

34. Механизм образования пептидных связей .Пептидилтрансферазная активность рибосомы.
Образование пептидной связи происходит сразу же после отщепления комплекса EF-1 и ГДФ от рибосомы. Эта стадия процесса получила название реакции транспептидации (рис. 4-39).
В ходе этой реакции остаток метионина Мет-тРНКIМет связывается с a-аминогругшой первой аминокислоты, присоединённой к тРНКaa1 и расположенной в А-центре, образуется первая пептидная связь. Установлено, что пептидилтрансферазная активность большой субъединицы рибосомы принадлежит 28S рРНК. К настоящему времени обнаружена целая группа РНК, обладающая
176
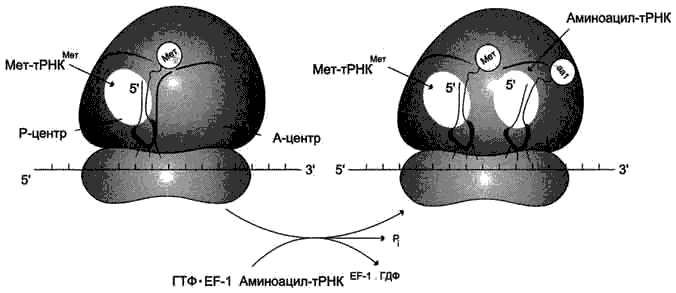
Рис. 4-38. Включение аа1-тРНКaa1 в рибосому. aа1-тРНКaa1 взаимодействует с рибосомой в виде тройного комплекса, состоящего из фактора элонгации EF-1I, аа1-тРНKaa1 и ГТФ. Антикодон аа-тРНКаа1 комплементарен и антипараллелен кодону мРНК в А-центре. Связывание аа1-тРНКaa1 происходит за счёт энергии гидролиза ГТФ до ГДФ и Рi
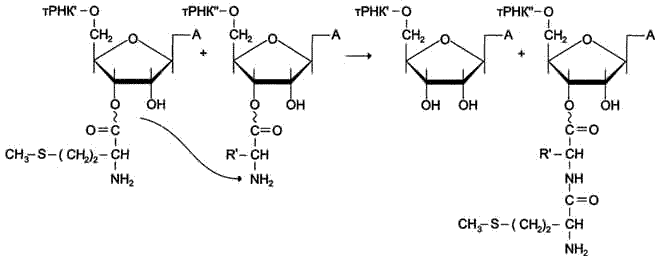
Рис. 4-39. Реакция транспептидации. Метионин от Мет-тРНКiМет, находящегося в Р-центре, присоединяется к α-МН2 -группе аминоацильного остатка аа1-тРНКaa1 А-центра с образованием новой пептидной связи.
свойствами ферментов. Эти каталитически активные РНК получили название рибозимов (см. раздел 2). Полагают, что рибозимы можно считать "реликтами" раннего периода эволюции, когда белки ещё не приобрели такого значения, как в последующие периоды.
35.Инициация трансляции и её механизм. Факторы инициации.
Механизмы трансляции прокариот и эукариот существенно отличаются, поэтому многие вещества, подавляющие прокариотическую трансляцию, в значительно меньшей степени действуют на трансляцию высших организмов, что позволяет использовать их в медицинской практике как антибактериальные средства безопасные для организма млекопитающих.
Процесс трансляции разделяют на
§ инициацию — узнавание рибосомой стартового кодона и начало синтеза.
§ элонгацию — собственно синтез белка.
§ терминацию — узнавание терминирующего кодона (стоп-кодона) и отделение продукта.
Синтез белка в большинстве случаев начинается с AUG-кодона, кодирующего метионин. Этот кодон обычно называют стартовым или инициаторным. Инициация трансляции предусматривает узнавание рибосомой этого кодона и привлечение инициаторной аминоацил-тРНК. Для инициации трансляции необходимо также наличие определённых нуклеотидных последовательностей в районе стартового кодона. Немаловажная роль в защите 5'-конца мРНК принадлежит 5'-кэпу. Существование последовательности, отличающей стартовый AUG от внутренних совершенно необходимо, так как в противном случае инициация синтеза белка происходила бы хаотично на всех AUG-кодонах.
Процесс инициации обеспечивается специальными белками — факторами инициации (англ. initiation factors, сокращённо IF; эукариотические инициаторные факторы обозначают eIF, от англ. eukaryotes).
Механизмы инициации трансляции у про- и эукариот существенно отличаются: прокариотические рибосомы потенциально способны находить стартовый AUG и инициировать синтез на любых участках мРНК, в то время как эукариотические рибосомы обычно присоединяются к мРНК в области кэпа и сканируют её в поисках стартового кодона.
Большинство молекулярных механизмов, осуществляющих регуляцию экспрессии генов на уровне трансляции, реализуется на стадии инициации биосинтеза белка. Помимо субъединиц эукариотических рибосом и белков, обычно ассоциированных с 5'- и 3'-концевыми последовательностями мРНК, в инициации принимают участие по меньшей мере 11 белковых факторов, построенных более чем из 25 полипептидов (табл. I.11 ).
Фактороы, необходимых для инициации трансляции эукариотических мРНК эукариотическими рибосомами и обозначаются как eIF-1, eIF-2, eIF-3, eIF-4A, eIF-4B, eIF-4C, eIF-4D, eIF-4E, eIF-4F, eIF-5 и т.д.
eIF-2 , eIF-3 и eIF-5 строго необходимы для инициации, в то время как eIF-1 , eIF-2B , eIF-4A ,eIF-4B , eIF-4C , eIF-4E , eIF- 4F оказывают большее или меньшее стимулирующее действие. Роль и вклад eIF-2A и eIF-4D в инициацию вообще не ясна, и имеются сомнения, являются ли он действительно необходимыми факторами инициации ( Спирин А.С., 1986 ). В экспериментах in vitro белок eIF-2A стимулирует связывание инициаторной тРНК с 40S субчастицей в отсутствиеГТФ ; белок eIF-4D стимулирует реакцию инициаторной метионил-тРНК с пуромицином в составе конечного инициаторного 80S-комплекса .
Все факторы инициации, кроме eIF-2A и eIF-4D, по- видимому, РНК-связывающие белки в том смысле, что имеют неспецифическое сродство к любой высокомолекулярной РНК и способны образовывать с ней более или менее стабильные комплексы