

6.8.4. Координація рефлекторних процесів
Координацією називають взаємодію між нейронами і нервовими процесами, що спрямована на інтеграцію функцій різних органів і систем. Нервова система організовує доцільну рефлекторну відповідь тільки завдяки взаємоузгодженості між усіма фізіологічними процесами. Взаємоузгодженість стає можливою в результаті дії механізмів, які грунтуються на двох групах принципів - морфологічних і функціональних.
1) Конвергенція. Імпульси, що надходять у центральну нервову систему різними аферентними волокнами, можуть сходитись, або конвергувати, до одних і тих же проміжних або ефекторних нейронів. У спинному і довгастому мозку конвергенція має обмежений характер. На вставних нейронах і мотонейронах конвертують аферентні імпульси з різних ділянок одного і того ж рецептивного поля. У підкоркових ядрах і корі здійснюється конвергенція імпульсів, які виникають у різних рефлексогенних зонах. Конвергенцію забезпечують чисельні синапсами на тілі і дендритах нейронів. Конвергенція лежить в основі просторової сумації і оклюзії.
2) Окпюзія (закупорка). На нейронах нервових центрів закінчуються не тільки волокна їх власних аферентних входів, а й інших центрів. Така морфологічна особливість може зумовити оклюзію або, навпаки, полегшення.
Явище оклюзії полягає у тому, що при максимальному одночасному подразненні двох аферентних нервів рефлекс може бути менший, ніж арифметична сума рефлексів, одержаних при подразненні окремо кожного з аферентних нервів.
Механізм виникнення оклюзії зображено на рис. 5.
Рис. 5. Схема явища оклюзії
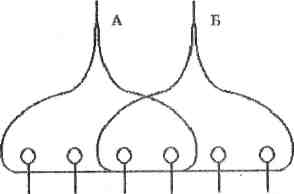
На цьому рисунку зображено мотонейрони, що одержують імпульси від двох груп аферентних волокон А і Б. При максимальному подразненні волокон А збуджуються 4 мотонейрони, при максимальному подразненні волокон групи Б -теж 4. Слід би очікувати, що при одночасному подразненні волокон групи А і Б будуть збуджуватись 8 нейронів. Проте збуджуються лише 6 мотонейронів, оскільки внаслідок конвергенції два нейрони отримують одночасно імпульси по волокнах А і Б.
3) Просторове полегшення зумовлене тим, що рецепторний нейрон утворює у нервовому центрі синапси з популяцією нейронів. У такій популяції розрізняють центральну (порогову) ділянку і периферичну (підпорогову) ділянку (див. рис. 6). На нейронах порогової ділянки утворюється декілька синапсів (на рисунку по 2), а на нейронах підпорогової - їх менше (на рисунку по 1). Внаслідок цього при роздільному подразненні аферентних волокон А і Б виникають рефлекторні реакції, сила яких визначається кількістю збуджених нейронів центральних зон (по 3). Одночасне подразнення волокон А і Б збуджуватиме і нейрони підпорогової ділянки (2). Тому сила рефлекторної реакції буде більшою (3+3+2=8), ніж арифметична сума реакцій (3+3=6), що викликані роздільним подразненням волокон.
4) Іррадіація. Імпульси, що надходять у центральну нервову систему під час сильгіого і тривалого подразнення, викликають збудження не тільки нейронів цього рефлекторного центру, а й інших центрів. Таке поширення збудження від одних центрів до інших називають іррадіацією. Іррадіація збудження проходить завдяки наявності чисельних розгалужень відростків нейронів, а також наявності вставних нейронів. Особлива роль у механізмах іррадіації належить ретикулярній формації. Протидіють іррадіації гальмівні нейрони і гальмівні синапси.
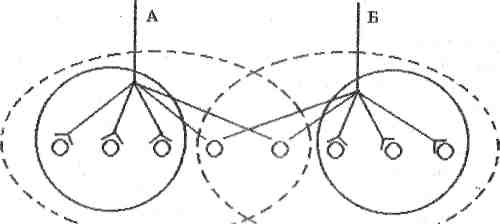
Рис. 6. Схема явища просторового полегшення
5). Індукція. Взаємовідносини між нервовими центрами визначаються так званими індукційними процесами. Розрізняють одночасну (негативну і позитивну) індукцію, а також послідовну (негативну і позитивну) індукцію. Якщо збудження певного центру викликає гальмування іншого, таке явище називають одночасною негативною індукцією. Якщо гальмування одного центру веде до збудження іншого, - це приклад одночасної позитивної індукції. Наприклад, збудження центру м'язів-згиначів зумовлює гальмування центру м'язів-розгиначів; збудження центру ковтання гальмує центр вдиху.
Послідовна індукція проявляється у посиленні активності попередньо загальмованого центру (послідовна позитивна індукція) або у гальмуванні попередньо збудженого центру (послідовна негативна індукція).
6) Реципрокна іннервація. Принцип реципрокної, або спряженої, іннервації, найкраще продемонструвати на прикладі іннервації м'язів-антагоністів кінцівок. Згідно цього принципу, збудження центру однієї групи м'язів супроводжується реципрокним гальмуванням центрів м'язів-антагоністів (одночасна негативна індукція). Так, збудження центру м'язів-згиначів однієї кінцівки спричинює гальмування центрів-розгиначів цієї ж кінцівки. Одночасно збуджується центр м'язів-розгиначів і гальмується центр м'язів-згиначів протилежної кінцівки. Реципрокні взаємовідносини між процесами збудження і гальмування забезпечують послідовність рухбвих актів і локомоцію в цілому.
7) Принцип загального кінцевого шляху. Один і той же руховий рефлекс може бути викликаний подразненням різних рецепторів. Це означає, що одні і ті ж мотонейрони входять до складу багатьох рефлекторних дуг. Отже, мотонейрони утворюють загальний кінцевий шлях різних рефлексів і можуть бути зв'язані з будь-якими рецепторними нейронами. Такі зв'язки здійснюються за допомогою контактних нейронів. Кількість рецепторних нейронів приблизно у 5 разів більша, ніж ефекторних.
8) Принцип домінанти. Прикладом взаємодії рефлексів служить встановлений О.О.Ух томським (1923) феномен домінанти. Домінантою називають тимчасово панівний у центральній нервовій системі осередок стійкого збудження, який змінює і підкоряє собі роботу всіх інших центрів. Виникнення домінантного осередку збудження завжди веде до більш-менш вираженого спряженого гальмування інших центрів. Домінантний осередок збудження характеризується: 1) підвищеною збудливістю; 2) стійкістю збудження; 3) здатністю до сумації; 4) здатністю до гальмування інших центрів. Формування у центральній нервовій системі домінантного центру веде до того, що подразнення різних рецептивних полів спричиняє рефлекторну відповідь, яка характерна для домінантного центру.
Домінанта може бути екзогенною, ендогенною і такою, що викликана експериментально. Наведемо приклади експериментально встановленої домінанти. Подразнення моторної ділянки кори, збудження якої спричиняє згинання передньої кінцівки, під час рефлексу ковтання підсилює цей рефлекс, але не викликає згинання.
Екзогенна домінанта виникає під впливом подразників зовнішнього середовища. Прикладом екзогенної домінанти, викликаної зниженою температурою взимку у жаб, є щільне підтягування задніх кінцівок до тулуба. Будь-який зовнішній вплив підсилює цей процес. Ендогенна домінанта виникає внаслідок зміни внутрішнього середовища, наприклад, концентрації глюкози або статевих гормонів. Ендогенна домінанта спостерігається у самців жаб весною під впливом статевих гормонів. Вона проявляється у тому, що обнімальний рефлекс підсилюється при будь-якому подразненні шкіри.
Домінантними стають, звичайно, ті центри, що пов'язані із задоволенням життєво важливих у даний момент потреб. У людини і вищих тварин виникають харчова, статева, оборонна й інші домінанти.