

Занятие №4. Оогенез амфибий и рыб. Захарченко е. О.
Яйца рыб и амфибий происходят от популяции оогониальных стволовых клеток, способных ежегодно давать новую генерацию ооцитов. То есть период размножения продолжается и у взрослой особи. У лягушки Rana pipiens оогенез длится 3 года. В течение первых двух лет происходит очень медленное увеличение размеров ооцита. В течение 3-го года быстрое накопление желтка приводит к тому, что ооцит приобретает дефинитивные размеры. Созревание ооцитов осуществляется партиями: первая генерация созревает вскоре после метаморфоза, следующая - годом позже. В процессе вителлогенеза у амфибий желток образует желточные пластинки. Большая часть яиц характеризуется высокой степенью асимметрии, и именно во время оогенеза происходит спецификация анимально-вегетативной оси. Концентрация желтка повышается примерно в 10 раз в направлении от анимального к вегетативному полюсу. Включение вителлогенина одинаково по всей поверхности ооцита, а различия в концентрации желтка обусловлены его перемещением внутри ооцита. Если желточные пластинки формируются в будущем анимальном полушарии, то они перемещаются внутрь, к центру клетки. На вегетативном полюсе пластинки не изменяют своего положения, оставаясь в течение длительного времени на периферии и постепенно увеличиваясь в размерах. Они медленно смещаются от кортекса вглубь, а их место занимают новые желточные пластинки, образующиеся у поверхности. В результате этого дифференциального внутриклеточного транспорта в вегетативном полушарии количество желтка возрастает до тех пор, пока здесь не сосредоточится около 75% желтка зрелого ооцита Xenopus. Расположение органелл также становится асимметричным. Из АГ формируются кортикальные гранулы, сначала равномерно по всей цитоплазма, затем – они мигрируют к периферии клетки, располагаясь здесь между пигментными гранулами. К этому времени приурочена репликация МТХ (их образуются миллионы). Ближе к концу вителлогенеза цитоплазма становится стратифицированной (многослойной). Кортикальные гранулы, МТХ и пигментные гранулы обнаруживаются в кортикальном слое ооцита. Во внутренней цитоплазме возникают четкие градиенты. Желточные пластинки все более концентрируются у вегетативного полюса, а гликогеновые гранулы, рибосомы, МТХ и ЭПР – все ближе к анимальному. Наблюдается ооплазматическая сегрегация, хорошо изученная на неравномерном распределении специфических мРНК в цитоплазме ооцита.
У Xenopus стадия лептотены мейоза длится 3-7 суток, зиготены – 5-9 суток, пахитены – около 3 недель. Диплотена может длиться годы. Под действием прогестерона, секретируемого фэ в ответ на гонадотропные гормоны гипофиза, в течение 6 часов происходит дезинтеграция зародышевого пузырька, исчезновение микроворсинок, дезинтеграция ядрышек, сокращение и миграция хромосом типа лщ к анимальному полюсу, инициируется трансляция запасенных мРНК. Вскоре происходит первое деление мейоза, и зрелое яйцо выделяется из яичника в процессе овуляции.
В яичнике амфибий выделяют четко видные лопасти, окруженные выростами оболочки яичника, в каждой лопасти можно наблюдать различные стадии оогенеза.
Оогонии: мелкие клетки с крупным ядром, лежат группами.
Ооциты на стадии малого роста: от малых до средних размеров, с гомогенной цитоплазмой, крупным ядром. В ядре видны многочисленные ядрышки на периферии, хромосомы типа лщ в центре. Иногда хорошо видна кариосфера: хромосомы собраны в центре ядра и окружены капсулой, состоящей из псевдомембран и ассоциированных с ними ядрышек. Изменяется форма ядра: от гладкой до сильно извилистой. Вокруг ооцита – первичная оболочка, пронизанная выростами фолликулярных клеток и ооцита. Фолликулярный эпителий плоский, однослойный, сверху лежит базальная мембрана и один слой клеток теки, включающей сосуды. К концу малого роста около ядра формируется т.н. желточное тело, или тело Бальбиани – скопление АГ и МТХ, ранее считавшееся центром образования желтка. Теперь показано, что хотя желток и может появляться здесь в следствии наличия АГ, к его образованию тело Бальбиани отношения не имеет.
Переход к стадии большого роста – стадия вакуолизации: в периферической области цитоплазмы появляются вакуоли – предшественники кортикальных гранул, вскоре начнут появляться желточные пластинки.
Ооциты на стадии большого роста: ядро сильно изрезанной формы с многочисленными краевыми ядрышками, его диаметр возрастет в 7-8 раз. Между периферией ядра и цитоплазмой идет постоянный интенсивный обмен по механизму «блеббинга» - образования пузырьков на внутренней и наружной ядерных мембранах. В цитоплазме идет активное накопление желтка в виде пластинок, пигмент меланин распределяется в кортикальном слое, преимущественно на анимальном полюсе. Активные синтез и накопление полисахаридов. Фолликулярная оболочка и тека истончаются. Тека иногда не видна вовсе. В период большого роста увеличивается периооцитное пространство, становясь максимальным к овуляции, когда из состава первичной оболочки исчезают отростки фэ и ооцита. Ближе к периоду созревания ядро начинает перемещаться к анимальному полюсу.
In vivo созревание регулируют гонадотропные гормоны. Причем выделяют гормонозависимый период (рецепторы к гонадотропину находятся на фэ, которые под действием этого гормона выделяют прогестерон) и период инерции созревания. Весной в природе действие гонадотропина максимальное, а осенью – минимальное. Созревание возможно стимулировать in vitro с помощью прогестерона.
На протяжении всего оогенеза происходит атрезия (деградация) ооцитов, они теряют форму, накапливают пигмент.
После овуляции оставшиеся клетки фэ и теки образуют своеобразные чехлики, которые видны меду фолликулами на срезах яичника.
КОРТИКАЛЬНЫЕ ГРАНУЛЫ
В ооцитах многих животных (плоские, круглые и кольчатые черви, моллюски, членистоногие, иглокожие, хордовые, в т.ч. ланцетник, миноги, осетровые и костистые рыбы, амфибии, рептилии, млекопитающие) в ходе оогенеза происходит накопление особых кортикальных гранул, которые формируются при участии элементов АГ. Изначально они рассеяны по всей цитоплазме, но на завершающих этапах оогенеза концентрируются в кортикальной области ооцита. Обычно это сравнительно мелкие пузырьки, диаметром до 1 мкм, но у осетровых рыб они достигают 3-4 мкм, а у костистых они называются кортикальными альвеолами и достигают 30-40 мкм. Кортикальные гранулы содержат разного рода ферменты, сульфомукополисахариды и другие вещества, которые играют важную роль в процессе оплодотворения.
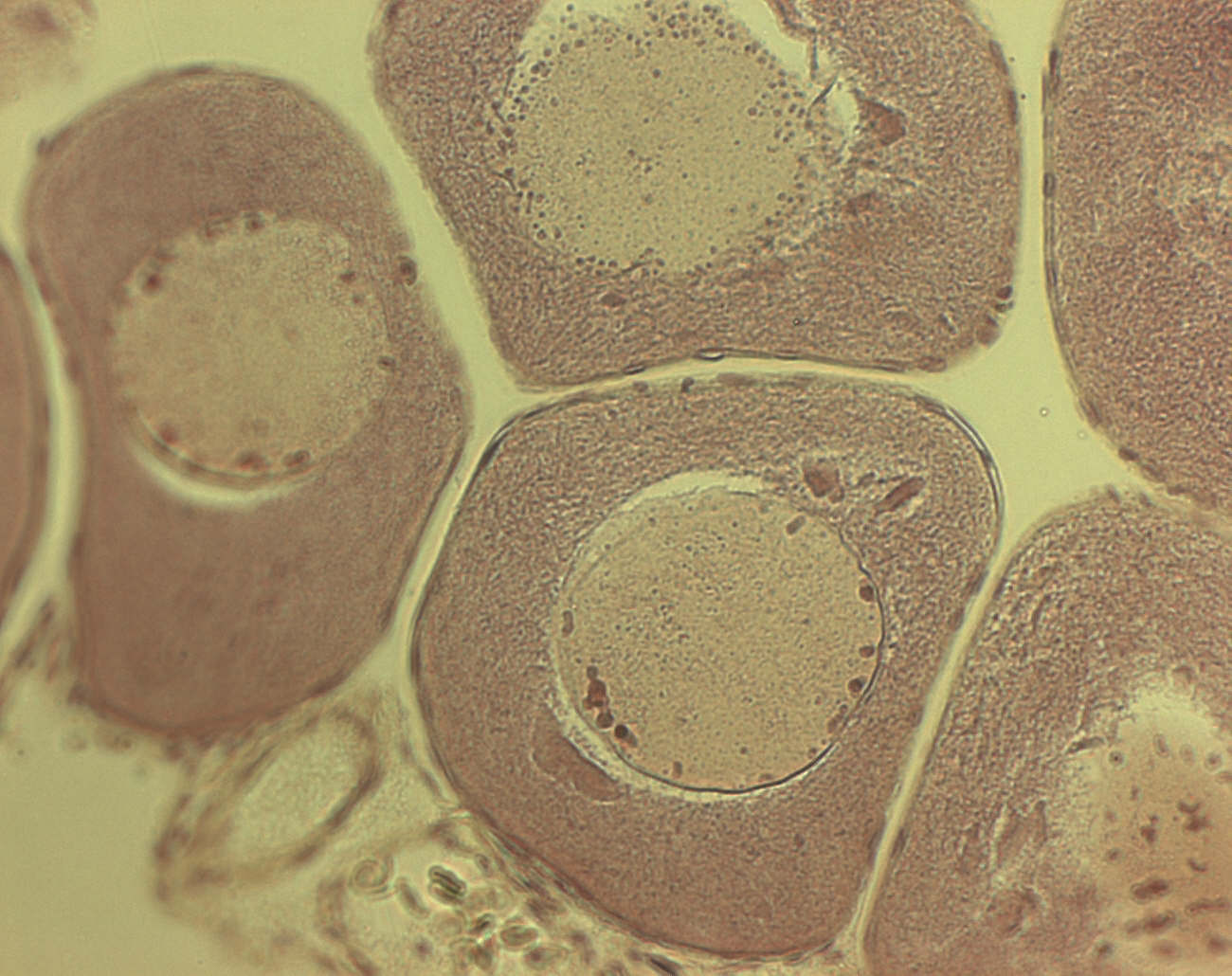
1
2
Рис.1 Ооцит малого роста. 1- фоликулярный эпителий, 2-ядро
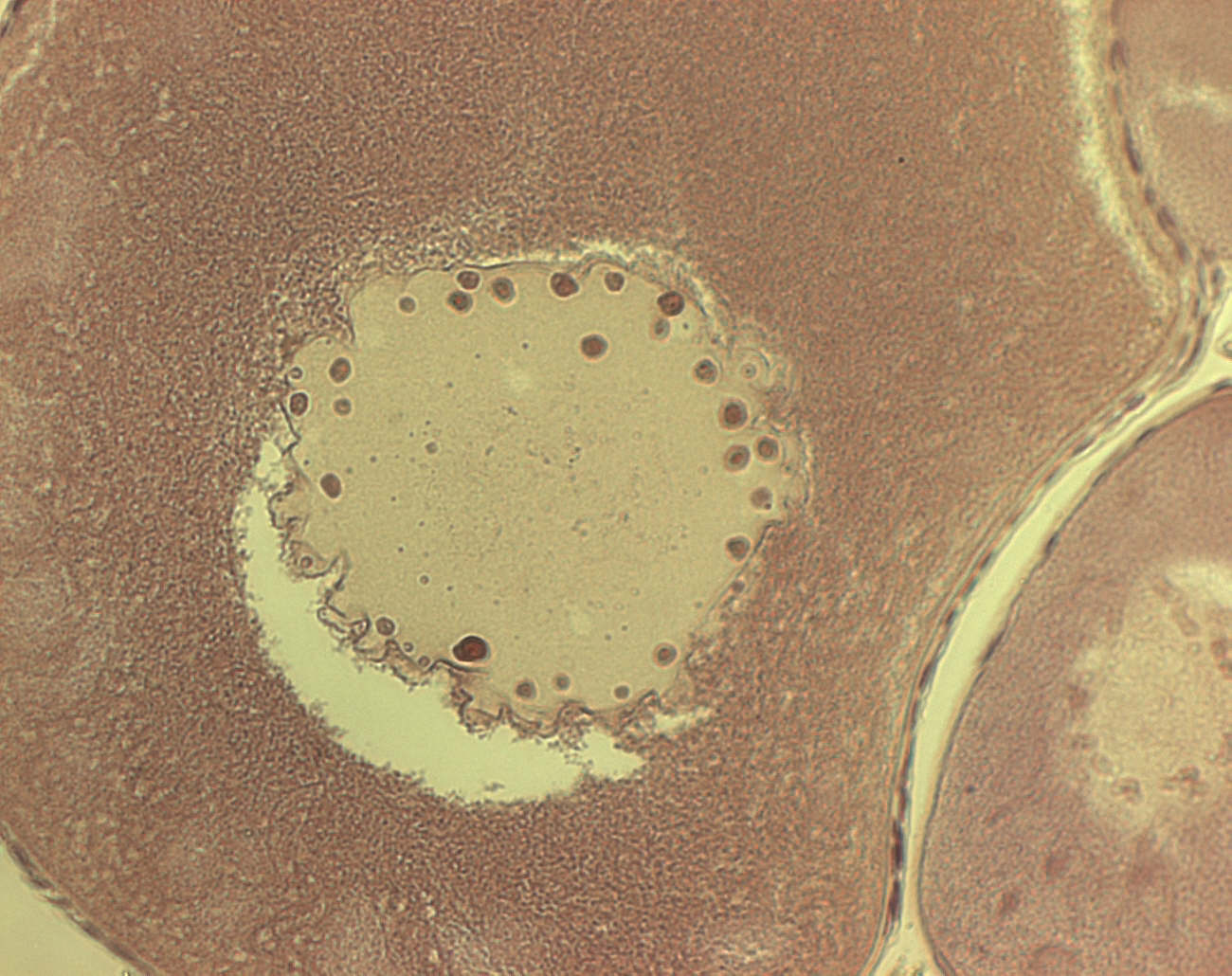
Рис.2 Лопастное ядро ооцита на стадии большого роста
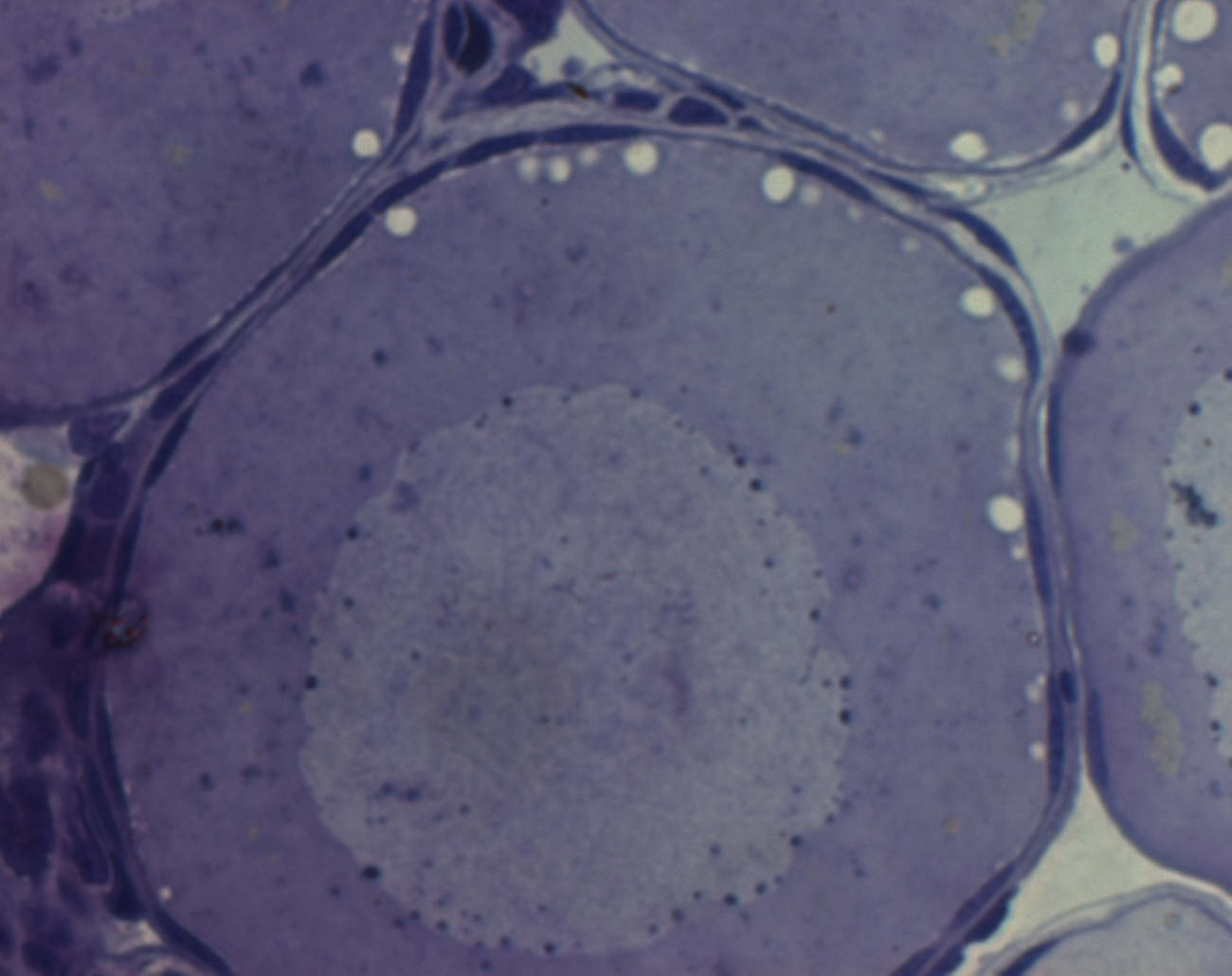
Рис.3 Вакуолизация
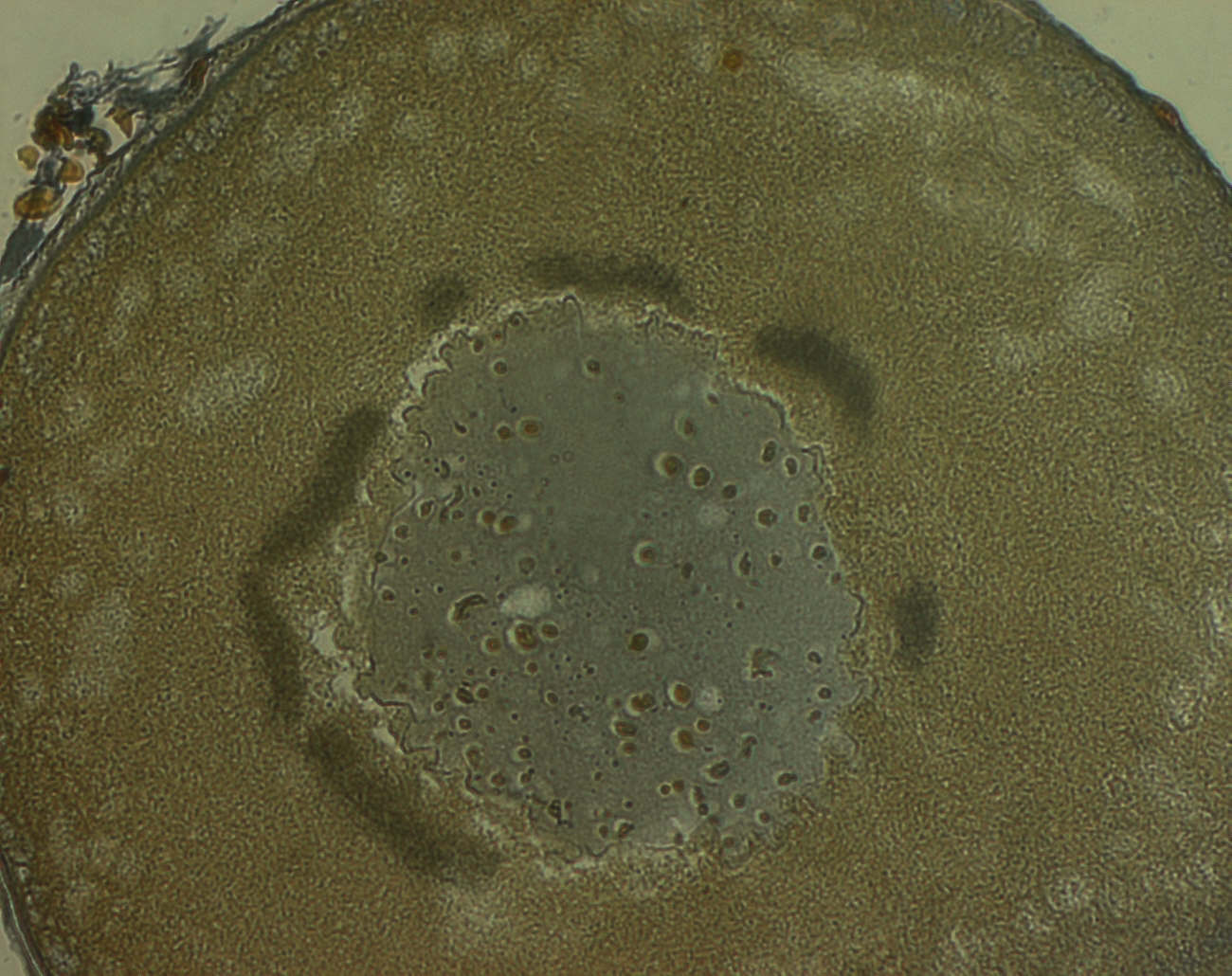
Рис.4 Тельце Бальбиани