

Сперматогенез у хвостатых амфибий
Семенники амфибий фолликулярно-цистного типа. Образование семенных фолликулов происходит в зачатковой зоне семенника. Клетки фолликулярного эпителия, выстилающие полость семенных фолликулов, образуют цисты, внутри которых в тесном контакте с цитоплазматическими отростками фолликулярных клеток происходит сперматогенез. В составе цист одного фолликула половые клетки развиваются синхронно. В связи с сезонностью размножения количество фолликулов, содержащих половые клетки тех или иных стадий спрематогенеза, на протяжении года закономерно изменяется. Фолликулы разных стадий сперматогенеза расположены в семеннике зонами соответственно последовательности стадий сперматогенеза. Фолликулярные клетки образуют стенку цист на протяжении периодов размножения и роста половых клеток (при этом количество фолликулярных клеток в стенках цист увеличивается в ходе периода размножения). При переходе к спермиогенезу цитоплазма каждой фолликулярной клетки стягивается к ее ядросодержащему участку, одновременно происходит имплантация в нее сперматид. Циста разрушается, и каждая фолликулярная клетка с внедренным в нее пучком сперматид формирует сперматофорную группировку. После спермиации освобожденные от связи со спермиями фолликулярные клетки фагоцитируют остаточные спермии, затем, по всей вероятности, дегенерируют, либо повторно участвуют в образовании семенных фолликулов.
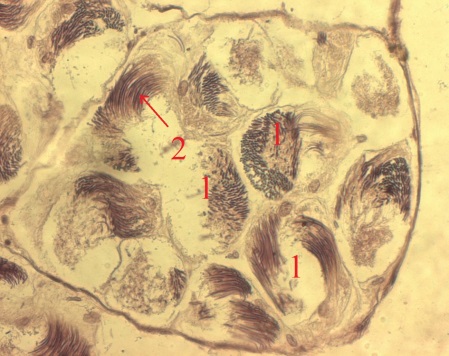
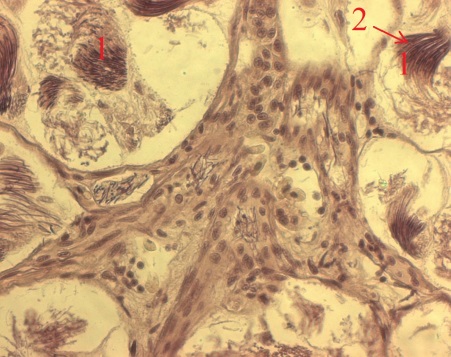
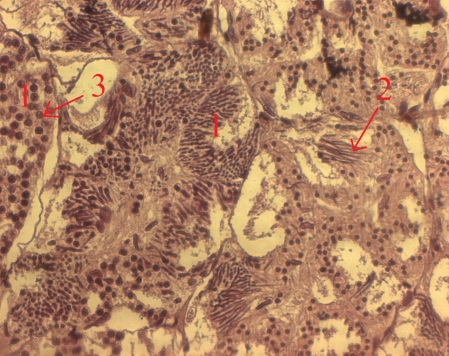
Рис. 4. Срезы семенника тритона (X20) (1 – циста; 2 – сперматозоиды; 3 – сперматиды).
Сперматогенез у бесхвостых амфибий
Бесхвостые амфибии, как и костные рыбы, имеют канальцево-цистный
тип стоения семенников.
костные рыбы, имеют канальцево-цистный
тип стоения семенников.
Сперматогенная зона находится в семенном канальце. Стенка цисты многоклеточная. В фолликулярном эпителии, выстилающем семенные канальцы, расположены сперматогонии. С началом очередной волны сперматогенеза (весна) клетки фолликулярного эпителия формируют цисты, внутри которых осуществляется сперматогенез. В составе половых клеток одной цисты он протекает синхронно. Стенка каждой цисты состоит из фолликулярных клеток, цитоплазматические отростки которых разветвляются между половыми клетками. С началом спермиогенеза (осень) происходит имплантация сперматид в стенку цист и подтягивание цитоплазмы фолликулярных клеток вместе с внедренными сперматидами к базальной мембране канальца, образуются пучки спермиев, внедренных в апикальную цитоплазму фолликулярных клеток. По завершении спермиации фолликулярные клетки фагоцитируют остаточные спермии.
Я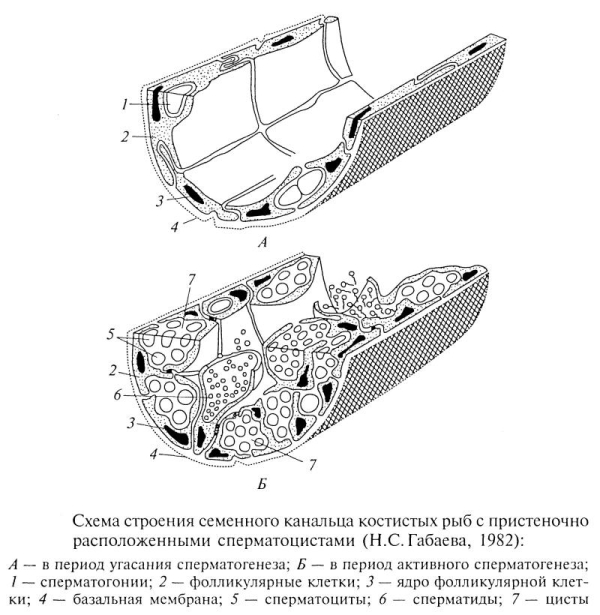 дро
фолликулярной клетки неправильной
формы, лопастное, с крупным ядрышком,
объем ядра увеличивается в период
имплантации сперматид. В цитоплазме
фолликулярных клеток: МТХ, ЭПР
(преимущественно гладкий), рибосомы,
АГ, микрофиламенты, гранулы гликогена,
липидов. Спермиация происходит под
воздействием гонадотропинов.
дро
фолликулярной клетки неправильной
формы, лопастное, с крупным ядрышком,
объем ядра увеличивается в период
имплантации сперматид. В цитоплазме
фолликулярных клеток: МТХ, ЭПР
(преимущественно гладкий), рибосомы,
АГ, микрофиламенты, гранулы гликогена,
липидов. Спермиация происходит под
воздействием гонадотропинов.
Сперматогенный цикл имеет годовую периодичность. Судьба остаточных фолликулярных клеток не вполне ясна. Некоторые исследователи видели у них митозы и полагают возможным их повторное участие в образовании цист. Подобно этому у других видов бесхвостых амфибий строение и цитохимия фолликулярных клеток меняется синхронно со сперматогенезом. Полагают, что клетки фолликулярного эпителия участвуют в синтезе стероидов и регуляции сперматогенеза.
Существует три типа сперматогенеза.
Прерывный тип характеризуется тем, что сперматогенез либо нарушен, либо полностью прерван во время суровых погодных условий, а иногда даже летом (рис. 1(а)). Яичко характеризуется наличием ниши стволовых клеток и дегенерацией зрелых клеток в период покоя, например, у Bufo japonicus. В период покоя яичко рефрактерно к любому стимулирующему фактору, гормональному или экологическому, но на сегодняшний день только два вида из исследованных обладают прерывным циклом сперматогенеза - это европейская красная лягушка Rana temporaria и большая четырехглазая лягушка из южной Аргентины, Pleurodema bufonia. Изменение массы яичек является показателем сезонного репродуктивного статуса и имеет максимальное значение в начале сезона размножения с последующим минимумом в конце сезона, когда семенные канальцы полностью опустошены. После спаривания наступает небольшой период рефрактерности, после чего цикл сперматогенеза повторяется.
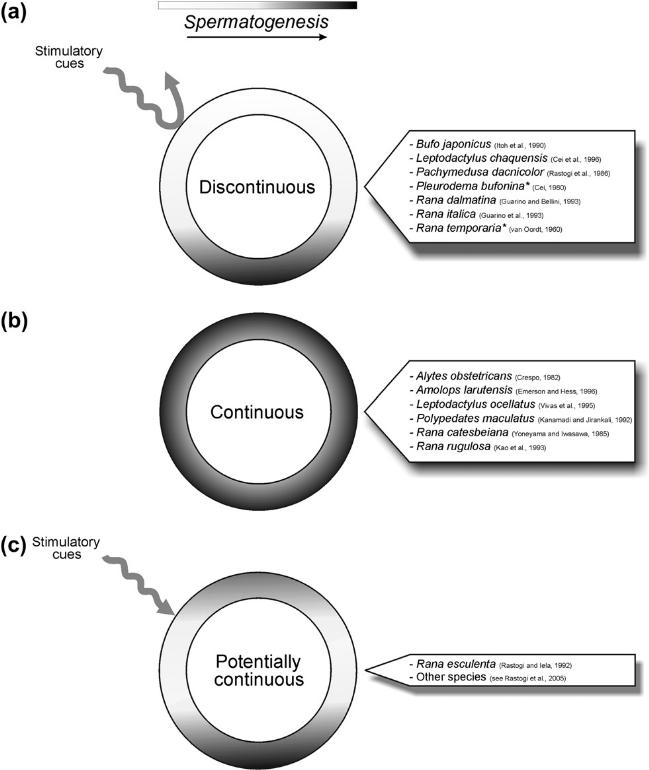
Рисунок 1. Generalized diagram of the three types of spermatogenetic cycle displayed in anuran species. In ‘stimulatory cues’ are included some endogenous factors such as hormones and environmental factors such as temperature, humidity, photoperiod, food availability, and rainfall. For each type of spermatogenetic cycle (A, B, C), examples are reported in the label on the right. * Discontinuous sensu stricto: spermatogenesis cannot be stimulated by any means during the seasonal quiescence.
Непрерывный тип сперматогенеза (рис.1 (b)), наоборот, характеризуется протеканием различных стадий сперматогенеза в течение года, т.е. воспроизводство не носит сезонный характер (например, жаба-акушерка, Alytesobstetricans. Однако, могут возникать некоторые сезонные колебания относительной частоты образования половых клеток, включающие мейотические и / или спермиогенные этапы. У таких видов более или менее непрерывное производство мужских гамет в течение большей части года.
Третий тип сперматогенеза, характерный для некоторых видов бесхвостых амфибий, определяется как потенциально непрерывный (рис. 1(с)). Он встречается у среднеевропейской зеленой или съедобной лягушки, R. esculenta. Сперматогенез происходит строго сезонно, с умеренным или сильным подавлением образования гамет в холодные месяцы года, когда может происходить массовая дегенерация мейотических клеток, в частности, первичных сперматоцитов.
Различия между прерывным и потенциально непрерывным типами сперматогенеза основаны на том, что в первом случае наблюдается абсолютная рефрактерность сперматогенных тканей к любым стимулам, тогда как во втором случае можно получить ответ на стимулирующее воздействие (температурное или гормональное) в любое время на протяжении зимы.
У видов, имеющих прерывный тип сперматогенеза, изменение массы яичек является показателем сезонного репродуктивного статуса и имеет максимальное значение в начале сезона размножения с последующим минимумом в конце сезона, когда семенные канальцы полностью опустошены. После спаривания наступает небольшой период рефрактерности, после чего цикл сперматогенеза повторяется.
Гормональная регуляция
Самцы с хорошо развитыми мозолями или другими вторичными половыми признаками обычно имеют более высокий уровень андрогенов. Такие самцы готовы к спариванию. Выраженность большинства вторичных половых признаков варьирует в зависимости от сезона, достигая максимального развития в период размножения, что коррелирует с наблюдаемыми флуктуациями уровня циркулирующих андрогенов. В связи с этим рассматривается роль высоких концентраций андрогенов в определении поведения самцов в период размножения. Сезонный цикл изменения концентраций половых стероидов, циркулирующих в крови и выявляемых в яичках, может быль связан с циклом изменения массы яичка или типом сперматогенеза. Тесная связь между изменением массы яичек, сперматогенной активностью и концентрацией андрогнов в плазме крови была найдена у лягушки с прерывным типом сперматогенеза P. dacnicolor (Rastogi et al., 1986, цит. по Rastogi et al., 2011).
Уровень андрогенов в плазме крови широко варьирует не только в период размножения между особей одного вида, но и между особями разных видов. Уровень тестостерона у бесхвостых амфибий различается на порядки: от 595 нг/мл, например, у зрелых самцов Bufo mauritanicus (против 4,4 нг/мл в зимний период), до пикового значения менее1 нг/мл в сезон размножения, например, Dicroglossus occipitalis. Та же закономерность характерна для других изученных андрогенов, таких, как 5а-дигидротестостерон (DHT), D4-андростендион (Rastogi et al., 2005, цит. по Rastogi et al., 2011) или 11-кетотестостерон (Murphy et al., 2006, цит. по Rastogi et al., 2011). Такие различия могут быть связаны с видовыми особенностями плазматических стероид-связывающих белков, а также с методикой, местом и временем отбора проб. Однако, несмотря на межвидовые различия, уровень андрогенов может значительно возрастать в период размножения, в сотни раз превышая фоновое значение, наблюдаемое в течение года.
Андрогены известны как факторы, вовлеченные в регуляцию сперматогенеза. Сезонное повышение уровня циркулирующих гормонов соответствует периоду, в течение которого самцы демонстрируют репродуктивное поведение. Исключение из этой схемы составляют самцы глухой лягушки Rana blythii, у которых низкий уровень андрогенов сохраняется и во время сезона размножения (Emerson & Hess,1996, цит. по Rastogi et al., 2011).
Эстрогены также секретируются в яичках. Наибольшая концентрация 17b-естрадиола в плазме крови – около 4 нг/мл, - была зарегистрирована у R. esculenta. предварительные исследования in vitro показали, что Е2 стимулирует пролиферацию первичных сперматогоний также, как и внедрение тучных клеток, а в то же время имеет тенденцию подавлять интратестикулярный уровень андрогенов у R. Еsculenta (Minucci, Di Matteo, Chieffi, Pierantoni, and Fasano, 1997, цит. по Rastogi et al., 2011).
Зависимость гаметогенного отдела от секреторной активности эндокринного отдела была изучена экспериментально на R. еsculenta. Использование цитотоксического агента (этандиметансульфонат) привело к морфологической и функциональной дезорганизации тканей эндокринного компонента, что отразилось в снижении плазматического и интратестикулярного уровня андрогенов и разрушении сперматогенных тканей (цит. по Rastogi et al., 2011). После остановки цитотоксического воздействия, поврежденные ткани восстановились вместе с уровнем циркулирующих андрогенов, показывая, что некоторые местные, интратестикулярные факторы играют роль в процессе восстановления.
Результаты, полученные в попытках экспериментально продемонстрировать участие тиреоидных гормонов в функционировании гонад, оказались неубедительными (цит. по Rastogi et al., 2011). Тем не менее, последние неопубликованные данные показывают наличие рецепторов к гормонам щитовидной железы, передающие сигнал через TRB-A1 мРНК экспрессию, в яичке лягушки в течение зимних месяцев (неопубликованные наблюдения, цит. по Rastogi et al., 2011).