

7.2 Жирные кислоты
Жирные кислоты – алифатические карбоновые кислоты – в организме могут находиться в свободном состоянии (следовые количества в клетках и тканях) либо выполнять роль строительных блоков для большинства классов липидов.
В природе обнаружено свыше 200 жирных кислот, однако, в тканях человека и животных в составе простых и сложных липидов найдено около 70 жирных кислот, причем более половины из них в следовых количествах. Практически значительное распространение имеют немногим более 20 жирных кислот. Все они содержат четное число углеродных атомов, главным образом от 12 до 24. Среди них преобладают кислоты, имеющие С16 и С18 (пальмитиновая, стеариновая, олеиновая и линолевая). Нумерацию углеродных атомов в жирно-кислотной цепи начинают с атома углерода карбоксильной группы. Примерно 3/4 всех жирных кислот являются непредельными (ненасыщенными), т.е. содержат двойные связи. Ненасыщенные жирные кислоты человека и животных, участвующие в построении липидов, обычно содержат двойную связь между (9-м и 10-м атомами углеводородов); дополнительные двойные связи чаще бывают на участке между 11-м атомом углерода и метильным концом цепи. Своеобразие двойных связей природных ненасыщенных жирных кислот заключается в том, что они всегда отделены двумя простыми связями, т.е. между ними всегда имеется хотя бы одна метиленовая группа (—СН=СН—СН2—СН=СН—). Подобные двойные связи обозначают как «изолированные».
Таблица 10. Некоторые физиологически важные насыщенные жирные кислоты
Число атомов С |
Тривиальное название |
Систематическое название |
Химическая формула соединения |
6 |
Капроновая |
Гексановая |
СН3−(СН2)4−СООН |
8 |
Каприловая |
Октановая |
СН3−(СН2)6−СООН |
10 |
Каприновая |
Декановая |
СН3−(СН2)8−СООН |
12 |
Лауриновая |
Додекановая |
СН3−(СН2)10СООН |
14 |
Миристиновая |
Тетрадекановая |
СН3−(СН2)12−СООН |
16 |
Пальмитиновая |
Гексадеконовая |
СН3−(СН2)14−СООН |
18 |
Стеариновая |
Октадекановая |
СН3−(СН2)16−СООН |
20 |
Арахиновая |
Эйкозановая |
СН3−(СН2)18−СООН |
22 |
Бегеновая |
Докозановая |
СН3−(СН2)20−СООН |
24 |
Лигноцириновапя |
Тетракозановая |
СН3−(СН2)22−СООН |
В растворах жирно-кислотная цепь может образовывать бесчисленное количество конформаций вплоть до клубка, в котором имеются и линейные участки различной длины в зависимости от числа двойных связей. Клубки могут слипаться между собой, образуя так называемые мицеллы. В последних отрицательно заряженные карбоксильные группы жирных кислот обращены к водной фазе, а неполярные углеводородные цепи спрятаны внутри мицеллярной структуры. Такие мицеллы имеют суммарный отрицательный заряд и в растворе остаются суспендированными благодаря взаимному отталкиванию.
Известно также, что при наличии двойной связи в жирнокислотной цепи вращение углеродных атомов относительно друг друга ограничено. Это обеспечивает существование ненасыщенных жирных кислот в виде геометрических изомеров, причем природные ненасыщенные жирные кислоты имеют цис-конфигурацию и крайне редко транс-конфигурации.
Таблица 11. Некоторые физиологически важные ненасыщенные жирные кислоты
Число атомов С |
Тривиальное название |
Систематическое название |
Химическая формула соединения |
Моноеновые кислоты |
|||
16 |
Пальмитиновая |
9-гексадеценовая |
СН3−(СН2)5−СН=СН−(СН2)5СООН |
18 |
Олеиновая |
9-октаддеценовая |
СН3−(СН2)7−СН=СН−(СН2)7СООН |
Диеновые кислоты |
|||
18 |
Линолевая |
9,12-октадеценовая |
СН3−(СН2)4−СН=СН−СН2− −СН=СН− (СН2)7СООН |
Триеновая кислоты |
|||
18 |
Линоленовая |
9,12,15-октадекатриеновая |
СН3−СН2−СН=СН−СН2− −СН=СН− СН2− СН=СН (СН2)7−СООН |
Тетраеновые кислоты |
|||
20 |
Арахидоновая |
5,8,11,14-эйкозатетраеновая |
СН3−СН2−СН=СН−СН2− СН=СН−СН2− СН=СН−СН2− СН=СН−СН2− СН= − (СН2)5−СООН |
Считают, что жирной кислоте с несколькими двойными связями цис-конфигурация придает углеводородной цепи изогнутый и укороченный вид. По этой причине молекулы этих кислот занимают больший объем, а при образовании кристаллов упаковываются не так плотно, как транс-изомеры. Вследствие этого цис-изомеры имеют более низкую температуру плавления (олеиновая кислота, например, при комнатной температуре находится в жидком состоянии, тогда как элаидиновая – в кристаллическом). Цис-конфигурация делает ненасыщенную кислоту менее стабильной и более подверженной катаболизму.
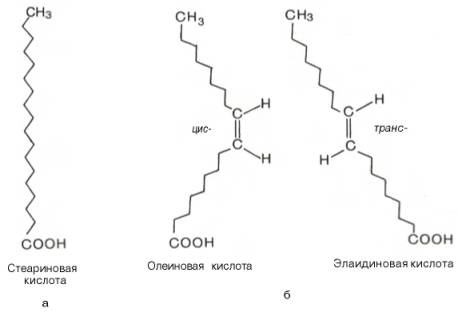
Рис.23 Конфигурация 18-углеродных насыщенных (а) и мононенасыщенных (б) жирных кислот.