, изнутри – отрицательно. Это обусловлено различным содержанием ионов натрия (Na+) и калия (K+) по обе стороны мембраны – внутри сердечных клеток больше K+, а снаружи - Na+(рисунок 9). В покое мембрана кардиомиоцитов непроницаема для ионов Na+, но частично пропускает ионы K+. В результате процесса диффузии в соответствии с концентрационным градиентом ионы K+выходят из кардиомиоцита, увеличивая положительный заряд на его поверхности. Внутренняя сторона мембраны при этом становится отрицательной за счет скопления анионов кислот (хлоридов - Cl-, сульфатов - SO42-, фосфатов - PO43- и т.д.), для которых мембрана не проницаема. У клеток рабочего миокарда потенциал покоя составляет – 90 мВ.
При действии какого-либо раздражителя (электрические импульсы, гуморальные факторы и т.д.) кардиомиоциты приходят в состояние возбуждения, внешним выражением которого является развитие на поверхности их мембраны потенциала действия. Механизм деполяризации кардиомиоцитов имею ту же природу, что и потенциал действия развивающийся в нервных и скелетно-мышечных волокнах.
В потенциале действия различают следующие фазы (рисунок 10):
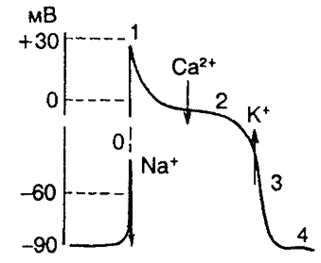
Рисунок 10 - Потенциал действия одиночного кардиомиоцита желудочка 0-4 – фазы потенциала действия кардиомиоцита
Фаза 0 – деполяризация, которая характеризуется повышением натриевой проницаемости за счет активации быстрых натриевых каналов клеточных мембран. В этот период Na+лавинообразно входит в клетку. Эта фаза заканчивается достиже-нием критического уровня деполяризации, при котором происходит изменение знака мембранного потенциала (с -90 мВ до +30 мВ).
Фаза 1 – быстрая начальная реполяризация – связана с активацией медленных натриевых и кальциевых каналов;
Фаза 2 – медленная реполяризация (плато), характеризующееся дальнейшим повышением входа в клетку ионов кальция (Са2+). В период плато натриевые каналы инактивируются и клетка находится в состоянии абсолютной невозбудимости или рефрактерности.
Фаза 3 – быстрая конечная реполяризация обусловлена активацией калиевых каналов. В период фазы 3 закрываются кальциевые каналы за счет чего падает кальциевый ток, дополнительно деполяризующий мембрану. Это ускоряет процесс реполяризации;
Фаза 4 – потенциал покоя, в период которого за счет работы калий-натриевого насоса полностью восстанавливается градиент концентраций Na+ и K+ по обе стороны мембраны. Калий-натриевый насос представляет собой белок встроенный в мембрану, который работает таким образом, что выкачивает из клетки 3 иона Na+ и закачивает обратно 2 иона K+.
Развитие возбуждения в клетках рабочего миокарда и в пейсмейкерах проводящей системы существенно различается. Отличительной особенностью клеток проводящей системы является то, что потенциал действия у них развивается в период покоя – спонтанная диастолическая деполяризация. На этом механизме обусловлена их авторитмическая активность. Такая особенность развития возбуждения в пейсмейкерах обусловлена следующими характерными только для них свойствами (рисунок 11):

Рисунок 11 – Потенциал действия пейсмейкера синусно-предсердного узла. Стрелками показа медленная диастолическая деполяризация
они имеют потенциал покоя от – 55 мВ до – 60 мВ (в отличие от сократительных волокон миокарда, обладающих потенциалом покоя от – 85 мВ до – 90 мВ;
их мембрана обладает повышенной проницаемостью для ионов Na+ по сравнению с другими клетками миокарда;
в связи с вышеуказанными свойствами они не способны поддерживать постоянный потенциал действия;
потенциал действия в них, называемый «кардиостимулирующим потенциалом» имеет малую крутизну подъема;
они имеют слабовыраженную стадию реполяризации: фаза медленной реполяризации (плато) почти отсутствует, за ней сразу следует фаза быстрой реполяризации, во время которой мембранный потенциал покоя достигает уровня -50 мВ – -60 мВ (вместо -85 мВ – -90 мВ в рабочем миокарде), после чего вновь начинается фаза диастолической деполяризации.
Возбудимость сердечной мышцы развивается циклически, что выражается законом периодической невозбудимости сердца или рефлексом Мэрея:
в систоле отсутствует возбудимость миокарда;
в диастоле сердечная возбудимость достигает самых высоких уровней.
С момента возникновения потенциала действия, который длится около 0,3 с и до конца его плато (во время фаз 0, 1 и 2) мембрана кардиомиоцитов становится невосприимчивой к действию других раздражителей, т.е. находится в абсолютной рефрактерности. Соотношения между потенциалом действия клеток миокарда, его сокращением и динамикой возбудимости показаны на рисунке 12. Различают период абсолютной рефрактерности (полная невозбудимость); период относительной рефрактерности, во время которой сердечная мышца может отвечать сокращением лишь на очень сильные раздражения и соответствует фазе быстрой реполяризации; период супернормальной возбудимости, когда сердечная мышца может отвечать сокращением на подпороговые раздражения.
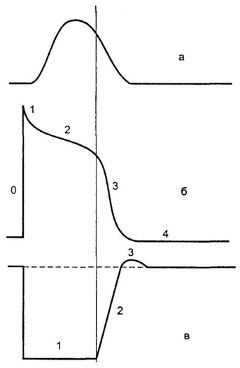
Рисунок 12 – Соотношение между механокардиограммой, потенциалом действия кардиомиоцита и динамикой возбудимости миокарда а – механокардиограмма; б – потенциал действия кардиомиоцита: 0-4 фазы потенциала действия, в – динамика возбудимости: пунктирной линией обозначена исходная возбудимость, 1 – абсолютная рефрактерность, 2 – относительная рефрактерность, 3 – супернормальная возбудимость. Вертикальная линия, проходящая через все кривые демонстрирует, что начало восстановления возбудимости {граница между абсолютной и относительной рефрактерностью (кривая в) соответствует началу фазы 3 потенциала действия (кривая б) и началу диастолы (кривая а)}.
Сокращение (систола) миокарда продолжительностью 0,3 с по времени примерно совпадает с длительностью общей рефрактерности, представляющей собой сумму абсолютной и относительной рефрактерности. Следовательно, в периоде сокращения сердце не способно реагировать на другие раздражители. Наличие длительной рефрактерной фазы препятствует развитию непрерывного укорочения (тетанус) сердечной мышцы, что привело бы к невозможности осуществления сердцем нагнетательной функции.
Если нанести внеочередное раздражение на миокард в период расслабления (диастолы), когда его возбудимость частично или полностью восстановлена, сердечная мышца отвечает экстрасистолой, которая имеет менее выраженную амплитуду, чем обычная систола. При этом, после любой экстрасистолы всегда следует удлиненный период покоя. Компенсаторная длительная пауза наступает из-за потери физиологической систолы генерируемой синусно-предсердным узлом Кейт-Флака.
Динамика изменения возбудимости миокарда соответствует фазам сердечного цикла. Сердечный цикл представляет собой многократно повторяющуюся последовательность сокращения (систолы) и расслабления (диастолы) с последующим наполнением предсердий и желудочков кровью, которая затем в определенном количестве выбрасывается сердцем в аорту и легочную артерию.
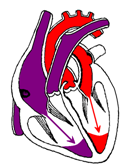
Продолжительность сердечного цикла равно примерно 0,8 с. Цикл состоит из трех основных повторяющихся этапов: систолы предсердий, систолы желудочков, общей паузы. Началом каждого цикла считается систола предсердий, длящаяся 0,1 с. Давление в предсердиях при этом поднимается до 5-8 мм рт.ст. Обратному току крови из предсердий препятствует замыкание в период их систолы сфинктеров, расположенных в устьях полых вен. Желудочки в этот момент расслаблены, атриовентрикулярные клапаны свободно свисают в их полость вследствие чего возможно свободное поступление крови в желудочки (рисунок 13а). При этом, полулунные клапаны закрыты вследствие затекания в них крови в период предыдущей систолы желудочков.
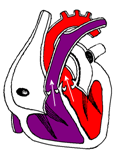
Систола предсердий сменяется систолой желудочков продолжительностью 0,33 с (рисунок 13б). В момент систолы желудочков предсердия оказываются уже расслабленными. Из-за наличия определенной задержки проведения импульса возбуждения через атриовентрикулярный узел сокращения желудочков начинается заведомо позже сокращения предсердий. Систола желудочков разделяется на периоды напряжения и изгнания. Период напряжения (0,08 с) представляет собой период развития давления, которое должно превысить уровень давления в основных артериальных стволах (аорта и легочная артерия). В этом периоде выделяют фазу асинхронного сокращения (0,05 с) и фазу изометрического сокращения (0,03 с) миокарда.
|
а |
Систола предсердий Диастола желудочков |
|
б |
Диастола предсердий Систола желудочков: период напряжения = фаза асинхронного сокращения + фаза изометрического сокращения |
|
в |
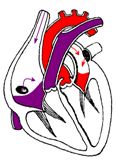
Диастола предсердий Систола желудочков: период изгнания = фаза быстрого изгнания + фаза медленного изгнания |
|
г |
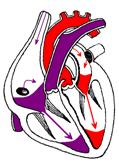
Диастола предсердий Диастола желудочков: протодиастолический период; изометрическое расслабление |
|
д |
Диастола предсердий Диастола желудочков: фаза быстрого наполнения + фаза медленного наполнения кровью |
|
|
Рисунок 13 (а-д) – Сердечный цикл а-д – циклическая последовательность смены фаз сердечного цикла |

По мере повышения давления в желудочках возрастает сопротивление их мышечных слоев, что способствует захлопыванию атриовентрикулярных клапанов. В этот момент полулунные клапаны еще также закрыты, поэтому полость желудочка оказывается замкнутой и объем крови в полости остается постоянным. Таким образом «вход» и «выход» из желудочков оказывается закрытым. Вследствие того, что кровь, как любая жидкость, практически несжимаема, длина волокон миокарда не изменяется, увеличивается их только напряжение (изометрическое напряжение).
Это способствует стремительному росту давления в желудочках. В силу неравномерного вовлечения в процесс сокращения левого желудочка форма сердца изменяется, верхушка подтягивается к основанию и форма сердца из эллипсовидной становится более близкой к шарообразной. Это облегчает изгнание крови при развитии силы основным слоем левого желудочка – циркулярным. В силу этих изменений сердце стенкой левого желудочка ударяется о внутреннюю поверхность грудной клетки (первый систолический тон сердца). Поскольку в этом периоде изменяется форма левого желудочка, а значит, и длина некоторых мышечных волокон, не вполне правильно считать данные период периодом изометрического сокращения по аналогии с изолированной мышцей.
К концу периода напряжения быстро нарастающее давление в левом и правом желудочках становится выше давления в аорте и легочном стволе, полулунные клапаны открываются и кровь из желудочков устремляется в эти сосуды. Начинается период изгнания крови, длящийся примерно 0,25 с (рисунок 13в). Период изгнания крови из желудочков состоит из фазы быстрого (0,12 с) и фазы медленного изгнания (0,13 с). В начале периода изгнания давление крови в полости желудочков продолжает нарастать, достигая примерно 130 мм рт.ст. в левом и 25 мм рт.ст. в правом. В результате кровь быстро изливается в аорту и легочную артерию, объем желудочков резко уменьшается. Это фаза быстрого изгнания. После открытия полулунных клапанов выброс крови из сердца начинает замедляться, сокращение миокарда желудочков слабеет, наступает фаза медленного изгнания.
Таким образом, выброс крови их желудочков происходит в начале с максимальной скоростью, которая постепенно снижается в силу того, что градиент давления между левым желудочком и аортой, являющийся движущейся силой для изгнания, столь же быстро уменьшается. Изгнание завершается как следствие исчезновения градиента давления в связи с ограниченной растяжимостью аорты, так и вследствие прекращения сокращения миокардиальных клеток.
В конце фазы медленного изгнания миокард желудочков начинает расслабляться, наступает диастола (0,47 с). Давление в желудочках падает, кровь из аорты и легочного ствола устремляется обратно в полсти желудочков и захлопывает полулунные клапаны, препятствуя обратному току крови из этих сосудов. При этом возникает второй диастолический тон сердца.
Время от начала расслабления желудочков до захлопывания полулунных клапанов называется протодиастолическим периодом (0,04 с). Таким образом, снова наступает короткий период, во время которого закрыты аортальные клапаны и еще не открылись атриовентрикулярные (рисунок 13г). При этом давление в желудочках продолжает снижаться. Длина волокон миокарда не изменяется, поэтому данный период назван периодом изометрического расслабления (0,08 с). Когда же окажется, что давление в желудочках чуть меньше, чем в предсердиях, открываются предсердно-желудочковые клапаны и кровь из предсердий устремляется в желудочки.
Начинается период наполнения желудочков кровью, которая будет выброшена в следующем цикле. Этот период длится 0,25 с и делится на фазы быстрого (0,08с) и медленного (0,17 с) наполнения. Колебания стенок желудочков вследствие быстрого притока крови к ним вызывают появление третьего тона сердца. Таким образом, фаза быстрого наполнения обусловлена градиентом давления между предсердиями и желудочками и поэтому протекает пассивно, без сократительной работы кардиомиоцитов. Тот период, когда желудочки пассивно заполняются кровью, но предсердия еще не сокращаются, называется общей диастолой всего сердца (рисунок 13д).
К концу фазы медленного наполнения возникает систола предсердий, во время которой кровь дополнительно закачивается в желудочки. Таким образом начинается новый цикл.
Поскольку в основе возбудимости сердечной мышцы лежат электрофизиологические процессы возбуждение сердца может быть вызвано воздействием электрического тока. Под действием электрических импульсов, источником которых являются электронные стимуляторы, сердце начинает возбуждаться и работать в заданном режиме. Такой способ воздействия на сердце нашел практическое применение в медицине. Например, в патологии можно наблюдать состояние сердечной мышцы, когда она сокращается чрезвычайно часто – до 400 (при трепетании) и до 600 (при мерцании или фибрилляции) в 1 мин. При этом каждое мышечное волокно сокращается асинхронно. При таком сокращении сердце не может осуществлять нагнетание крови. У человека, как правило, такое состояние смертельно, если не принять быстро меры для его прекращения. Единственным эффективным способ в таких случаях является воздействие сильным (напряжение в несколько киловольт) ударом электрического тока, вызывающим одновременно возбуждение всех мышечных волокон миокарда, после чего восстанавливается синхронность их сокращения.
С помощью воздействия электрических стимулов можно также изучать динамику изменения возбудимости сердечной мышцы на различных стадиях цикла.
4.Скелетные мышцы и их функции и свойства.