

21.укажите роль пигментов фикобилинов в процессе фотосинтеза? Фитобиллины – к группе , 90 процентов энергиисвета поглащ-ся ими. Осн.-ая роль: доп.пигмент для улавливания света (оранжевие синие желтые спектры) 22. Перечислите компоненты электротранспортной цепи фотосинтеза.
Фотосистема 1. В его состав в качестве реакционного центра входит молекула хлорофилла с максимумом поглощения 700 нм, а также молекула хлорофилла с длинной волны 675-695нм.
Первичным акцептором электронов является молекула хлорофилла с длинной волны 695нм. Вторичными акцепторами являются железосерные белки А2 и А3. Под действием света в фотосистеме 1 восстанавливается водорастворимый железосерный белок – ферредоксин (Ф8), а окисляется медьсодержащий водорастворимый белок – пластоцианин (Пц)
Фотосистема 2 включает в себя реакционный центр, куда входят молекула хлорофилла, поглощ. Светом с длинной волны Р680 нм. В качестве первичных акцепторов выступает предшественник хлорофилла – молекула феофетина. В качестве вторичных акцепторов выступает пластохинон PQ и окислитель молекулы воды. Также в состав фотосистемы 2 входит белковый комплекс S-системы, представляющий собой систему окисляющую воду и переносчик электронов Z системы, которая служит донором электронов для Р680.
23. Опишите процесс циклического транспорта электронов.
Циклический транспорт электронов(ЦТЭ) – путь этапного переноса электронов Р700 через ряд промежуточных переносчиков. В этом процессе принимает участие только Фотосистема 1 и образуется только молекула АТФ. Данный транспорт электронов характерен для фотосинтезирующих бактерий и эукариотических организмов, испытывающие действие негативных факторов внешней среды. Открыта в 1954г.
циклический транспорт электронов это процесс, в котором фотосистема I под действием света катализирует окисление пластоцианина на люменальной поверхности мембраны тилакоида и восстановление ферредоксина на стромальной стороне мембраны. Дальнейшее окисление ферредоксина и восстановление пула пластохинонов является участком, не совпадающим с линейным транспортом.
24. Опишите процесс нециклического транспорта электронов
НТЭ включает 2 фотосистемы, образующие 2 энергетические молекулы NADPH и АТФ. Происходит выделение воды с выделением кислорода. Электрон с ФС1 используется для синтеза NADPН, электроны с ФС2 идет на заполнение дырок ФС1, заполняется за счет электронной, образовавшихся в процессе фотолиза воды.
нециклический транспорт электронов это перенос электронов от воды к НАДФ+, осуществляемый с участием двух фотосистем, цитохромного b6 f-ком¬плекса и промежуточных низкомолекулярных переносчиков — пула пластохинонов (PQ), пластоцианина (Пц), ферредоксина (ФД) и ферредоксин-НАДФ-оксидоредуктазы (ФНР). Он сопровождается выделением кислорода и восста¬новлением НАДФ+. Сопряжен с синтезом АТФ (нециклическое фотофосфорилирование).
25. Перечислите этапы фотолиза воды
ФОТОЛИЗ ВОДЫ — расщепление молекулы воды, в частности в процессе фотосинтеза. Вследствие Ф. в. образуется кислород, выделяющийся зелеными растениями на свету. Для нормального протекания процесса фотолиза воды необходимо полноценное минеральное питание и доступность подвиж. Воды
26. Опишите механизм фотофосфолирования
Это присоединение фосфатов с выделением АТФ. Энергия поглащения хлорофиллом при переносе электрона от одного переносчика к другому обеспечивает преобразование в энергию химических связей. Таким образом фотофосфолирование это синтез АТф протекающий с использованием видимой части солнечной радиации. Впервые предложил эту теорию Митчел.
27. Этапы с1-путь фотосинтеза. Цикл Кальвина
С3-путь фотосинтеза или цикл Кальвина
Этот путь ассимиляции СО2 обнаружен у всех фотосинтезирующих растений. Он был назван циклом Кальвина в честь американского биохимика М. Кальвина, который с сотрудниками открыл и изучил его в 1946-1956 годах с помощью метода меченых атомов и хроматографии. Растения разное время подкармливали 14СО2 и хроматографировали экстракты. Радиоактивные изотопы по химическим свойствам не отличаются от стабильных. Принимая участие в реакциях, они помечают те соединения, в которые входят. Цикл состоит из трех этапов: карбоксилирования, восстановления, регенерации первичного акцептора СО2 и синтеза конечного продукта фотосинтеза (рис. 5.5).
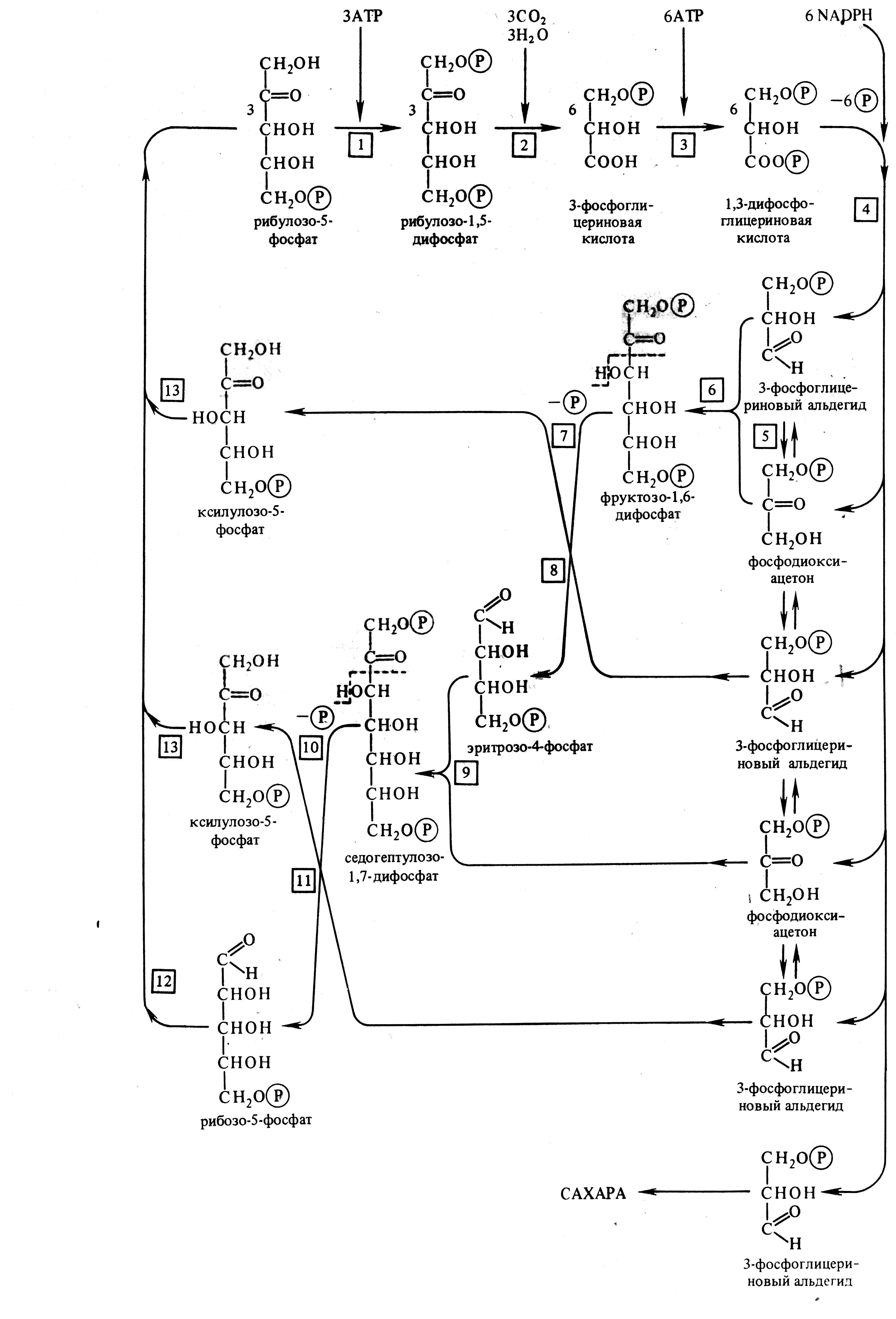
Рис. 5.5. Цикл Кальвина (С3-путь фотосинтеза.
1 – фосфорибулокиназа, 2 – рибулозодифосфаткарбоксилаза, 3 - фофсфоглицераткиназа, 4 – триозофосфатдегидрогеназа, 5 – триозофосфатизомераза, 6 – альдолаза, 7 – фосфатаза, 8 – транскетолаза, 9 – альдолаза, 10 – фосфатаза, 11 – транскетолаза, 12 – рибозофосфатизомераза, 13 – фосфокетопентоэпимераза (по В. В. Полевому).
1. Карбоксилирование. Фосфорибулокиназа фосфорилирует при участии АТФ рибулозо-5-фосфат с образованием АДФ и рибулозо-1,5-дифосфата. Последний является акцептором СО2 и под действием рибулозодифосфаткарбоксилазы присоединяет СО2. В результате образуются 2 молекулы 3-фосфоглицериновой кислоты (3-ФГК).
2. Восстановление. Фосфоглицераткиназа при участии АТФ фосфорилирует 3-ФГК и образовавшаяся 1,3-дифосфоглицериновая кислота восстанавливается с помощью НАДФН и дегидрогеназы фосфоглицеринового альдегида до 3-фосфоглицеринового альдегида (3-ФГА).
3. Регенерация. После фиксации трех молекул СО2 и образования шести молекул 3-ФГА пять из них используются для синтеза рибулозо-5-фосфата, а одна молекула 3-ФГА – для образования глюкозы.
Триозофосфатизомераза превращает 3-ФГА в фосфодиоксиацетон. Затем альдолаза образует из 3-ФГА и фосфодиоксиацетона фруктозо-1,6-дифосфат. Он теряет один остаток фосфорной кислоты под влиянием фруктозо-1,6-дифосфатазы и превращается во фруктозо-6-фосфат. Транскетолаза переносит гликолевый альдегид от фруктозо-6-фосфата на 3-ФГА с образованием эритрозо-4-фосфата и ксилулозо-5-фосфата. Альдолаза присоединяет фосфодиоксиацетон к эритрозо-4-фосфату с образованием седогептулозо-1,7-дифосфата. Он дефосфорилируется фосфатазой и под влиянием транскетолазы соединяется с 3-ФГА. Продуктами этой реакции являются ксилулозо-5-фосфат и рибозо-5-фосфат. Две молекулы ксилулозо-5-фосфата при участии рибулозофосфатэпимеразы и одна молекула рибозо-5-фосфата под действием рибозофосфатизомеразы превращаются в три молекулы рибулозо-5-фосфата.
Шестая молекула 3-ФГА используется для синтеза фруктозо-1,6-дифосфата при повторении цикла. Из двух молекул фруктозо-1,6-дифосфата образуются фруктозо-6-фосфат и глюкозо-1-фосфат. Последний, взаимодействуя с уридинтрифосфатом, дает уридинфосфоглюкозу. Она и фруктозо-6-фосфат образуют сахарозофосфат, который после дефосфорилирования превращается в сахарозу. Следовательно, для образования одной молекулы сахарозы необходимо прохождение четырех циклов Кальвина. Крахмал синтезируется из уридинфосфоглюкозы под влиянием амилосинтетазы.
Среди продуктов фотосинтеза обнаружены аминокислоты. При недостатке НАДФН 3-ФГК превращается не в 3-ФГА, а в пировиноградную кислоту. Она, присоединяя аммиак, образует аланин. Из пировиноградной кислоты в цикле Кребса образуются органические кислоты, которые в ходе реакций аминирования и переаминирования дают аминокислоты.
28. С4-путь. Цикл Хейча-Стейлера
Австралийскими учеными М. Хетчем и К. Слэком был описан С4-путь фотосинтеза, характерный для тропических и субтропических растений (сахарный тростник, кукуруза), ряд сорных растений: ежовник, просо, щетинник. Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по С4-пути, имеются два типа клеток и хлоропластов: 1) мелкие гранальные пластиды в клетках мезофилла листа;
2) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки. Имеются обкладочные клетки: в них мало хлоропластов, но много крахмальных зерен. Клетки обкладки имеют утолщенные клеточные стенки, содержат большое количество хлоропластов и митохондрий, расположены вокруг сосудистых пучков в 1 или 2 слоя. Совокупность указанных особенностей анатомического строения получила название корончатой анатомии или корончатого. Хлоропласты разных типов клеток характеризуются не только особенностями строения, но и разным типам фосфорилирования. В клетках мезофилла по преимуществу происходит нециклическое фосфорилирование и образуется НАДФН, необходимый для цикла Кальвина, идущего в клетках обкладки. В хлоропластах клеток обкладки идет только циклическое фосфорилирование. Такое разделение типов фосфорилирования, возможно, связано с тем, что к хлоропластам клеток обкладки, расположенным в глубине листа, проникает по преимуществу более длинноволновый свет, который не поглощается фотосистемой, ответственной за разложение Н20.
На первом этапе С4-пути углекислый газ, диффундирующий в лист через устьица, попадает в цитоплазму клеток мезофилла с мелкими хлоропластами, в которых и происходит реакция карбоксилирования фосфоенолпировиноградной кислоты (феп). Образуется щавелевоуксусная кислота (оксалоацетат). ЩУК преобразуется в яблочную кислоту (малат) или аспарагиновую (аспартат). Восстановление до маната происходит в присутствии НАДФН, а для образования аспартата необходимо наличие NH4+. Затем яблочная (или аспарагиновая) кислота, по-видимому, по плазмодесмам передвигается в клетки обкладки. В клетках обкладки яблочная кислота декарбоксилируется ферментом малатдегидрогеназой до пировиноградной кислоты (пируват) и С02. Реакция декарбоксилирования может варьировать у разных групп растений с использованием разных ферментов. С02 поступает в хлоропласты клеток обкладки и включается в цикл Кальвина—присоединяется к РДФ(рибулозодифосфат) (В дальнейшем РДФ превращается в 2(3ФГК). ФГК - -3фосфоглицериновая кислота). Пируват возвращается в клетки мезофилла и превращается в первичный акцептор С02 — ФЕП. Таким образом, при С4-пути реакция карбоксилирования происходит дважды. Это позволяет растению создавать запасы углерода в клетках. Акцепторы С02 (ФЕП и РБФ) регенерируют, что и создает возможность непрерывного функционирования циклов. Фиксация С02 с участием ФЕП и образованием малата или аспартата служит своеобразным насосом для поставки С02 в хлоропласты обкладки, функционирующих по С3-пути. Поскольку при таком механизме фотосинтеза принимают участие два типа клеток и два типа хлоропластов, этот путь называют еще кооперативным.
«+»:
-С4 более полно усваивает СО2. С4 растения накапливают углекислый газ в органических кислотах (в малате), для осуществления фотосинтеза даже в закрытых устьицах. С4-путь наиболее эффективно использует воду. С4 может фиксировать углекислый газ при более низких ее концентрациях.
P.S. В клетках мезофилла происходит последовательное превращение АТФ (аденозинтрифосват)-АДФ(ди) –АМФ (моно). (по лекции)