

МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
имени М.В. ЛОМОНОСОВА
Биологический факультет
Методическая разработка по спецкурсу
ГАМЕТОГЕНЕЗ
Москва, 2012 г.
Филатов М. А.
Занятие №1. Первичные половые клетки.
В рамках данного курса студентам предстоит изучить особый раздел эмбриологии – гаметогенез. Хотя непосредственным предметом науки «эмбриология» в узком смысле этого слова является зародыш на разных этапах эмбрионального развития, очевидно, что изучение нового организма, начинающего свое существование после процесса оплодотворения, т.е. слияния отцовской и материнской половых клеток, не будет полноценным, если не иметь представления о самих половых клетках.
Гаметогенез – это сложный процесс, в результате которого происходит образование половых клеток - гамет. Гамета – это репродуктивная клетка (то есть клетка, принимающая участие в процессе размножения) животных и растений. Как правило, гаметы обладают гаплоидным набором хромосом. В процессе гаметогенеза происходит дифференцировка высокоспециализированных половых клеток, связанная как со специфическими изменениями ядерного аппарата (прежде всего – редукция числа хромосом), так и с существенным преобразованием клетки в целом (например, накопление питательных веществ яйцеклеткой и уменьшение объема цитоплазмы у сперматозоида в сравнении с другими клетками).
Гаметогенез у животных бывает двух типов: локализованный и диффузный.
При диффузном гаметогенезе гаметы развиваются в любом участке тела. Диффузный гаметогенез наблюдается у губок, некоторых кишечнополостных, плоских червей.
В случае локализованного гаметогенеза гаметы развиваются в половых железах – гонадах. Локализованный гаметогенез характерен для подавляющего большинства животных. В гонадах помимо развития половых клеток также осуществляется синтез различных гормонов.
Существуют различные формы полового размножения, однако в данном курсе рассматривается гаметогенез животных, при этом мы будем иметь дело с частным случаем гетерогамии – оогамией. Это - такой тип полового процесса, при котором в ходе оплодотворения гаметы, резко различные по размерам, форме и поведению, сливаются, образуя зиготу. Оогамия свойственна всем многоклеточным животным, многим низшим и высшим растениям.
Прежде чем рассматривать подробно два частных случая гаметогенеза: образование яйцеклеток – оогенез, и образование сперматозоидов – сперматогенез, необходимо остановиться на вопросе о происхождении половых клеток.
Первичные половые клетки.
Д искуссия
о происхождении половых клеток ведет
свое начало со второй половины 19 века.
Тогда оформилось две основные теории
относительно происхождения половых
клеток.
искуссия
о происхождении половых клеток ведет
свое начало со второй половины 19 века.
Тогда оформилось две основные теории
относительно происхождения половых
клеток.
Вильгельм Вальдейер (Waldeyer, 1870) считал, что развитие половых клеток у высших животных возможно из эпителия гонады, т.е. из дифференцированных клеток половой железы. Поэтому он называл эпителий гонады герминативным эпителием. Впоследствии была доказана ошибочность его взглядов, однако термин остался. Итак, наряду с Мюллером и Гертвигом, Вальдейер полагал, что существует качественное единство всех клеток организма. В пользу этой теории говорило и то, что позднее было показано генетическое тождество всех клеток организма.
Август Вейсман (1834-1914)

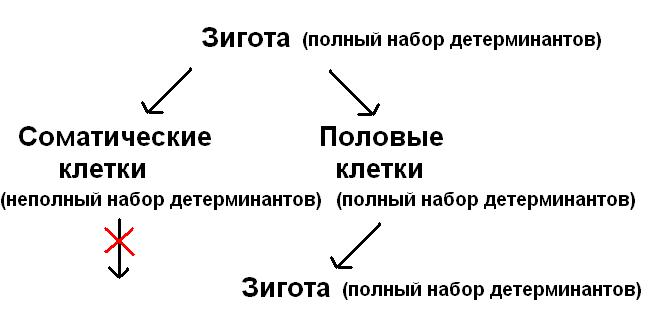
Нусбаум (Nussbaum, 1880) и Вейсман (Weismann, 1885, 1892) придерживались другой точки зрения. Впервые Нусбаум подчеркнул факт раннего обособления половых клеток в развитии и высказал идею о непрерывности линии этих клеток в ряду поколений – идею «зародышевого пути». Затем в работах Вейсмана гипотеза зародышевого пути получила дальнейшее развитие и была сформулирована «теория зародышевой плазмы». Теория основана на представлении о том, что при дифференцировке клеток зародыша из ядра в цитоплазму выходят некие частицы (детерминанты) – носители зачатков различных признаков, определяющие организацию всех клеток тела. Совокупность детерминантов он назвал зародышевой плазмой (идиоплазмой). Оплодотворенное яйцо по Вейсману содержит всю зародышевую плазму, а образующиеся при делении потомки зиготы теряют те или иные детерминанты. Полный набор детерминантов остается только в клетках половой линии, что и является их главным отличием от линии соматических клеток. Взгляды Нусбаума и Вейсмана слились в единую теорию Нусбаума-Вейсмана. Однако чаще сейчас встречается именно теория Вейсмана, говорящая о непрерывности зародышевой плазмы. Согласно этой теории существует две линии клеток – соматические и половые. В онтогенезе эти две линии расходятся очень рано, и затем переход между ними невозможен. Соматические клетки обеспечивают существование организма, и представляют собой тупиковый, «смертный» путь развития. Половые клетки по Вейсману являются как бы «бессмертной» линией, так как участвуют в размножении, передавая потомкам зародышевую плазму и обеспечивая непрерывность поколений. Основываясь на представлениях Вейсмана, можно сказать, что прямые митотические предки наших собственных половых клеток когда-то находились в гонадах наших эволюционных предков (непрерывность!).
Теорию Вейсмана можно представить на наглядной схеме:
Вскоре после опубликования работ
Вейсмана его теория, казалось бы, получила
экспериментальное подтверждение в
исследованиях Бовери (Boveri,
1899). Он описал явление диминуции хроматина
у аскариды Ascaris megalocephala.
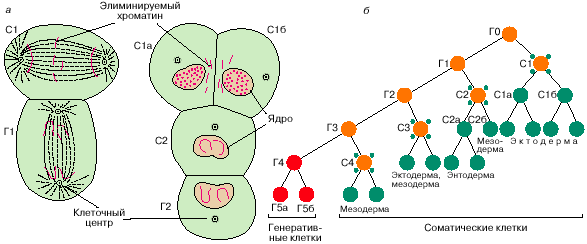
А - Цитологическая картина диминуции хроматина на примере лошадиной аскариды Parascaris univalens
Б - схема дифференцировки клеток зародышевого пути и соматических клеток на ранних стадиях эмбрионального развития аскариды. Пресоматические клетки С1, С2, С3 и С4, подвергшиеся диминуции, изображены кружками, окруженными четырьмя точками.
Уже на стадии 2 бластомеров хромосомы в одном из бластомеров фрагментируются, их дистальные участки отбрасываются и не входят в состав формирующегося ядра, затем это повторяется еще в трех клеточных делениях. В итоге полноценный набор хромосом остается только у одной клетки – родоначальницы половой линии. Все остальные клетки с неполноценными хромосомами дают начало только соматическим клеткам. Казалось бы, гипотеза подтвердилась, и найдена причина различия половых и соматических клеток. Вслед за этим Ру предложил и Вейсман разработал мозаичную теорию развития, согласно которой яйцо представляет собой мозаику предшествующих зачатков, а развитие – распределение материала этих зачатков по клеткам. Эта теория была основана на преформистских взглядах, и просуществовала недолго. Однако она оказалась полезной при развитии представлений об ооплазматической сегрегации и эквивалентности ядер в развитии.

Теодор Бовери (1826-1915)
Именно благодаря этой теории сегодня мы знаем о существовании животных с крайним мозаичным типом развития, присущим первичноротым, например нематоде C. elegans, у которой подробно описана судьба каждого бластомера, и при удалении того или иного бластомера, можно получить организм с отсутствующими клетками определенного типа. Однако сегодня мы также знаем и о существовании регуляций в развитии – способности бластомеров принимать судьбу утраченных, заменяя их и становясь на новый для себя путь развития. Регуляционное развитие присуще в основном вторичноротым.
Теория Вейсмана несомненно очень важна для понимания проблемы происхождения половых клеток. Однако, его взгляды претерпели со временем изменения:
Во-первых, диминуция хроматина была найдена еще у некоторых видов аскарид, плавунца, а среди позвоночных – у некоторых миксин. У остальных животных диминуции хроматина не наблюдается. Поэтому ее нельзя считать причиной разделения половой и соматической линий клеток.
Во-вторых, не у всех животных половые и соматические клетки навсегда расходятся в начале онтогенеза. Так, у кишечнополостных, губок, плоских червей и оболочников соматические клетки легко становятся половыми даже у взрослого организма (например, трансформация высокоспециализированных хоаноцитов губок в половые клетки). Сам Вейсман говорил о существовании клеток-предшественников обеих клеточных линий – как половой, так и соматической (фактически, подошел к проблеме стволовых клеток). Нусбаум же утверждал, что обособление половых клеток происходит исключительно в процессе первых делений дробление, и яйцо является непосредственным предшественником ППК. Он подчеркивал существование недифференцированных клеток у кишечнополостных (интерстициальные клетки), из которых образуются как половые, так и соматические клетки на протяжении всей жизни организма.
В-третьих, появилась теория ооплазматической сегрегации, и представления о том, что именно зародышевая плазма (специфический участок цитоплазмы яйца, содержащий некие факторы), наследуемая в результате дробления определенными бластомерами, действует на ядра будущих первичных половых клеток, определяя их развитие по этому пути. Согласно современным представлениям, зародышевая плазма – область цитоплазмы с определенной морфологической структурой, в состав которой входят цистерны ЭПР, митохондрии, электронноплотные гранулы сложного строения, белки, РНК материнского происхождения.
Различие соматических клеток между собой и их отличие от половых клеток можно теперь трактовать не разницей в наличии половых детерминантов, а дифференциальной активностью генов и различными паттернами метилирования ДНК в этих клетках.
В настоящее время происхождение половых клеток изучено у многих животных. Зачаток половых клеток получил название примордиальные, или первичные половые клетки (ППК, гоноциты) – primordial germ cells. Некоторые исследователи выделяют этот зачаток как отдельный – четвертый – зародышевый листок. Мы знаем, что ППК у большинства животных закладываются в онтогенезе достаточно рано, в дальнейшем переход между соматическими и половыми клетками, как правило, невозможен. ППК закладываются всегда независимо от зачатка гонады, то есть экстрагонадно, и лишь позже мигрируют, и в итоге заселяют зачаток гонады. Отличительные особенности ППК: размеры 10-22 мкм, очень большое пузыревидной ядро, наличие особых структур в цитоплазме.
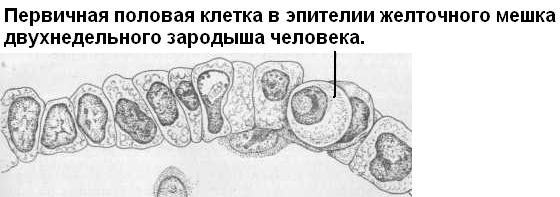
П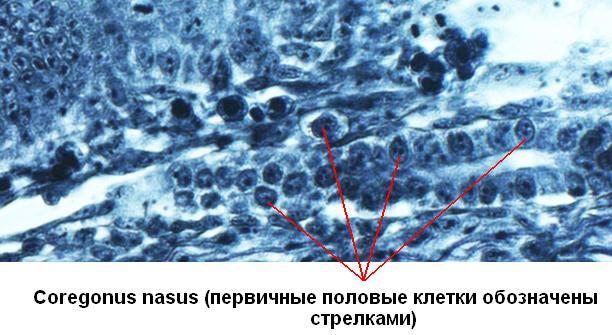 роцессы,
приводящие к экспрессии специфических
генов и образованию ППК, можно разделить
на:
роцессы,
приводящие к экспрессии специфических
генов и образованию ППК, можно разделить
на:
- ооплазматическую сегрегацию (транспорт молекул в те компартменты яйца, где произойдет закладка ППК);
- механизмы неравномерного наследования половой плазмы предшественниками ППК и соматических клеток;
- дифференциальную посттранскрипционную регуляцию генов в половой и соматической линиях;
- поддержание плюрипотентности и особого статуса ППК.
Рассмотрим частные примеры обособления ППК у различных животных. При этом в каждом отдельном случае укажем методы идентификации ППК, их локализацию, способы пролиферации и миграции.
В целом, сейчас принято выделять два типа детерминации линии половых клеток:
- обособление части клеток зародыша в результате наследования определенных детерминантов материнского происхождения, сконцентрированных в особой зоне цитоплазмы в следствие ооплазматической сегрегации. Такой тип характерен для беспозвоночных, а также для таких позвоночных, как бесхвостые амфибии. У этих животных зародышевая плазма морфологически выражена и содержит гранулы. У рыб и птиц половую плазму возможно маркировать;
- индукция ППК на более поздних стадиях развития. Это характерно для млекопитающих и хвостатых амфибий.
Рассмотрим теперь более детально формирование первичных половых клеток у различных групп животных.