

152.Теорія походження еукаріот
Еукаріотичні організми - це організми, що мають оформлене ядро. До еукаріотів відносяться гриби, рослини, тварини. Відмінності між прокариотами і еукаріотами настільки кардинальні, що їх відносять до різних надцарству живої природи.
Для еукаріот характерні такі ознаки:
1. ДНК лінійна.
2. ДНК завжди в комплексі з білками гистонами (ДНП).
3. ДНК входить до складу хромосом.
4. Є органели.
5. Рибосоми крупніше прокариотических.
6. Є мітоз.
7. Є мейоз.
8. Є гамети.
Гіпотези виникнення життя на Землі не внесли ясності в питання про виникнення клітини.. Що стосується походження клітин еукаріот, то з цієї проблеми існує дві точки зору. Згідно з першою гіпотезою, всі клітинні органели ведуть своє походження від плазмалеми: вони утворилися шляхом впячувания окремих ділянок і подальшої диференціації та спеціалізації. Л. Моргуліс запропонувала СЕТ-теорію - теорію серії ендосімбіогенезів (серіальна ендосімбіогенетіческая теорія). Згідно СЕТ-теорії, становлення клітини еукаріот відбувалося в кілька етапів на основі симбіозу (мутуалізму).
В результаті адаптивної радіації прокаріотів - їх екологічної диференціювання - виникло колосальне розмаїття цих організмів. З'явилися бактерії хемосинтетики, фотосинтетики, аероби (грам-негативні бактерії, що мають цикл Кребса), анаероби.
Базою для ендосимбіоз послужили мікоплазмоподібні прокаріоти, які могли сбраживать глюкозу до двох-і трехуглсродних кінцевих продуктів, т. к. вони володіли метаболічним шляхом Ембдена-МейергофаПарнаса. На першому етапі ендосімбіогенеза аеробні паличкоподібні прокаріоти були захоплені, але не переварені мікоплазмоподобнимі прокариотами. Прокаріоти не здатні до фагоцитозу, але мікоплазми мають тільки плазмалемму і тому - мінливу форму тіла. Завдяки володінню ендосімбіонтов циклом трікарбо-нових кислот (циклом Кребса) вони стали промітохоп-дріями, а потім мітохондріями. На другому етапі ендосімбіогенеза виник дігеномиий організм вступив в постійний контакт з бактеріями спірохетами, точніше, спіроплазма. З часом вони перетворилися в джгути еукаріотичних клітин. Ядерна оболонка з'явилася як захист від багатьох ферментів ендосімбіонтів - мітохондрій. Виникла еукаріотична клітина стала вихідною для тварин і грибів. Становлення рослинної клітини сталося в три етапи. Трігеномний організм вступив в симбіоз з ціанобактеріями, які з часом перетворилися на хлоропласти.
Докази на користь СЕТ-гіпотези: наявність у мітохондрій і пластид подвійної оболонки (власної та ва-куолярной), наявність власної кільцевої - прокариотической ДНК, наявність дрібних прокариотических рибосом, незалежний від ядра ритм розмноження мітохондрій і пластид.
ПОХОДЖЕННЯ еукаріотичної клітини
Викопні останки клітин еукаріотичного типу виявлені в породах , вік яких не перевищує 1,0-1,4 млрд. років. Більш пізніше виникнення , а також схожість в загальних рисах їх основних біохімічних процесів ( самоподвоєння ДНК , синтез білка на рибосомах ) змушують думати про те , що еукаріотичні клітини походять від предка , що мав прокаріотичну будову.
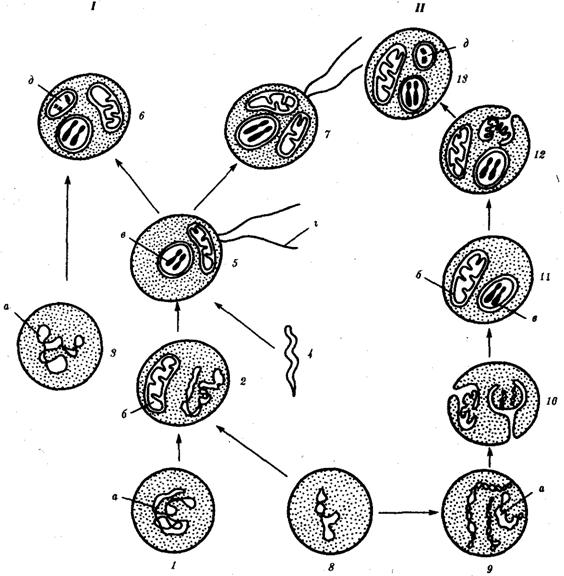
Найбільш популярна в даний час симбиотическая гіпотеза походження еукаріотичних клітин , згідно якої ( рис. 1.4) основою , або клітиною - господарем , в еволюції клітини еукаріотичного типу послужив анаеробний прокаріот , здатний лише до амебоідного руху . Перехід до аеробного дихання пов'язаний з наявністю в клітині мітохондрії , які відбулися шляхом змін симбіонтів - аеробних бактерій , що проникнули в клітину- господаря і співіснували з нею
 .
.
Рис. 1.4. Происхождение эукариотической клетки согласно симбиотической (I) и инвагинационной (II) гипотезам:
1 — анаэробный прокариот (клетка-хозяин), 2 — прокариоты, имеющие митохондрии, 3 — сине-зеленая водоросль (презумптивный хлоропласт), 4 —сиирохетообразная бактерия (презумпгивный жгутик), 5 — примитивный эукариот со жгутиком, 6 — растительная клетка, 7 — животная клетка со жгутиком, 8 — аэробный прокариот (презумптивная митохондрия), 9 — аэробный прокариот (клетка-родоначальница согласно гипотезе II), 10 —инвагинации клеточной оболочки, давшие ядро и митохондрии, 11 — примитивный эукариот 12 — впячивание клеточной оболочки, давшее хлоропласт, 13 — растительная клетка; а—ДНК прокариотической клетки, б — митохондрия, в —ядро эукариотической клетки, г —жгутик, д — хлоропласт
Серйозним аргументом на користь симбіотичного походження мітохондрій , центриолей і хлоропластів є те , що перераховані органели мають власну ДНК. Разом з тим білки баціллін і тубулін , з яких складаються джгутики і війки відповідно сучасних прокаріот і еукаріот , мають різну будову. У бактерій не знайдено також структур з властивою жгутикам , війкам , базальним тільцям і центриолям еукаріотичних клітин комбінацією мікротрубочок : « 9 + 2 » або « 9 + 0 ».
Центральним і важким для відповіді є питання про походження ядра. Припускають , що воно також могло утворитися з симбіонту - прокаріоти . Збільшення кількості ядерної ДНК , у багато разів перевищує в сучасній еукаріотичної клітини її кількість в мітохондрій або хлоропласті , відбувалося , очевидно , поступово шляхом переміщення груп генів з геномів симбіонтів . Не можна виключити , однак , що ядерний геном формувався шляхом нарощування генома клітини - господаря ( без участі симбіонтів ) .
Згідно інвагінаційної гіпотези , предковою формою еукаріотичної клітини був аеробний прокаріот (рис. 1.4) . Усередині такої клітини -хазяїна знаходилося одночасно кілька геномів , що спочатку прикріпилися до клітинної оболонки . Органели , що мають ДНК , а також ядро , виникли шляхом впячування і відшнуровування ділянок оболонки з подальшою функціональною спеціалізацією в ядро , мітохондрій , хлоропласти . У процесі подальшої еволюції сталося ускладнення ядерного геному , з'явилася система цитоплазматичних мембран.
Інвагінаційна гіпотеза добре пояснює наявність в оболонках ядра , мітохондрій , хлоропластів , двох мембран. Однак вона не може відповісти на питання, чому біосинтез білка в хлоропластах і мітохондріях в деталях відповідає такому в сучасних прокариотических клітинах , але відрізняється від біосинтезу білка в цитоплазмі еукаріотичної клітини.
Теорія симбіотичного походження еукаріотичної клітини
Існує гіпотеза про те , що певною мірою автономні і несучі певну кількість власної ДНК мітохондрії і пластиди представляють собою видозмінені прокаріотичні організми , які знайшли "притулок" в крупніших гетеротрофних клітинах -господарях - попередниках еукаріот . Все, або майже все , що нині живуть еукаріоти містять у своїх клітинах мітохондрії , а все автотрофні еукаріоти - також і пластиди . Можливо , вони були придбані в результаті незалежних випадків симбіозу , точніше - внутрішнього симбіозу ( ендосимбіоз ) . Більші гетеротрофні клітини , попередниці еукаріотичних клітин , очевидно , захищали свої симбіотичні органели від різних несприятливих умов навколишнього середовища. У свою чергу , прокариотические симбіонти виявилися корисними завдяки здатності використовувати енергію сонячного світла ( фотосинтез ) і можливості використовувати молекулярний кисень для окислення органічних речовин. В результаті еукаріоти змогли заселити сушу , а також ту частину водного середовища (зазвичай з високою рН) , де прокаріоти щодо нечисленні.