

Глава 3 Физиология канонических и неканонических вирусов
Наиболее общими физиологическими свойствами канонических и большинства неканонических вирусов являются: 1) автономность генетических структур, способных функционировать и репродуцироваться в восприимчивых к ним клетках животных, растений, простейших, грибов, бактерий; 2) отсутствие собственных метаболических систем, обеспечивающих питание, дыхание, рост, размножение и репарацию поврежденного генома. Исключение составляют прионы, не имеющие в своем составе ни автономных генетических структур, ни собственных метаболических систем.
§ 1. Физиология канонических вирусов.
Вне живой клетки вирионы канонических вирусов покоятся без признаков жизни и движения; они не размножаются, не питаются, не дышат; то есть проявляют себя как неживая материя. Но вирусы мгновенно «оживают» при их проникновении в про- или эукариотические клетки. При этом за короткое время в клетках образуется многочисленное вирусное потомство, нередко с измененной первичной структурой генома.
Канонические вирусы – это примитивные живые организмы, обладающие, за исключением метаболизма, основными признаками живой материи: органической структурой, наследственностью и изменчивостью. Канонические вирусы являются внутриклеточными генетическими паразитами, поскольку способны только размножаться, используя для этого метаболические системы, энергетический и пластический материал живых про- и эукариотических клеток. По своей сути вирус представляет собой внеклеточную органеллу, которая очень хорошо приспособлена для переноса нуклеиновой кислоты от одной клетки к другой.
Основными структурными и функциональными отличиями канонических вирусов от прокариотов и эукариотов являются: наличие только одного типа нуклеиновой кислоты – ДНК или РНК; отсутствие клеточного строения; неспособность к росту и бинарному делению; отсутствие собственных метаболических систем; воспроизведение потомства за счет одной нуклеиновой кислоты, а не за счет своих составных частей; использование пластического материала, энергии и рибосом клетки-хозяина для синтеза собственных белков и нуклеиновых кислот; воспроизведение вирусов происходит только в клетках более высоко организованных живых существ.
Репродукция вирусов. Вирусы размножаются уникальным способом, получившим название дизъюнктивной репродукции. Репродукция – это размножение вирусов путем многократного копирования вирусной геномной нуклеиновой кислоты, копии которой являются и геномом для новых вирионов, и матрицей для синтеза вирусных белков. Репродукция является дизъюнктивной, то есть разобщенной, потому что репликация вирусных нуклеиновых кислот и биосинтез вирусных белков происходят в разных частях клетки – ядре и цитоплазме. В процессе дизъюнктивной репродукции вирусы подчиняют генетические механизмы инфицированной клетки для реализации вирусной генетической информации в ущерб генетических программ клетки-хозяина.
У подавляющего большинства вирусов размножение обеспечивают проникающие в клетку-мишень вирионы, которые содержат геномную ДНК или РНК. У некоторых вирусов (пикорнавирусы) геномные нуклеиновые кислоты являются инфекционными. Они способны самостоятельно проникать в чувствительные клетки и обеспечивать полноценную репродукцию вирусов. Например, удельная инфекционность изолированных РНК пикорнавирусов составляет одну миллионную часть от инфекционности вириона.
Репродуктивный цикл вирусов складывается из нескольких стадий: 1) адсорбция; 2) пенетрация (проникновение в клетку); 3) дезинтеграция («раздевание», депротеинизация) вириона; 4) синтез вирусных белков и нуклеиновых кислот; 5) сборка вириона; 6) выход вириона из клетки. Три первых стадии являются ранними, подготовительными, обеспечивающими проникновение вирусного генома в чувствительную клетку. В течение четвертой-шестой стадий, считающихся поздними, происходит реализация стратегии вирусного генома максимально эффективным путем.
1 стадия – адсорбция, или прикрепление вирионов к чувствительной клетке, является высокоспецифическим процессом, в реализации которого принимают участие вирусные и клеточные рецепторы. Вирусные рецепторы – это преимущественно белки или гликопротеины, располагающиеся в виде шипиков на поверхности капсида и суперкапсида простых и сложных вирионов соответственно. Рецепторы клеток могут быть представлены белками, липидами или гликолипидами, которые расположены в особых углублениях плазмолеммы. Вирусные и клеточные рецепторы подходят друг к другу, как «ключ к замку», что обеспечивает тропизм вирусов к клеткам определенных органов и тканей. Напротив, несоответствие рецепторов лежит в основе видового иммунитета. Рецепторы являются высоко консервативными, изменение химической структуры или пространственной конфигурации узнающих участков рецепторов может привести к изменению тропизма вируса, либо к потере его инфекционности. Процесс адсорбции не требует затрат энергии и протекает в 2 фазы: 1) фаза притяжения, обусловлена электростатическими силами разнозаряженных ионных группировок; 2) фаза прикрепления, происходит благодаря структурной гомологии вирусных и клеточных рецепторов. Процесс адсорбции может быть блокирован путем экранирования рецепторов специфическими антителами, либо путем разрушения рецепторов. Кроме прикрепления, рецепторы необходимы для транспортировки вирионов внутрь клетки.
Адсорбция инфекционных вирусных нуклеиновых кислот также происходит путем рецепторного контакта.
2-я стадия – пенетрация. Осуществляется 4 способами: 1) виропексисом; 2) слиянием мембран; 3) трансфекцией; 4) инокуляцией. Виропексисом и слиянием мембран в клетку проникают только вирионы, при этом способ проникновения определяется строением вирусной оболочки. Трансфекция характерна исключительно для изолированных инфекционных вирусных геномов, инокуляция – для бактериофагов.
Путем слияния мембран в клетку проникают сложные вирионы, имеющие гликолипидный суперкапсид (парамиксо-, герпесвирусы, ВИЧ-1). После рецепторной адсорбции таких вирионов под действием F-белков (белков слияния) происходит соединение суперкапсида и плазмолеммы клетки-мишени. Суперкапсид встраивается в плазмолемму и «раскрывается», вследствие чего нуклеокапсид проникает непосредственно в цитоплазму клетки. Процессу слияния способствуют кислые значения рН. Поскольку суперкапсиды содержат специфические вирусные антигены и остаются в составе плазмолеммы, инфицированная клетка становится мишенью для антигензависимых Т-лимфоцитов/киллеров.
Виропексисом (рецепторным эндоцитозом, пиноцитозом) в клетки проникают как простые, не имеющие суперкапсида, так и сложные вирионы. Виропексис простых вирионов. В месте рецепторной адсорбции цитолемма клетки инвагинирует, образуя вакуоль, содержащую вирион. Фагосома отделяется от цитолеммы, погружается в цитоплазму и соединяется с лизосомой, образуя фаголизосому. В кислом содержимом фаголизосомы происходит «раздевание» вириона. Виропексис сложных вирионов происходит аналогичным образом, при этом в фагосому заключается вирион вместе с суперкапсидом. Это отсрочивает иммунный ответ макроорганизма, поскольку вирусные антигены на клеточной поверхности не остаются, как это имеет место при проникновении путем слияния мембран. После образования фаголизосомы, благодаря суперкапсидным F-белкам происходит слияние липидных слоев суперкапсида и мембраны лизосомы. В результате этого нуклеокапсид оказывается в цитоплазме клетки, а суперкапсид остается в составе фаголизосомы.
Трансфекция. Инфекционные вирусные нуклеиновые кислоты проникают в чувствительную клетку, как и многие сложные биомолекулы, путем активного транспорта через каналы в цитолемме. Этот процесс сопровождается затратой энергии и привлечением ферментативных транспортных систем клетки-хозяина.
Инокуляция. Характерна для бактериофагов. После адсорбции на бактериальной клетке чехол отростка бактериофага сокращается, и стержень отростка прободает клеточную оболочку. Через полость стержня внутрь бактерии проникает вирусный геном, что исключает стадию дезинтеграции.
3-я стадия – дезинтеграция («раздевание», депротеинизация) вириона. Дезинтеграция – это процесс полного или частичного разрушения капсида, сопровождающийся выходом геномной ДНК или РНК из вириона в цитоплазму или нуклеоплазму клетки. В зависимости от способа пенетрации и места репликации вирусов дезинтеграция вирионов происходит различно.
Дезинтеграция вирионов, проникших в клетку путем виропексиса. В результате образования фаголизосомы вирионы оказываются в кислой среде (рН=4-5), обусловленной содержимым лизосомы (рис. 1). Кислую реакцию среды создают водородные ионы органических и неорганических кислот, что необходимо для активации протеолитических и других ферментов, также находящихся в лизосоме. Для разрушения белка необходимо, чтобы активные центры протеолитических ферментов соединились с участками диссоциации белковой молекулы. Однако участки диссоциации капсидных белков обращены внутрь капсида, что делает их для лизосомальных ферментов недоступными. Вместе с тем, капсидные белки представляют собой диполи, то есть они уложены в капсиде таким образом, что отрицательно заряженные радикалы аминокислот обращены внутрь капсида, а положительно заряженные радикалы – наружу. Такая ориентация молекул капсидных белков обусловлена преимущественно положительным зарядом вирусной ДНК или РНК.
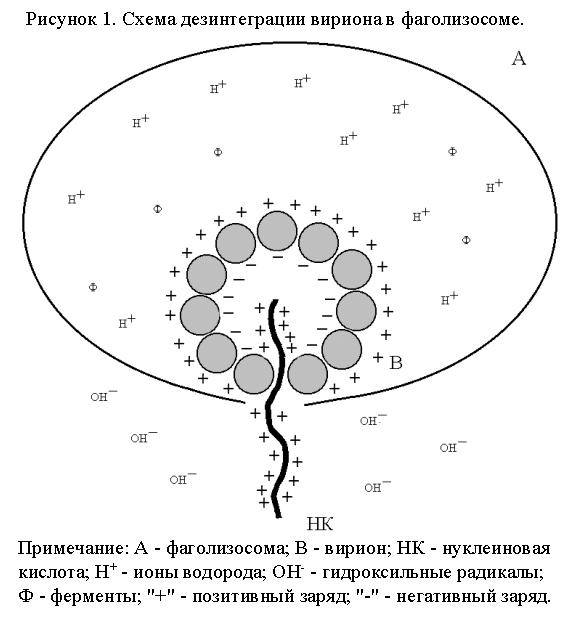 На
вирион, находящийся в фаголизосоме,
действуют 2 электростатических поля:
наружное и внутреннее. Наружное
электростатическое поле обусловлено
положительно заряженным содержимым
фаголизосомы и отрицательно заряженным
содержимым цитоплазмы. Поскольку
наружная поверхность капсида вириона
имеет положительный заряд, то вирион
под действием наружного электростатического
поля движется к мембране фаголизосомы
до тесного контакта
с ней, если до этого рецепторы, удерживающие
вирион в фагосоме, не были разрушены
лизосомальными протеазами. Тесный
контакт вириона с мембраной необходим
для обеспечения бездефектного выхода
геномной ДНК или РНК из вириона в
цитоплазму. Если выход геномной
нуклеиновой кислоты произойдет в жидкой
среде фаголизосомы, то это приведет к
инактивации вирусного генома. Например,
у пикорнавирусов
даже единичный разрыв в первичной
структуре РНК ведет к ее полной
инактивации.
На
вирион, находящийся в фаголизосоме,
действуют 2 электростатических поля:
наружное и внутреннее. Наружное
электростатическое поле обусловлено
положительно заряженным содержимым
фаголизосомы и отрицательно заряженным
содержимым цитоплазмы. Поскольку
наружная поверхность капсида вириона
имеет положительный заряд, то вирион
под действием наружного электростатического
поля движется к мембране фаголизосомы
до тесного контакта
с ней, если до этого рецепторы, удерживающие
вирион в фагосоме, не были разрушены
лизосомальными протеазами. Тесный
контакт вириона с мембраной необходим
для обеспечения бездефектного выхода
геномной ДНК или РНК из вириона в
цитоплазму. Если выход геномной
нуклеиновой кислоты произойдет в жидкой
среде фаголизосомы, то это приведет к
инактивации вирусного генома. Например,
у пикорнавирусов
даже единичный разрыв в первичной
структуре РНК ведет к ее полной
инактивации.
Первичное разрушение капсида вызывает внутреннее электростатическое поле, которое создается между положительно заряженным содержимым фаголизосомы и отрицательно заряженными полюсами капсидных белков. Положительный электростатический потенциал содержимого фаголизосомы значительно превышает таковой для отрицательно заряженных полюсов капсидных белков, в связи с чем белковые молекулы ротируются навстречу положительному потенциалу. Это приводит к деформации капсида, разрыву межкапсидных связей и образованию пор. Через образовавшиеся поры капсида из середины вириона экспонируются гидрофобные остатки белков капсида, что приводит к его слиянию с мембраной фаголизосомы. В мембране образуется пора, через которую положительно заряженная ДНК или РНК, по закону притяжения разноименных зарядов, попадает в отрицательно заряженную цитоплазму. Оставшийся в фаголизосоме капсид подвергается в последующем полному ферментативному гидролизу.
Дезинтеграция вирионов, проникших в клетку путем слияния мембран. Вирусы, размножающиеся в цитоплазме клетки, могут «раздеваться» 2 способами. Первый способ (парамиксо-, бунья-, арено-, реовирусы и др.) заключается в слиянии вириона с клеточной лизосомой, внутри которой дезинтеграция идет по механизму, описанному ранее для вирусов, проникающих в клетку путем виропексиса. Второй способ характерен для вирусов, имеющих внутри капсида ферменты эндонуклеазы, которые разрушают капсид после попадания вириона в цитоплазму клетки. Вирусы, размножающиеся в ядре клетки, из цитоплазмы могут быть транспортированы по каналам эндоплазматической сети специальными белками внутрь ядра. По сравнению с цитоплазмой (рН=7,2), реакция среды кариоплазмы (рН=7,12) менее щелочная, что обусловлено присутствием большого количества ДНК и РНК. Положительный заряд нуклеиновых кислот обусловливает наличие электростатического поля, которое вызывает ротацию отрицательно заряженных полюсов капсидных белков на 180 ˚. Это сопровождается образованием пор в капсиде, через которые вирусные ДНК или РНК выходят в кариоплазму. В образовании пор в капсиде у некоторых вирусов (ВИЧ, ВГВ) могут участвовать внутрикапсидные ферменты – эндонуклеазы.
«Раздевание» вирусов, размножающихся в ядре, может проходить и в цитоплазме клетки при захвате вирионов лизосомами или при активации внутрикапсидных эндонуклеаз. В этом случае в ядро клетки транспортируется вирусная ДНК или РНК.
4-я стадия – синтез вирусных белков и нуклеиновых кислот. Проникнув в клетку, вирусный геном блокирует синтез клеточных белков и запускает синтез вирусоспецифических молекул. Существует 4 пути подавления трансляции клетки-хозяина с помощью вирусной инфекции: 1. Ингибирование клеточных ферментов и иРНК путем их протеолитической или ковалентной модификации. 2. Селективное разрушение регуляторных белков клетки. 3. Нарушение ионного состава внутри клетки таким образом, чтобы преобладала трансляция вируса, а не клетки. 4. Конкурентное использование клеточных рибосом путем образования коротких иРНК (ортомиксовирусы) или путем использования вирусом собственных геномов в качестве иРНК (пикорнавирусы).
Реализация вирусной генетической информации в клетке-хозяине включает 3 процесса: 1. Транскрипцию – многократное копирование вирусной генетической информации в виде информационной РНК (иРНК). 2. Трансляцию – перевод вирусной генетической информации из иРНК в последовательность аминокислотных остатков в строящейся полипептидной цепи белка. 3. Репликацию – образование дочерних вирусных геномов, которые являются точной копией родительских.
Транскрипция. В зависимости от свойств генома может происходить как в ядре, так и в цитоплазме клетки-хозяина, при участии только клеточных или вирусных ферментов – транскриптаз/полимераз, или при их взаимодействии (табл. 1).
Таблица 1