

4.Реполяризація
Збільшення потоку іонів калію, спрямованого із клітини назовні, призводить до зменшення мембранного потенціалу, що, у свою чергу, обумовлює зменшення проникності мембрани для іонів натрію.
Таким чином, другий етап характеризується тим, що потік іонів калію із клітини назовні зростає, а зустрічний потік іонів натрію зменшується. Така реполяризація мембрани триває, доки не відбудеться відновлення потенціалу спокою — реполяризація мембрани. Після цього проникність для іонів калію також падає до вихідної величини. Зовнішня поверхня мембрани за рахунок позитивно заряджених іонів калію, що вийшли в середовище, знову набуває позитивного потенціалу відносно внутрішнього.
5.Слідові деполяризація і гіперполяризація
У кінцевій фазі відбувається уповільнення відновлення мембранного потенціалу спокою, і при цьому реєструються слідові реакції у вигляді деполяризації та гіперполяризації, обумовлені повільним відновленням вихідної проникності для іонів К+.
Фізіологічна роль ПД-полягає в тому,що він забезпечує збудження клітини та передачу цього збудження на інші клітини,а також підтримку та активацію у збудливих клітинах специфічних функцій.
6.Види транспорту йонів через мембрани клітин,значення для утворення електричних потенціалів.Йонні канали і помпи,їхні відмінності.
Транспорт іонів поділяється на активний і пасивний:
Активний транспорт речовин крізь мембрану здійснюється проти градієнта їхньої концентрації із затратою енергії АТФ та за участю спеціальних мембранних білків — транспортних АТФаз, які також називаються іонними насосами. Найбільш поширеними в клітині тварин є Н+-АТФаза, Na+,K+-АТФаза і Са2+-АТФаза, що являють собою цілі мембранні комплекси із складною структурою. Функціональне значення біологічних насосів полягає у підтримці всередині клітини постійного іонного складу. Na+,K+-АТФаза сприяє виведенню Na+ з клітини та надходженню К+ до клітини за допомогою енергії АТФ і є прикладом антипортного транспорту. Із впливом на натрієвий насос пов’язаний механізм дії деяких фармацевтичних препаратів. Так, напр. серцеві глікозиди (дигоксин, уабаїн, строфантин К) пригнічують Na+,K+-АТФазу; деякі діуретики (тіазиди) інгібують активний транспорт Na+ та/або Cl– в епітелії канальців нирок; омепразол знижує кислотність шлункового соку, незворотно пригнічуючи протонний насос Н+-АТФазу парієтальних клітин шлунка. Са2+-АТФаза високочутлива до дії різноманітних тіолових отрут та ін. Крім вищенаведених видів активного транспорту, виділяють специфічні механізми переміщення речовин, пов’язані з порушенням цілісності мембрани, ендоцитоз та екзоцитоз. При ендоцитозі плазматична мембрана утворює вирости, які потім перетворюються на внутрішньоклітинні пухирці, що містять захоплений клітиною матеріал. Ці процеси відбуваються із витратою енергії АТФ.
Пасивний транспорт відбувається мимовільно без затрати енергії шляхом дифузії, осмосу та полегшеної дифузії. Дифузія — це транспорт молекул та іонів через мембрану з ділянки з високою концентрацією до ділянки з низькою концентрацією, тобто речовини надходять за градієнтом концентрації. Дифузія може бути простою та полегшеною. Якщо речовини добре розчинні в ліпідах, то вони проникають до клітини шляхом простої дифузії. Напр. оксиген, потрібний клітині при диханні, та вуглекислий газ у розчині швидко дифундують крізь мембрани. Таким способом проникають до клітини також деякі фармацевтичні препарати, які є ліпідорозчинними. Вода також здатна проходити крізь мембранні пори, що утворені білками, і переносити молекули та іони речовин, які в ній розчинені. Дифузію води крізь напівпроникну мембрану називають осмосом. Вода переходить з ділянки з низькою концентрацією солей до ділянки, де їхня концентрація вища. Тиск на мембрану, що виникає при цьому, називають осмотичним. Усі живі клітини здатні регулювати осмотичний тиск, змінюючи концентрацію речовин поза клітиною та всередині клітини. Речовини, які не є розчинними у ліпідах, транспортуються через іонні канали, утворені в мембрані білками, чи за допомогою білків-переносників. Це полегшена дифузія, шляхом якої, напр. здійснюється надходження глюкози до еритроцитів. Серед систем пасивного транспорту важливу роль відіграють іонні канали, які забезпечують проникність мембрани для Na+, K+, Ca2+. Na+-канали активуються вератрадином, батрахотоксином, блокуються амілоридом, тріамтереном; K+-канали блокуються місцевими анестетиками (лідокаїном, дикаїном), деякими протисудомними (дифеніл, карбамазепін, вальпроати, фенобарбітал та ін.) і протиаритмічними засобами (аміодарон); Ca2+-канали чутливі до цілої низки хімічних речовин, зокрема верапамілу, дилтіазему, ніфедипіну та інших похідних дигідропіридинів. Трансмембранний обмін (антипорт) чи односпрямований транспорт (симпорт) іонів здійснюється спеціальними білками-переносниками. Система односпрямованого транспорту (котранспорту) представлена (Na++K++Cl–)-переносником, що є чутливим до дії діуретиків (фуросеміду, амілориду, туметаніду). Виключення або різка зміна властивостей переносників і каналів лежить в основі дії багатьох токсичних і фармацевтичних речовин. Деякі речовини — іонофори, до яких належать різні антибіотики (валіноміцин, амфотерицин В, нонактин, енніатини, аламетицин та ін.), синтетичні циклополіефіри самостійно здатні утворювати канали у ліпідному бішарі мембрани. Дія деяких ЛП заснована на зміні властивостей каналів і переносників, що дозволяє регулювати транспорт речовин у клітинах і організмі в цілому.
Іонні канали — трансмембранні білки, що утворюють пори через цитоплазматичну та інші біологічні мембрани, які допомагають встановлювати і управляти електричною напругою через мембрани всіх живих клітин (так званим мембранним потенціалом), дозволяючи рух певних іонів вниз за електрохімічним градієнтом.
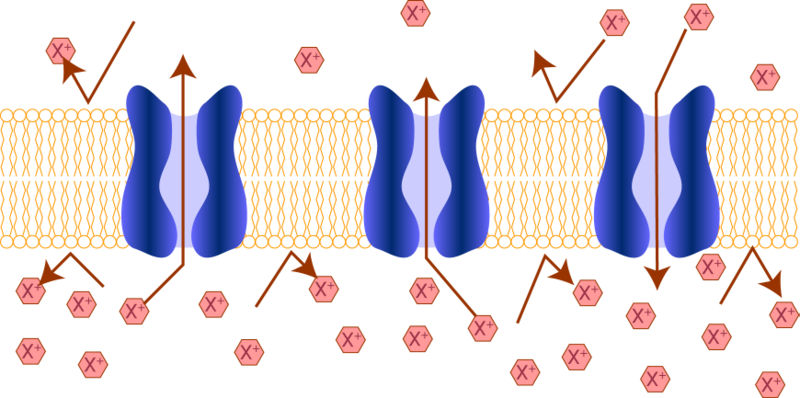
Селективнiсть iонних каналів багато в чому визначається геометрією каналу. Наприклад, кристалічний діаметр іона натрію – 0,19 нм, разом із гiдратною оболонкою його діаметр становить біля 0,3 нм. Устя каналу 0,3-0,5 нм. Для того, щоб пройти через канал (головне – через селективний фільтр), іон натрію або iнший іон повинен звільнитися від гiдратної оболонки і тільки в «голому» виглядi може пройти через канал. Занадто великий іон не може ввійти в канал, занадто маленький не спроможний віддати гiдратну оболонку в селективному фільтрі, тому не може вискочити з каналу.
Натрієві канали мають устя, селективний фільтр, ворiтний механізм. Ворота в них двох типів – активацiйнi (м-ворота) і iнактивацiйнi (h-ворота). В умовах спокою (МП=80 м, наприклад), активацiйнi ворота закриті, але готовi в будь-яку хвилину відчинитися, а iнактивацiйнi ворота відкриті. При зниженні МП (наприклад, до 60 м) активацiйнi ворота вiдчиняються і впускають іони натрію в клітину, але незабаром починають закриватися iнактивацiйнi ворота (відбувається iнактивацiя натрієвих каналів). Через якiсь час закриваються активацiйнi ворота, вiдчиняються iнактивацiйнi ворота, і канал готовий до нового циклу. Канал блокується тетродоксином, місцевими анестетиками (новокаїном, іншими речовинами). Це використовується в медичній практицi.
Калієві канали теж достатньо селективні – в основному пропускають іони калію. Вони блокуються тетраетиламонiєвим. Процеси iнактивацii в них виражені слабко.
Кальцієві канали – мають всі атрибути іонного каналу (устя, ворiтний механізм, фільтр). Блокуються іонами марганцю, нікелю, кадмiю (двовалентні іони), а також лікарськими речовинами – верапамiлом, нiфедипiном, дильтиаземом, що використовуються в клінічній практицi.
Робота іонних насосів
Коли м'яз скорочується, то, на це витрачається енергія. Це зрозуміло. Однією з показників витрати енергії є споживання кисню. Але виявляється, що кисень споживається і яка покоїться м'язом чи нервом. 1932-го р. М. Березина, працювала до лабораторій англійського біофізика А. Хілла, показала, наприклад, що нерв краба у спокої споживає 50% тієї кількості кисню, потрібної йому за посиленою роботі. У зв'язку з цими даними Хілл писав: «Отже, щось роблячи, а й просто перебуваючи готовності до відповідальності, нерв споживає близько половини тієї енергії, що він використовує за максимального відповіді». Зараз достеменно відомо, що цю енергію переважно витрачається га підтримку іонних концентрацій, отже, і підтримку потенціалу спокою.
Нагадаємо, що ПП переважно створюється з допомогою різниці концентрацій калію всередині клітини, де його багато, і поза клітиною, де її замало. Але мембрана проникна, хоча у невеличкий ступеня, також і іонів натрію. Іони натрію повинні проникати всередину клітини по градієнту потенціалу, а назовні натомість мають іони калію. У результаті процесу ПП повинен поступово знижуватися. Однак у живих організмах цього не відбувається. Існує якийсь механізм, який постійно підтримує ПП, отже, і різницю концентрацій калію усередині якого і поза клітини. Цей механізм повинен переміщати іони калію всередину клітини, тобто. туди, де з їхніми концентрація вище, ніж зовні, а такі переміщення проти градієнта концентрації вимагають витрати енергії.
Непрямі дані, що дають, що енергія спочиваючого нерва витрачається для підтримки ПП, отримано ще30-годи, коли Р. Джерард показав, що обсяг ПП в нервовому волокні прямо залежить від змісту кисню вомивающей волокно середовищі.
Вивчення механізму підтримки іонній концентрації є одним із найважливіших завдань біоенергетики – розділу молекулярної біології. Біоенергетика, що вивчає, звідки жива клітина отримує енергію та потім її витрачає, виявилася як найтісніше що зелектробиологией. Які ж молекулярні механізми підтримки іонних концентрацій?
Ми згадували, що у зовнішню клітинну мембрану вмонтовані різноманітних білкові молекули. Виявляється, дехто з тих молекул грають роль своєрідних насосів, «закінчуючи» іони калію всередину клітини, і викачуючи іони натрію назовні. Вони і називаються – «іонні насоси». Ці білки, дуже складно влаштовані, є справжню молекулярну машину, яка вміє робити дивовижні речі. Наприклад, показано, що вона не має два активних центру, однією із може захоплювати іон калію, іншим – натрію. Знайдено ще й «паливо», у якому працює ця машина. Це особливий хімічну сполуку –аденозинтрифосфорная кислота. Відомий також «коефіцієнт корисної дії» цього: досліди з радіоактивними ізотопами показали, що енергії розпаду однієї молекули АТФ достатньо викачування назовні трьох іонів натрію ізакачивания всередину клітини двох іонів калію. Як працює ця молекулярна машина, достеменно відомо, проте його можна уявити, наприклад, таку схему. Захопивши одним активним центром з зовнішньої середовища іон калію, іншим – із внутрішньої – іон натрію, вона, споживаючи АТФ, повертається всередині мембрани на 180°. Іон натрію виявляється поза клітини, і там відокремлюється, а іон калію потрапляє всередину і також звільняється, після чого молекула білка приймає початкове положення і всі починається спочатку. Цей білок, відкритий 1957 р. З.Скоу, зазвичай називаютьнатрий-калиевим насосом.
Якщо припинити подачу кисню до клітини, то ній кілька днів зникає АТФ і припиняється перекачування калію і натрію; тоді різниці концентрацій починають вирівнюватися і ПП починає падати. Якщо таку клітину запровадити АТФ, то насос відновлює роботи й ПП відновлюється. І це пояснює дослідиДже-рарда.
Ми знаємо, що згадані процеси в організмі регулюються. Серце бігуна б'ється втричі частіше, ніж в спокійно сидячого людини. Робота серця регулюється нервової системою. А чи можна якось регулювати роботу молекули, управляти молекулярної машиною?
Виявилося, робота іонних насосів управляється концентрацією іонів всередині клітини, і за її межами. У цьому робота насоса пришвидшується надлишком іонів калію зовні клітини чи надлишком іонів натрію всередині клітини.
Оскількинатрий-калиевий насос викачує назовні більше іонів натрію, ніж закачує всередину іонів калію, він змінює як концентрацію цих іонів, а й мембранний потенціал. Томунатрий-калиевий насос називаютьелектрогенним насосом. У кожному циклі роботи насос викидає назовні зайвий іон натрію і тим самимгиперполяризует мембрану. Після однієї чи кількох ПД у клітині виявляється надлишокКа+; це активує роботу насоса. Інтенсивно викачуючи натрій, насос може помітногиперполяризовать мембрану: МП може на 20мВ перевищувати ПП з допомогою роботи насоса. Отже, насоси як впливають на концентрації іонів, але можуть і джерелами помітної різниці потенціалів.