

Глава 2 Потенциал действия в кардиомиоцитах
Потенциал действия кардиомиоцитов состоит из 5 фаз (рис. 10): фазы 0 (быстрая деполяризация), фазы 1 (ранняя быстрая реполяризация), фазы 2 (плато), фазы 3 (завершающая быстрая реполяризация), фазы 4 (потенциал покоя и диастолическая деполяризация).
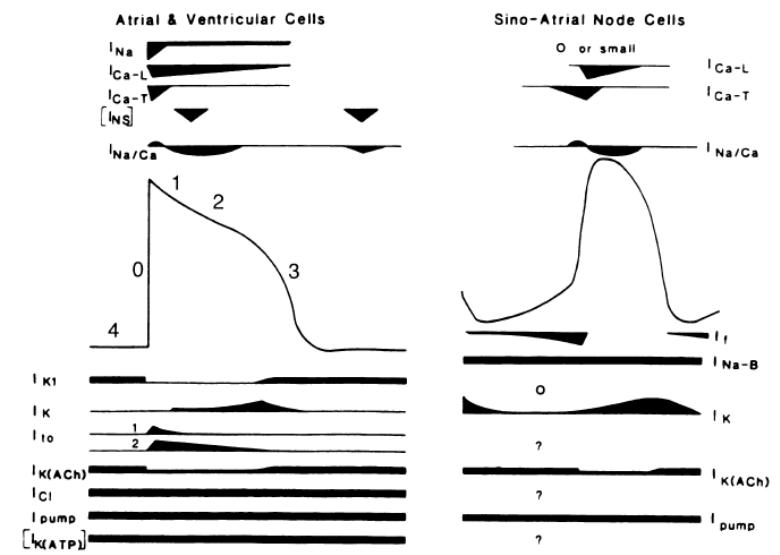
Рис. 10. Токи и каналы, вовлеченные в механизмы формирования потенциала покоя и потенциала действия кардиомиоцита (по H. T. Shih [86])
Амплитуда и продолжительность ПД различается в зависимости от локализации в сердце; это различие обусловлено качественными и количественными различиями в ионных каналах [86]. На рис. 10 показаны токи, вовлеченные в формирование ПП и ПД кардиомиоцита.
Потенциал покоя кардиомиоцитов варьирует в пределах от -50 до -95 мВ и обусловлен распределением ионов Na+, Ca2+, K+ и Cl- по обе стороны мембраны. Наибольший вклад в значение потенциала вносит K+. В состоянии покоя мембрана обладает довольно высокой проницаемостью для ионов K+, а значение мембранного потенциала близко к Ек (электрохимическому мембранному поьенциалу равновесному для K+). Проницаемость мембраны для K+ зависит от состояния калиевых каналов Iк1, активность которых возрастает в период реполяризации и гиперполяризации. Большая проницаемость мембраны для входящих токов которая характерна для значений МП ниже Ек препятствует интенсивной гиперполяризации. Через каналы Iк1 при небольшой деполяризации течет исходящий ток; оба этих процесса и являются причиной того, что ПП близок к Ек. каналы Iк1 обнаружены в предсердии, АВ-узле, пучке Гиса и волокнах Пуркинье, клетках миокарда желудочков, клетках SA. Другими механизмами, поддерживающими градиент концентраций Na+ и K+ является Na+/K+ насос и Na+, Ca2+ обменный механизм. Обнаружено несколько модификаторов потенциала покоя, в т.ч. АТФ – зависимые K+ каналы Iк(АТР) и ацетилхолинзависимые K+ каналы (Iк(Ach)). Iк(АТР) каналы закрыты при физиологических концентрациях внутриклеточного АТР. При снижении концентрации АТР до значений ниже 1мМ или при возникновении гипоксии эти каналы активируются, возникает исходящий ток и, как следствие – гиперполяризация. Эти каналы в СА не обнаружены. Iк(Ach) – это калиевый канал активируемый мускариновым рецептором посредством G-белка. Связывание Ах с рецептором приводит к гиперполяризации [86].
2.1. Фазы потенциала действия
Фаза «0» - быстрая деполяризация. Деполяризация в клетках предсердия и желудочков, а также клетках волокон Пуркинье и пучка Гиса обусловлена активацией потенциал-зависимых Na+ каналов. В клетках СА и АВ также были описаны эти каналы, однако они не играют главную роль в процессах деполяризации: здесь наибольшее значение имеют входящие Ca2+ токи. Активация натриевых каналов и вход Na+ обуславливают смещение мембранного потенциала к уровню ионного равновесного потенциала для Na+ (ЕNa) [86]. Транзиторное повышение входящего тока Na+ продолжается в течение 1-2 мсек. По мере возрастания концентрации Na+ внутри клетки и положительного заряда. Фаза 0 потенциала действия в СА и АВ медленнее, чем в клетках желудочков. Это обусловлено тем, что основной вклад в деполяризацию здесь вносят входящие токи Ca+. В сердце имеется два вида каналов: Т-типа (ICaT) и L-типа (ICaL). В клетках сердца наибольшее функциональное значение имеют каналы ICaL. Именно эти каналы, а также проходящие через них токи, обуславливают деполяризацию в клетках СА и АВ.
Фаза «1» - Ранняя быстрая реполяризация. После фазы 0 мембранный потенциал быстро и транзиторно (обратимо) возвращается к нулевому уровню. Этот процесс обусловлен инактивацией натриевых и кальциевых каналов, количество различных видов калиевых каналов очень велико: калиевые каналы потенциал-зависимые (Ito,t, Ito,s – транзиторные быстрые (t) и медленные (s), IKr и IKs - быстрые и медленные токи, IKur – очень быстрый ток и т.д.) и функционирующие вне зависимости от мембранного потенциала (IK1 – фоновый калиевый ток) (рис. 1 – Rudy; таблица 1 – Norbonne). Основную роль в фазе 1 играет быстрый транзиторный ток калия (Itof ), а также Itos .
Фаза «2» - Плато. Фаза 2 может продолжаться несколько сот миллисекунд. Деполяризация мембраны в фазу 0 активирует потенциал-зависимые кальциевые каналы; возникающий вследствие этого ток кальция через L-каналы ICaL и обуславливает плато. В течение фазы 2 снижается проводимость мембраны для большинства ионов, кроме Ca2+: натриевые каналы инактивированы, проводимость для К+ снижена; имеет место небольшой реполяризующий ток Cl- и токи, обусловленные функционированием Na+/К+ насоса.
Фаза «3» - Окончательная быстрая реполяризация. Эта фаза обусловлена инактивацией входящих токов катионов и активацией IKr и IKs [86].
Для кардиомиоцита в фазу плато и фазу деполяризации характерна рефрактерность. Рефрактерность принимает две формы: абсолютную и относительную. Согласно современным представлениям, рефрактерность обусловлена инактивацией натриевых каналов. Рефрактерность обеспечивает угасание волны возбуждения после того, как она охватит полностью весь миокард; таким образом, обратный вход этой волны невозможен, т.к. сердце находится в состоянии рефрактерности [2].
Электрофизиологические процессы, происходящие в пучках волокон Пуркинье, характеризуются рядом выраженных особенностей. Клетки в пучках волокон Пуркинье также обладают пейсмекерной активностью, однако она развивается значительно медленнее, чем в СА-узле. Выше отмечалось, что основной функцией волокон Пуркинье является проведение возбуждения от СА и АВ-узлов к клеткам миокарда желудочков. Для описания сложных процессов, которые при этом протекают, используется кабельная теория, описание которой, в целом, выходит за рамки настоящей работы. Здесь мы частично рассмотрим лишь шестнадцатый постулат и его практические реализации. Проводящая система желудочков состоит из сильно ветвящейся сети клеток, расположенных субэндокардиально. Отдельные клетки этой сети связаны друг с другом посредством вставочных дисков, глубинно внедряющихся в продольно ориентированные разрастания сарколеммы. Вставочные диски имеют четыре структурные области («промежуточные контакты»; десмосомы; межклеточные контакты или нексусы; неспециализированная сарколемма). Считается, что входящие в состав межклеточных дисков контакты обеспечивают электрическое взаимодействие между клетками. Согласно современным представлениям клетки волокон Пуркинье относятся к тканям с быстрым ответом (для таких тканей характерен входящий ток ионов Na+ в период ПД). Согласно 16-му постулату кабельной теории «влияние локальных токов на возбудимость зависит от интенсивности источника и характеристик тока; если выполнены требования, связанные с предельной длиной, возникает регенеративный распространяющийся ПД, если требования не выполнены, то наблюдается местный электрический ответ». С физиологической точки зрения это означает следующее. При пороговом значении мембранного потенциала в клетку начинают поступать ионы Na+ , которые заряжают внутренность клетки положительно по отношению к её окружению. Происходит деполяризация и в миоплазме образуется движущая сила, вызывающая возникновение тока, проходящего в продольном направлении. Основными источниками этого тока внутри клеток и между клетками являются ионы K+ . Ток, текущий в продольном направлении, вытесняет отрицательные заряды с внутренней поверхности окружающих участков мембраны, вызывая деполяризацию этих участков до уровня порогового потенциала. Цепь замыкается емкостным током, проходящим через мембрану и током внеклеточного пространства [2].
На рисунке 11 представлены типичные варианты потенциала действия в различных отделах сердца.
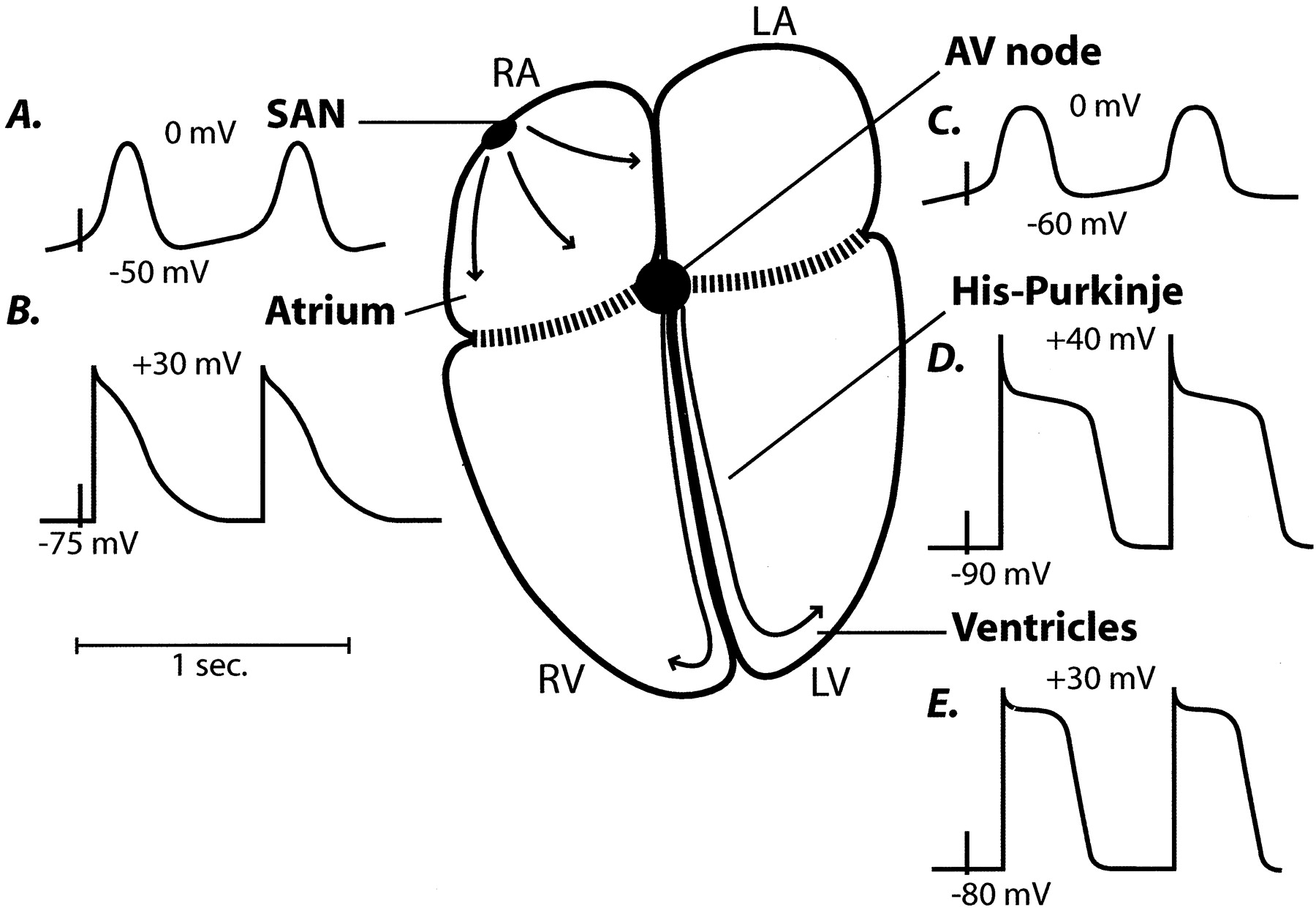
Рис. 11. Варианты потенциала действия в различных отделах сердца (по G. Schram [79])
Примечание: SAN – синатриальный узел, AV node – атриовентрикулярный узел, His-Purkinje – пучок Гиса и волокна Пуркинье, Ventricles – кардиомиоциты желудочка.
В норме потенциал действия генерируется в синатриальном узле (САУ) и проводится через предсердие к атриовентрикулярному узлу (АВУ). От АВУ ПД движется через пучок Гиса и волокна Пуркинье, достигая кардиомиоцитов желудочка и вызывая сокращение.
На рис. показаны ионные токи, вовлеченные в формирование ПД [79].
В таблице 7 представлены ионные токи, участвующие в формировании ПД кардиомиоцита.