

Политенные хромосомы
Впервые обнаружены Бальбиани в 1881-го, однако их цитогенетическая роль была выявлена Костовым, Пайнтером, Гейтцем и Бауером. Содержатся в клетках слюнных желёз,кишечника, трахей, жирового тела и мальпиг-х сосудов личинок двухкрылых.
Гигантские (политенные) хромосомы образуются в результате явления, называемого Политенией, когда число хромонем в хромосомах увеличивается, но образующиеся хроматиды не расходятся, расщепления хромосом не происходит, и они поэтому утолщаются, становясь крупными. Гигантские хромосомы используют для изучения функции генов в онтогенезе, а также для анализа мутационной изменчивости, происходящей в результате хромосомных перестроек под влиянием различных мутагенных факторов.
Явление совместного наследования генов, локализующихся в одной паре хромосом, было названо сцеплением генов. Морган наблюдал его у Дрозофилы при возвратном скрещивании гибридной особи, полученной от скрещивания серой мухи с нормальными крыльями с черной мухой, имеющей зачаточные крылья. В потомстве анализирующего скрещивания гибридного самца с самкой, несущей в гомозиготном состоянии рецессивные гены черного цвета тела и зачаточных крыльев, были обнаружены только два класса особей, повторяющих по фенотипу исходные родительские. Это служит генетическим доказательством сцепления генов. Сцепленное наследование – совместное наследование генов, ограничивающее их свободное комбинирование. Группа сцепления – совокупность генов, находящихся на одной хромосоме. Однако абсолютно полного сцепления генов почти не бывает. Оно обнаружено в основном только у самцов дрозофилы. Частота сцепления генов может нарушаться, вследствие чего гены, лежащие в разных участках гомологичных хромосом, могут разъединяться с сочетаться друг с другом в новых комбинациях. Причиной нарушения сцепления генов является кроссинговер.
№ 21
К нерегулярным типам полового размножения можно отнести партеногенетическое, гиногенетическое и андрогенетическое размножение животных и растений (рис. 27).
Партеногенез — это развитие зародыша из неоплодотворенной яйцеклетки. Явление естественного партеногенеза свойственно низшим ракообразным, коловраткам, перепончатокрылым (пчелам, осам) и др. Известен он также у птиц (индейки). Партеногенез можно стимулировать искусственно, вызывая активацию неоплодотворенных яиц путем воздействия различными агентами.
Различают партеногенез соматический, или диплоидный, и генеративный, или гаплоидный. При соматическом партеногенезе яйцеклетка не претерпевает редукционного деления или если и претерпевает, то два гаплоидных ядра, сливаясь вместе, восстанавливают диплоидный набор хромосом (автокариогамия) ; таким образом в клетках тканей зародыша сохраняется диплоидный набор хромосом.
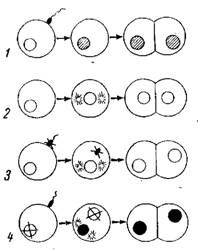 При
генеративном партеногенезе зародыш
развивается из гаплоидной яйцеклетки.
Например, у медоносной пчелы (Apis mellifera)
трутни развиваются из неоплодотворенных
гаплоидных яиц путем партеногенеза.
При
генеративном партеногенезе зародыш
развивается из гаплоидной яйцеклетки.
Например, у медоносной пчелы (Apis mellifera)
трутни развиваются из неоплодотворенных
гаплоидных яиц путем партеногенеза.
Различные типы полового размножения:
1 — нормальное оплодотворение; 2 — партеногенез: 3 — гиногенез; 4 — андрогеиез.
Партеногенез у растений очень часто называют апомиксисом. Поскольку апомиксис широко распространен в растительном мире и имеет большое значение при изучении наследования, рассмотрим его особенности.
Наиболее распространенным типом апомиктического размножения является тип пар- теногенетического образования зародыша из яйцеклетки. При этом чаще встречается диплоидный апомиксис (без мейоза).
Наследственная информация и при образовании эндосперма, и при образовании зародыша получается только от матери. У некоторых апомиктов для формирования полноценных семян необходима псевдогамия —- активация зародышевого мешка пыльцевой трубкой. При этом один спермий из трубки, достигая зародышевого мешка, разрушается, а другой сливается с центральным ядром и участвует только в образовании ткани эндосперма (виды из родов Potentilla, Rubus и др.). Наследование здесь происходит несколько отлично от предыдущего случая. Зародыш наследует признаки только по материнской линии, а эндосперм — и материнские и отцовские.
Гиногенез. Очень сходно с партеногенезом гиногенетическое размножение. В отличие от партеногенеза при гиногенезе участвуют сперматозоиды как стимуляторы развития яйцеклетки (псевдогамия), но оплодотворения (кариогамии) в этом случае не происходит; развитие зародыша осуществляется исключительно за счет женского ядра (рис. 27, 3). Гиногенез обнаружен у круглых червей, живородящей рыбки Molliensia formosa, у серебряного карася (Platypoecilus) и у некоторых растений—■ лютика (Ranunculus auricomus), мятлика (род Роа pratensis) и др.
Гиногенетическое развитие можно вызвать искусственно, если перед оплодотворением сперму или пыльцу облучить рентгеновыми лучами, обработать химическими веществами или подвергнуть действию высокой температуры. При этом разрушается ядро мужской гаметы и теряется способность к кариогамии, но сохраняется способность к активации яйца.
Явление гиногенетического размножения имеет большое значение для изучения наследственности, так как при этом потомство получает наследственную информацию только от матери. Таким образом, при бесполом размножении, партеногенезе и гиногенезе потомство должно быть сходно только с материнским организмом.
Андрогенез. Прямой противоположностью гиногенеза является андрогенез. При андрогенезе развитие яйца осуществляется только за счет мужских ядер и материнской цитоплазмы (рис. 27, 4). Андрогенез может иметь место в тех случаях, когда материнское ядро почему-либо погибает до момента оплодотворения.
Если в яйцеклетку попадает один сперматозоид, то развивающийся зародыш с гаплоидным набором хромосом оказывается нежизнеспособным или маложизнеспособным. Жизнеспособность андрогенных зигот нормализуется, если восстанавливается диплоидный набор хромосом. Как было уже показано, в тех случаях, когда происходит полиспермия у животных и в яйцеклетку одновременно проникает несколько сперматозоидов, возможно слияние двух отцовских пронуклеусов и образование диплоидного ядра. Развитие андрогенных особей до взрослого состояния наблюдалось лишь у тутового шелкопряда (Bombyx mori) и паразитической осы (Habrobracon juglandis). Андрогенетическое размножение, как исключение, обнаружено и у некоторых растений (табак, кукуруза и др.).
№ 29
Моногибридным называется скрещивание, при котором родительские формы отличаются друг от друга по одной паре контрастных, альтернативных признаков.
Признак —любая особенность организма, т. е. любое отдельное его качество или свойство, по которому можно различить две особи. У растений это форма венчика (например, симметричный—асимметричный) или его окраска (пурпурный—белый), скорость созревания растений (скороспелость—позднеспелость), устойчивость или восприимчивость к заболеванию и т. д.
Примерами моногибридного скрещивания, проведенного Г. Менделем, могут служить скрещивания гороха с такими хорошо заметными альтернативными признаками, как пурпурные и белые цветки, желтая и зеленая окраска незрелых плодов (бобов), гладкая и морщинистая поверхность семян, желтая и зеленая их окраска и др.
Единообразие гибридов первого поколения (первый закон Менделя) – у гибридов первого покаления появл только 1 из пары альтернат признаков, доминантный. При скрещивании гороха с пурпурными и белыми цветками Мендель обнаружил, что у всех гибридных растений первого поколения (F1) цветки оказались пурпурными. При этом белая окраска цветка не проявлялась.
Мендель установил также, что все гибриды F1 оказались единообразными (однородными) по каждому из семи исследуемых им признаков.
Следовательно, у гибридов первого поколения из пары родительских альтернативных признаков проявляется только один, а признак другого родителя как бы исчезает. Явление преобладания у гибридов F1 признаков одного из родителей Мендель назвал доминированием, а соотв-й признак — доминантным. Признаки, не проявляющиеся у гибридов F1 он назвал рецессивными.
Поскольку все гибриды первого поколения единообразны, это явление было названо К. Корренсом первым законам Менделя, или законом единообразия гибридов первого поколения, а также правилом доминирования.
№10
Второй закон Менделя (закон расщепления): Во втором поколении появляются особи как с доминантными признаками, так и с пецессивными в соотношении 3:1 (расщепление по фенотипу). 1:2:1 – расщепление по генотипу.
Генотип – совокупность наследственных задатков.
Фенотип – совокупность свойств и признаков организма, которые являются результатом взаимодействия генов особи и окружающей среды.
Гены обозначаются буквами латинского алфавита.
Аллель – одно из возможных состояний гена.
Константные формы: АА и аа, которые имеют одинаковые аллели – гомозиготы, разные Аа, Вв – гетерозигота.
Несмешивание аллелей каждой пары альтернативных признаков в гаметах гибридного организма – явление чистоты гамет.
Гомозиготный организм образует 1 тип гамет АА – А, аа – а
Гетерозигота – 2 типа гамет Аа - А а
Совокупность всех признаков организма, начиная с внешних и кончая особенностями строения и функционирования клеток, тканей и органов, называется фенотипом. Этот термин может употребляться и по отношению к одному из альтернативных признаков.
Признаки и свойства организма проявляются под контролем наследственных факторов, т. е. генов. Совокупность всех генов организма называют генотипом.
№ 11
Анализирующее скрещивание – скрещивание гибрида с исходной формой, гомозигосной по рецессивному гену.
№12
Доминирование и др. взаимодействия генов. Различают полное доминирование, неполное доминирование, кодоминирование, аллельное исключение.
Полное доминирование — это вид взаимодействия аллельных генов, при котором фенотип гетерозигот не отличается от фенотипа гомозигот по доминанте, то есть в фенотипе гетерозигот присутствует продукт доминантного гена. Полное доминирование широко распространено в природе, имеет место при наследовании, например, окраски и формы семян гороха, цвета глаз и цвета волос у человека, резус-антигена и мн. др.
Неполное доминирование. Так называется вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву и имеет среднее (промежуточное) значение между ними. Имеет место при наследовании окраски околоцветника ночной красавицы, львиного зева, окраски шерсти морских свинок и пр.
Кодоминирование — вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву, и в фенотипе гетерозигот присутствуют продукты обоих генов. Имеет место при формировании, например, IV группы крови системы (АВ0) у человека.
Аллельным исключением называется отсутствие или инактивация одного из пары генов; в этом случае в фенотипе присутствует продукт другого гена (гемизиготность, делеция, гетерохроматизация участка хромосомы, в котором находится нужный ген).
В некоторых случаях имеются различия в передаче наследственных свойств со стороны материнского или отцовского организма, поэтому в генетике применяется реципрокное скрещивание (скрещивание двух форм в двух различных направлениях).
Желт. Х Зел.
Зел. Х Желт.
Для генетического анализа применяется скрещивание гибридов с одной из родительских форм – возвратное скрещивание (несёт данную пару аллелей в гомозиготном состоянии).
№ 13
Наследование — это процесс передачи генетической информации в ряду поколений. Наследуемые признаки могут быть качественными (моногенными) и количественными (полигенными).
III закон Менделя(Независимое наследование): отдельные парыпризнаков при дигибр скрещивании ведут себя в наследовании независимо, свободно сочетаясь др. с др. во всех возможных комбинациях.
Для дигибридного скрещивания Мендель использовал гомозиготные растения гороха, различающиеся одновременно по двум парам признаков. Одно из скрещиваемых растений имело желтые гладкие семена, другое-зеленые морщинистые. Все гибриды первого поколения этого скрещивания имели желтые гладкие семена. Следовательно, доминирующими оказались желтая окраска семян над зеленой и гладкая форма над морщинистой. Обозначим аллели желтой окраски А, зеленой-а, гладкой формы-В, морщинистой-b. Гены, определяющие развитие разных пар признаков, называются неаллельпыми и обозначаются разными буквами латинского алфавита. Родительские растения в этом случае имеют генотипы ААВВ и aabb, а генотип гибридов F1 -АаВb ,т. е. является дигетерозиготным.
Во втором поколении после самоопыления гибридов F1 в соответствии с законом расщепления вновь появились морщинистые и зеленые семена. При этом наблюдались след. сочетания признаков: 315 желтых гладких, 101 желтое морщ-е, 108 зеленых гладких и 32 зеленых морщин-х семян. Это соотношение близко к соотн-ю 9:3:3:1.
Комбинативная изменчавость – появление новых комбинаций признаков врезульт скрещивания.
Сущность дигибридного скрещивания. Организмы различаются по многим генам и, как следствие, по многим признакам. Чтобы одновременно проанализировать наследование нескольких признаков, необходимо изучить наследование каждой пары признаков в отдельности, не обращая внимания на другие пары, а затем сопоставить и объединить все наблюдения. Именно так и поступил Мендель.
Скрещивание, при котором родительские формы отличаются по двум парам альтерн-х признаков (по двум парам аллелей), называется дигибридным. Гибриды, гетерозиготные по двум генам, называют дигетерозиготными, а в случае отличия их по трем и многим генам —три- и полигетерозиготными соответственно.
Результаты ди- и полигибридного скрещивания зависят от того, располагаются гены, определяющие рассмотренные признаки, в одной хромосоме или в разных.
Полигибридное скрещивание – скрещивание форм, отличающихся друг от друга, по нескольким парам альтернативных признаков. При этом особь, гетерозиготная по n парам генов, может произвести 2n типов гамет, а в F2, при расщеплении потомства полигибридного скрещивания может образоваться 3n генотипов. Частоту данного генотипа в потомстве родителей, отличающихся определенным числом независимо наследуемых генов, можно вычислить следующим образом: надо подсчитать вероятность соответствующего генотипа для каждой пары генов отдельно, а затем перемножить. Например, надо рассчитать частоту генотипа AabbCc в потомстве от скрещивания АаВЬсс ×АаВЬСс. Вероятность генотипа Аа в потомстве от скрещивания Аа × Аа равна 1/2; вероятность генотипа bb в потомстве от скрещивания ВЬ × ВЬ равна 1/4; вероятность генотипа Сс равна также 1/2. Следовательно, вероятность генотипа AabbCc составляет 1/2*1/4*1/2 = 1/16.
№ 16
Эпистаз – явление, при котором ген из одной аллельной пары подавляет ген из другой аллельной пары
Гены подавители: супрессоры и ингибиторы. Могут быть доминирующими и рецессивными. Встречаются у растений и животных.
Эпистаз:
Доминантный эпистаз – подавление доминантной аллелью одного гена действия аллельной пары другого гена.
Наследование окраски оперения у кур:
С – окрашенное оперение
с – белое оперение
I > C, i – не влияет на окраску
P: ♀ CCII Х ♂ ccii
бел бел
F1: CcIi
бел
F2: 9 C-I- : 3 –C-ii : 3 cci- : 1 ccii
Бел окр бел бел
13:3 – доминантный эпистаз
При доминантном эпистазе встречается расщепление 12:3:1. Наблюдается при наследовании окраски кожуры у тыквы и лука.
Рецессивный эпистаз – рецессивная аллель одного гена, будучи в гомозиготном состоянии не даёт возможности проявиться доминантным или рецессивным аллелям других генов.
Под рецессивным эпистазом понимают явления при котором рецессивная аллель каждого из генов находится в гомозиготном состоянии, подавляея действие доминантных аллелей других генов. В этом случае наблюдается расщепление 9:7
Гены-ингибиторы. Взаимодействие генов, при котором один ген подавляет действие другого называется эпистазом , а гены, подавляющие действие других генов, называются генами-супрессорами или генами-ингибиторами. В некоторых случаях наличие одного доминантного гена подавляет проявление другого неаллельного гена (и доминантного, и рецессивного аллеля). Например, ген A обуславливает белый цвет плодов тыквы, при его наличии действие гена В не проявляется. При генотипе ааВВ или ааВb плоды тыквы имеют желтый цвет. Наконец, если оба гена представлены рецессивными аллелями, плоды тыквы имеют зеленый цвет. По этому же типу может наследоваться окраска шерсти собак , овец и других животных.
Под комплементарностью понимают такой тип взаимодействия генов, при котором два гена вместе обусловливают развитие нового признака, отличного от родительских вариантов. Существует не менее трех типов комплементарности:
1) доминантные гены различаются по фенотипическому проявлению;2) доминантные гены имеют сходное фенотипическое проявление; 3) и доминантные, и рецессивные гены имеют самостоятельное фенотипическое проявление.
Если доминантные аллели двух генов обусловливают разный фенотип, то в F, наблюдается расшепление 9:3:3:1. В качестве примера данного типа взаимодействия генов можно привести наследование формы гребня у кур.
У гибридов первого поколения доминантные гены А и В дополняют друг друга и вместе обусловливают ореховидную форму гребня, которой не было у родительских форм. При скрещивании гибридов F1: AaBb x AaBb во втором поколении, наряду с ореховидной, розовидной и гороховидной появляется простая форма гребня в соотношении: 9 А_ B_ : 3 А_ bb : 3 аа В : 1 аа bb («_» означает, что аллель в гомологичной хромосоме может быть как доминантным, так и рецессивным). В отличие от менделевского расщепления, наблюдаемого во втором поколении дигибридного скрещивания, в данном случае в первом поколении два гена действуют на один признак.
При другом варианте комплементарности доминантные аллели двух взаимодействующих генов не имеют собственного фенотипического проявления: новый фенотип у гибридов определяется одновременным присутствием в генотипе двух неаллельных доминантных генов.
Явление взаимодействия генов, при котором для проявления признака необходимо наличие двух неаллельных генов, называют комплементарностью (от лат. complementum – дополнение), а гены, необходимые для проявления признака, комплементарными, или дополнительными.
К омплементарное
взаимодействие генов. В том случае,
когда признак образуется при наличии
двух доминантных неаллельных генов,
каждый из которых не имеет самостоятельного
фенотипи-ческого выражения, гены
обозначают как комплементарные.
омплементарное
взаимодействие генов. В том случае,
когда признак образуется при наличии
двух доминантных неаллельных генов,
каждый из которых не имеет самостоятельного
фенотипи-ческого выражения, гены
обозначают как комплементарные.
№ 8
Генетическая инжене́рия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов изорганизма (клеток), осуществления манипуляций с генами и введения их в другие организмы.
Чтобы встроить ген в вектор, используют ферменты — рестриктазы и лигазы, также являющиеся полезным инструментом генной инженерии. С помощью рестриктаз ген и вектор можно разрезать на кусочки. С помощью лигаз такие кусочки можно «склеивать», соединять в иной комбинации, конструируя новый ген или заключая его в вектор. За открытие рестриктаз Вернер Арбер, Даниел Натанс и Хамилтон Смит также были удостоены Нобелевской премии (1978 г.).
Успехи генетической инженерии стали возможны благодаря изучению ферментов, позволяющих проводить химические операции с генетическим материалом. В генетической инженерии используется большая группа ферментов, способных гидролизовать, «разрезать» молекулы ДНК и РНК по специфическим центрам, «сшивать» фрагменты нуклеиновых кислот с образованием фосфодиэфирных связей, проводить полимеризацию нуклеотидов на матрице одноцепочечной ДНК или РНК. Совокупность этих свойств ферментов позволяет манипулировать ДНК, проводить ее модификацию.
Рестриктазы – это ферменты разрезающие ДНК по определенным нуклеотидным последовательностям. Главный инструмент генной инженерии. Рестриктазы – ферменты внутриклеточного иммунитета. После удаления Рестриктазы ДНК восстанавливаются.
Все ферменты делят на:
используемые для получения фрагментов ДНК
синтезирующие фрагменты ДНК на матрице ДНК
соединяющие фрагменты ДНК
позволяющие осуществлять изменение структуры концов фрагментов ДНК
применяемые для приготовления гибридизированных проб.
Каждый фермент, способный разрушить чужеродное ДНК, опознает в ней специфическую последовательность из 4-6 нуклеотидов. Соответств. последовательности в геноме бактерий замаскированы метированием остатков с помощью метилаз. ДНК - лигазы – ведут сшивку олигонуклеомидов между собой с образованием 2-х цепочных нуклеомидов. Ревертазы - (обратная транскриптаза) (РНК -зависимая ДНК-полимераза)- фермент РНК-содержащих вирусов, осуществляющих обратную транскрипцию, т.е. синтез ДНК провирусов на матрице вирусной РНК.
Векторы-переносчики –(плазмиды, вирусы)-это транскриптн. механизмы для ген. инф-ии.
Рестриктазы (рестрицирующие эндонуклеазы, эндонуклеазы рестрикции) - это ферменты, узнающие и атакующие определенные последовательности нуклеотидов в молекуле ДНК (сайты рестрикции).
Еще в 1953 году было обнаружено, что ДНК определенного штамма E. coli, введенная в клетки другого штамма (например, ДНК штамма В - в клетки штамма С) не проявляет, как правило, генетической активности, так как быстро расщепляется на мелкие фрагменты. В 1966 году было показано, что это явление связано со специфической модификацией хозяйской ДНК - она содержит несколько метилированных оснований, отсутствующих в немодифицированной ДНК, причем метилирование (добавление к основанию метильной группы) происходит уже после завершения репликации. Бактерия способна отличить свою собственную ДНК от любой вторгающейся «чужеродной» именно по типу ее модификации. За «метку» отвечают метилирующие ферменты модификации, так называемые ДНК-метилазы. Различие в модификации делает чужеродную ДНК чувствительной к действию рестрицирующих ферментов, которые узнают отсутствие метильных групп в соответствующих сайтах.
Системы рестрикции и модификации широко распространены у бактерий; их существование играет важную роль в защите резидентной ДНК от загрязнения последовательностями чужеродного происхождения.
Рестриктаза, которая расщепляла неметилированную ДНК была выделена в 1968 г. Мезельсоном и Юанем. Этот фермент был высокоспецифичен по отношению к определенной последовательности ДНК, но расщеплял молекулы неспецифически, в другом месте, на некотором удалении от участка узнавания. Вскоре, в 1970 г. Смит и Вилькокс выделили из Haemophilus influenzae первую рестриктазу, которая расщепляла строго определенную последовательность ДНК (Hind III). Поскольку разные бактерии по-разному метят свою ДНК, то и рестриктазы должны узнавать разные последовательности. И действительно, с тех пор выделены рестриктазы, узнающие более 150 сайтов рестрикции (мест расщепления ДНК).
Бактериальные плазмиды. Основная масса клеточной ДНК бактерий содержится в хромосоме. Однако кроме хромосом бактерии содержат большое количество очень маленьких кольцевых молекул ДНК плазмид длиной несколько тысяч пар оснований. Такие мини-хромосомы называют плазмидами.
Как правило, плазмиды имеют в своем составе гены устойчивости к антибиотикам, ионам тяжелых металлов (R-плазмиды), а также гены, контролирующие катаболизм некоторых органических соединений (плазмиды биодеградации, или D-плазмиды). Поскольку эти гены находятся в плазмидах, они представлены гораздо большим числом копий. Высокая копийность плазмид обеспечивает клетке синтез большого количества ферментов, химически нейтрализующих антибиотики или ксенобиотики, что и обеспечивает устойчивость к последним. Плазмиды, по-видимому, вездесущи, так как их выделяют из разных штаммов и видов бактерий, но не являются обязательными компонентами генома, а в некоторых природных штаммах плазмиды не обнаружены вообще.
Поскольку плазмидная ДНК значительно меньше хромосомной, ее довольно легко выделить в чистом виде. В присутствии ионов кальция плазмиды легко поглощаются бактериями-рецепиентами, Однако бактериальная клетка обычно может содержать в своем составе плазмиды одного типа. Это явление несовместимости плазмид. Существуют группы несовместимости – Inc-группы. В такой группе может быть несколько плазмид, совместимых между собой, но не совместимых с другими плазмидами. У этих плазмид сходны многие признаки и часто значительна гомология ДНК.
Число копий плазмиды в клетке может существенно варьировать. Это зависит от генетических особенностей как клетки, так и плазмиды. Плазмиды, находящиеся "под ослабленным контролем", могут размножаться до тех пор, пока их количество не достигнет 10-200 копий на клетку. Если же плазмида находится "под строгим контролем", она реплицируется с той же скоростью, что и главная хромосома. Такие плазмиды содержатся в клетке в одной или в нескольких копиях. Естественно, что для клонирования рекомбинантных ДНК стараются использовать плазмиды первого типа. Но это не обязательно, так как плазмиды в присутствии хлорамфеникола могут умножаться независимо от деления хромосомы, и количество копий плазмиды может многократно увеличиваться.
Одна их наиболее часто употребляемых плазмид для клонирования pBR 322 создана на основе плазмид природного происхождения, выделенных из E. coli. Эта плазмида содержит гены устойчивости к двум антибиотикам: ампициллину и тетрациклину, причем в генах устойчивости к этим антибиотикам имеются сайты рестрикции. Если фрагмент чужеродной ДНК встраивается в один из генов устойчивости, то последний инактивируется. Следовательно, успешное встраивание фрагмента чужеродной ДНК в один из этих генов легко детектировать по исчезновению у бактерий устойчивости к данному антибиотику. Но при этом сохраняется устойчивость к другому антибиотику. Таким образом вектор дает возможность детектировать только те клоны бактерий, которые содержат рекомбинантную плазмиду.
Вирусы
Есть вирусы, которые не ведут к гибели клетки, но встраиваются в геном клетки-хозяина и размножаются вместе с ней, либо вызывают ее неконтролируемый рост, т.е. превращают в раковую. К таким относятся ДНК-вирусы SV-40 и вирус полиомы. Внедрение некоторых опухолевых РНК-вирусов ведет к отпочковыванию вирусных частиц от клетки без ее лизиса. К таким вирусам относятся, например, ретровирусы (вирус саркомы Рауса и СПИДа). Для бактериальных клеток в качестве вектора часто используют бактериофаги.
Вирусы являются одними из главных кандидатов на роль векторов для введения чужеродной ДНК. При вирусной инфекции каждая клетка может получить большое число копий чужеродного гена. ДНК можно встраивать так, чтобы она находилась под контролем сильных вирусных промоторов, что обеспечит высокий уровень экспрессии гена, и его продукты будут более доступны для исследования.
сконструированы"челночные" векторы и их рекомбинантные производные, способные к репликации в животной и бактериальной клетке и эффективно экспрессирующие клонируемый ген в животной клетке. Наиболее распространенные векторы состоят из плазмиды рВR322.
Вирус должен быть жизнеспособным после рекомбинирования его ДНК. Легче всего вирусы вводятся в бактерии. Недостатком вирусов как векторов является их небольшая емкость. Кроме того, вирусы заражают небольшой круг хозяев.
№ 22
Изменчивость – свойство живых систем приобретать новые признаки, отличающие их от родительских форм.
Различают несколько типов изменчивости:
Наследственную (генотипическую) и ненаследственную (фенотипическую, паратипическую).
Индивидуальную (различие между отдельными особями) и групповую (между группами особей, например, различными популяциями данного вида). Групповая изменчивость является производной от индивидуальной.
Качественную и количественную.
Направленную и ненаправленную.
Модификационная (фенотипическая) изменчивость. В процессе индивидуального развития наблюдаются закономерные изменения морфологических, физиологических и др. особенностей организма. Такая изменчивость называется онтогенетической. Примером может являться постепенное физическое и умственное развитие человека. Все признаки и свойства организма наследственно детерминированы, однако организм наследует не сами признаки, а их возможность. Для того чтобы признак развился необходимы соответствующие условия внешней среды. Но даже в том случае, когда развитие признака происходит, степень его выраженности может варьировать в зависимости от условий внешней среды. Различия в проявлении одинаковых генотипов в различных условиях среды называется модификационной изменчивостью.
Но́рма реа́кции — способность генотипа формировать в онтогенезе, в зависимости от условий среды, разные фенотипы. Она характеризует долю участия среды в реализации признака. Чем шире норма реакции, тем больше влияние среды и тем меньше влияние генотипа в онтогенезе. Один и тот же ген в разных условиях среды может реализоваться в несколько проявлений признака (фенов).
Комбинативная (генотипическая) изменчивость. Изменение свойств и признаков организма может быть обусловлено изменением гена. Такие изменения называются мутациями. Изменчивость может быть обусловлена не только мутациями генов, но и различной их комбинацией. Комбинация генов может привести к появлению новых признаков или к новому их сочетанию. Такая изменчивость называется комбинативной и возникает в результате скрещивания.
Наследственная изменчивость — это форма изменчивости, вызванная изменениями генотипа, которые могут быть связаны с мутационной либо комбинативной изменчивостью.
Мутационная изменчивость
Гены время от времени подвергаются изменениям, которые получили название мутаций. Эти изменения имеют случайный характер и появляются спонтанно. Причины возникновения мутаций могут быть самыми разнообразными. Имеется целый ряд факторов, воздействие которых повышает вероятность возникновения мутаций. Это может быть воздействие определенных химических веществ, радиации, температуры и т. д. С помощью этих средств можно вызывать мутации, однако случайный характер их возникновения сохраняется и предсказать появление той или иной мутации невозможно.
Возникшие мутации передаются потомкам, т. е. определяют наследственную изменчивость, с одной важной оговоркой, связанной с тем, где произошла мутация. Если мутация произошла в половой клетке, то у нее есть возможность передаться потомкам, т. е. быть унаследованной. Если мутация произошла в соматической клетке, то она передается только тем клеткам, которые возникают из этой соматической клетки. Такие мутации называются соматическими, они не передаются по наследству.
Различают несколько основных типов мутаций:
1. Генные мутации, при которых изменения происходят на уровне отдельных генов, т. е. участков молекулы ДНК. Это может быть утрата нуклеотидов, замена одного основания на другое, перестановка нуклеотидов или добавление новых.
2. Хромосомные мутации, связанные с нарушением структуры хромосом. Они приводят к серьезным изменениям, которые могут быть обнаружены даже при помощи микроскопа. К таким мутациям относятся утраты участков хромосом (делеции), добавление участков, поворот участка хромосомы на 180°, появление повторов.
3. Геномные мутации, вызванные изменением числа хромосом. Мо гут появляться лишние гомологичные хромосомы, в хромосом ном наборе на месте двух гомологичных хромосом оказывают ся три — трисомия. В случае моносомии наблюдается утрата одной хромосомы из пары. При полиплоидии происходит кратное увеличение генома. Еще один вариант геномной мутации — гаплоидия, при которой остается только одна хромосома из каждой пары.
На частоту возникновения мутаций влияют самые разнообразные факторы. При возникновении ряда геномных мутаций большое значение имеет, в частности, возраст матери.
Наследственность и изменчивость. Комбинативная изменчивость. Данный тип изменчивости определяется характером полового процесса. При комбинативной изменчивости возникают новые генотипы из-за новых комбинаций генов. Этот тип изменчивости проявляется уже на стадии образования половых клеток. Как уже было сказано, в каждой половой клетке (гамете) представлена только одна гомологичная хромосома из каждой пары. Хромосомы попадают в гамету абсолютно случайным образом, поэтому половые клетки одного человека могут довольно сильно отличаться по набору генов в хромосомах. Еще более важная стадия для возникновения комбинативной изменчивости-это оплодотворение, после которого у вновь возникшего организма 50% генов унаследовано от одного родителя, а 50%-от другого.
№ 23
Классификация мутаций по действию на наследственные структуры:
Генные: цитологические невидимые изменения в хромосомах
Хромосомные: внутрихромосомные и межхромосомные перестройки
Геномные: изменение количества хромосом
Цитоплазматические: изменения плазмогенов
Хромосомные мутации.
Внутрихромосомные изменения. К внутрихромосомным изменениям относятся: нехватки части хромосом (делеции), умножение тех или иных участков хромосом (дупликации), изменение линейного расположения генов в хромосоме вследствие перевёртывания на 180 градусов отдельных участков хромосомы (инверсии).
К малым нехватках относят такие , которые связаны с выпадением нескольких хромомер или только части хромомеры – микроделеции. Микроделеции можно обнаружить только при исследовании гигантских хромосом. Мелкие нехватки обычно сохраняются в гомозиготном состоянии. Большие нехватки в гомозиготном состоянии летальны, что подтверждает большое значение каждого локуса хромосомы для жизни клетки и организма.
Межхромосомные изменения. Существуют изменения, связанные с обменом участками между негомологичными хромосомами. Такие межхромосомные перестройки называются транслокациями.
Тип расположения хромосом в виде 8 дают жизнеспособные сбалансированные гаметы, т. к. в этом случае к одному полюсу отходят либо обе изменённые хромосомы, либо обе неизменённые. Когда хромосомы расположены в мейозе в виде колец, образуются гаметы с несбалансированными генами: в одних – гены повторяются дважды, в других – они отсутствуют.
Мутационный процесс делят на спонтанный и индуцированный. В тех случаях, когда мутации возникают под влиянием обычных природных факторов внешней среды, их относят к спонтанным. Мутации, возникающие под влиянием специальных воздействий, называют индуцированными.
По месту возникновения мутации делят на генеративные и соматические.
Мутации также делят по признакам и свойствам, которые они определяют, т. е. по фенотипу, и по характеру изменений в генотипе.
Классификация мутаций по фенотипу. Мутации делят на морфологические, физиологические и биохимические.
Морфологические мутации связаны с изменением в строении или свойствах органов, тканей или отдельных структур клетки. К ним относится, например, коротконогость у с/х животных, безглазость и бескрылость у насекомых и т. д.
У различных организмов известно большое кол-во физиологических мутаций, вызывающих изменения физиологических процессов
К биохимическим мутациям относятся мутации, изменяющие качественно или количественно синтез определённых химических веществ в организме.
Классификация мутаций по характеру изменений генотипа.
Генные, или точковые мутации: цитологические невидимые изменения в хромосомах.
Хромосомные: внутрихромосомные м внешхромосомные перестройки.
Геномные: изменение количества хромосом.
Цитоплазматические: изменения плазмогенов.
Организмы, у которых изменён генотипназываются мутантами.
№ 25
Автополиплоидия — кратное увеличение гаплоидного набора хромосом в клетке. В результате автополиплоидии образуются полиплоидные организмы — триплоиды, тетраплоиды и т. д. Чаще всего полиплоидами являются растения. Этот тип мутаций может возникать при выпадении цитокинеза, завершающего процесс митоза, отсутствии редукционного деления во время мейоза, либо при разрушении веретена деления при делении клеток.
Классификация полиплоидов: эуплоиды, гаплоиды, триплоиды, тетраплоиды.
Триплоиды - особи, клетки которых содержат три основных (моноплоидных) набора хромосом.
Гаплоиды - особи, клетки которых содержат один основной (моноплоидный) набор хромосом.
Тетраплоиды - особи, клетки которых содержат четыре основных (моноплоидных) набора хромосом.
Роль полиплоидов в эволюции и селекции. Полиплоидизация нарушает скоррелированные физиологобиохимические системы и в ряде случаев обусловливает повышенное содержание ценных химических веществ или, наоборот, уменьшает синтез тех или иных нежелательных для человека соединений (например, соединения азота у полиплоидной свеклы). Полиплоиды могут приобретать и другие ценные признаки, например, повышение устойчивости к заболеваниям и др. В настоящее время хозяйственно ценные полиплоиды получены у ряда сельскохозяйственных культур: сахарной свеклы, проса, мака, льна, редиса, кукурузы, земляники и др.
Колхицин и его использование для получения полиплоидов. Колхицин — алкалоид трополонового ряда, основной представитель семейства колхициновых алкалоидов (гомоморфинанов). Чаще всего для получения полиплоидов используют алкалоид колхицин, так как он обеспечивает большой процент полиплоидных форм у всех сельскохозяйственных культур. Раствором колхицина воздействуют либо на сухие, наклюнувшиеся и проросшие семена, либо на точки роста растений более позднего возраста. Известно множество приемов обработки колхицином. Они различаются по концентрации раствора, экспозиции и технике воздействия. и кариотипирования.
Аллополиплоид ( аллоплоид) - полиплоидный организм, содержащий хромосомные комплексы двух или большего числа исходных видов.
Наиболее распространенным типом гибридов является аллотетраплоид, который обладает двумя наборами генов от каждого растения-родителя. Русский цитогенетик Карпеченко (1928) получил новый вид и даже род растений, скрестив разные виды — редьку (Raphinus) и капусту (Brassica). Эти виды близки друг другу и принадлежат к одному семейству крестоцветных (Cruciferae), представителем которого является и горчица. Каждое растение имеет диплоидный набор из 18 хромосом, но хромосомы редьки содержат некоторые гены, которые не встречаются в хромосомах капусты и наоборот. Гибрид Карпеченко имел в каждой клетке по 18хромосом, девять от редьки и девять от капусты. Во время мейоза разные наборы генов не могут образовать пары, и гибрид не дает потомства. Однако, помимо обычных генов, образовывались немногочисленные 18 — хромосомные гаметы, и в поколении F2 появлялось несколько аллотетраплоидов. Они были полностью плодородными, так как два набора хромосом от редьки и два от капусты обеспечивали образование парных гомологичных хромосом во время мейоза. Аллотетраплоид, или амфидиплоид, получил научное название Paphanobrassica. У этого растения корни напоминали капустные, а листья были как у редьки, но, к сожалению, он не имел экономической ценности. Однако на его примере удалось продемонстрировать принцип образования плодовитых межвидовых и межродовых гибридов. По меньшей мере половина из природных полиплоидов — аллополиплоиды, например культурная пшеница. Она имеет 42 хромосомы, представляющие полный диплоидный набор из 14 хромосом от каждого из трех предков.
Анеуплоидия (гетероплоидия) - изменение числа хромосом, не кратное гаплоидному набору. В результате возникают особи с аномальным числом хромосом: моносомики (2n - 1), у которых не хватает одной хромосомы в какой-либо паре, и полисомики, у которых одна из хромосом может быть повторена несколько раз (например, трисомики — 2n + 1, тетрасомики — 2n + 2). У человека описаны трисомии по различным парам хромосом. Так, одна добавочная хромосома у человека вызывает болезнь Дауна — физическую и умственную отсталость; недостаток одной Х-хромосомы у женщин приводит к потере признаков пола (моносомия).
Различают несколько типов анеуплоидии. При моносомии утрачивается одна из хромосом диплоидного набора (2n – 1). При полисомии к кариотипу добавляется одна или несколько хромосом. Частным случаем полисомии является трисомия (2n + 1), когда вместо двух гомологов их становится три. При нуллисомии отсутствуют оба гомолога какой-либо пары хромосом (2n – 2).
Основной механизм возникновения анеуплоидии - нерасхождение и потери отдельных хромосом в митозе и мейозе. Вследствие нарушения баланса хромосом анеуплоидия приводит к понижению жизнеспособности и нередко к гибели анеуплоидов, особенно у животных (анеуплоидия лежит в основе ряда хромосомных болезней). В генетическом анализе с помощью анеуплоидии (скрещивая мутантов с анеуплоидами по определенным хромосомам) определяют, в какой группе сцепления находится исследуемый ген.
№ 26
Понятие о виде и популяции. Вид – это основная систематическая единица, занимающая определённый ареал и представляющая совокупность родственных по происхождению особей, качественно отличных лт других видов и не скрещивающихся с ними. Популяцию можно определить как совокупность особей одного вида, заселяющих определённую территорию, свободно скрещивающихся друг с другом и в той или иной степени изолированных от других совокупностей особей данного вида. Вид представляет собой генетически закрытую систему, популяция – система генетически открытая.
Значение работ С.С. Четверикова по генетике популяций. Основополагающее значение для генетики популяций имели работы Четверикова. В 1926 г. он опубликовал статью «О некоторых моментах эволюционного процесса с точки зрения современной генетики», в которой обосновал учение о генетической структуре популяций, разработал методы генетического анализа наследственности популяций и показал, что все эволюционные события происходят внутри популяции. Если группа особей какого-либо вида размножается путём самоопыления и возможность перекрёстного опыления у таких растений совершенно исключена, то это не популяция. В любой группе растений-самоопылителей составляющие её отдельные линии могут обмениваться генетическим материалом, передавать возникающие наследственные изменения друг другу, и она может рассматриваться как потенциальная популяция. В популяциях самоопылителей рецессивные мутации быстро переходят в гомозиготное состояние, проявляются фенотипически и попадают под действие отбора.
Понятие о частотах генов и генотипов. Закон Харди-Вайнберга выражает вероятностные распределения генотипов в любой свободно скрещивающейся популяции. Действие этого закона предполагает выполнение ряда условий:
Популяция имеет неограниченно большую численность;
Все особи в популяции могут совершенно свободно скрещиваться;
Гомозиготные и гетерозиготные по данной паре аллелей особи одинаково плодовиты, жизнеспособны и не подвергаются отбором;
Прямые и обратные мутации происходят с одинаковой частотой или они так редки, что ими можно пренебречь.
Концентрация генов – это их относительная частота в популяции. По закону Харди – Вайнберга в свободно скрещивающихся популяциях исходное соотношение гомозигот и гетерозигот остаётся постоянным.
№ 27
Закон Харди — Вайнберга — это закон популяционной генетики — в популяции бесконечно большого размера, в которой не действует отбор, не идет мутационный процесс, отсутствует обмен особями с другими популяциями, не происходит дрейф генов, все скрещивания случайны — частоты генотипов по какому-либо гену будут поддерживаться постоянными из поколения в поколение и соответствовать уравнению:
p² + 2pq + q² = 1
Где p² — доля гомозигот по одному из аллелей; p — частота этого аллеля; q² — доля гомозигот по альтернативному аллелю; q — частота соответствующего аллеля; 2pq — доля гетерозигот.
Практическое значение закона Харди — Вайнберга.
В медицинской генетике закон Харди — Вайнберга позволяет оценить популяционный риск генетически обусловленных заболеваний.
В селекции — позволяет выявить генетический потенциал исходного материала.
В экологии — позволяет выявить влияние самых разнообразных факторов на популяции. По отклонениям фактических частот генотипов от расчётных величин можно установить эффект действия экологических факторов.
№ 28
Для генетических исследований человек является неудобным объектом, так как у человека: невозможно экспериментальное скрещивание; большое количество хромосом; поздно наступает половая зрелость; малое число потомков в каждой семье; невозможно уравнивание условий жизни для потомства.
В генетике человека используется ряд методов исследования.