

1.3. Эколого-фитоценотические стратегии у орхидных. Орхидные-патиенты (ps-орхидные) и орхидные-патиент-эксплеренты (rs-орхидные)
1.3.1 Адаптивные признаки орхидных-патиентов
По характеру поведения в фитоценозе почти все орхидные являются патиентами, которые требуют специфических экологических условий, но сами не создают их и при этом плохо приспосабливаются к изменившимся условиям.
Орхидные – одно из тех семейств высших растений, большая часть которого приспособлена к жизни в неблагоприятных условиях (засухи или заморозков, засоления или временного просыхания субстрата, дефицита света или избытка радиации, и т.д.). Все эти стрессовые воздействия орхидные переживают с помощью специальных морфологических адаптаций (особого строения воздушных корней, метаморфозов листьев и побегов), ритмов развития (периодов покоя), специфического метаболизма кислот (САМ). Можно сказать, что все вегетативное тело орхидей (в особенности эпифитных) состоит из метаморфизированных органов. Многие адаптивные приспособления морфоструктур у орхидных появились в связи с изменениями способа фиксации углерода и водного баланса в различных климатических условиях. Приведенная схема теоретически возможных вариантов адаптивной эволюции учитывает тип метаболизма, биотические связи (эпифитизм, микотрофизм), а также периодичность роста орхидных, распространившихся в разных экологических нишах (рис. 2).
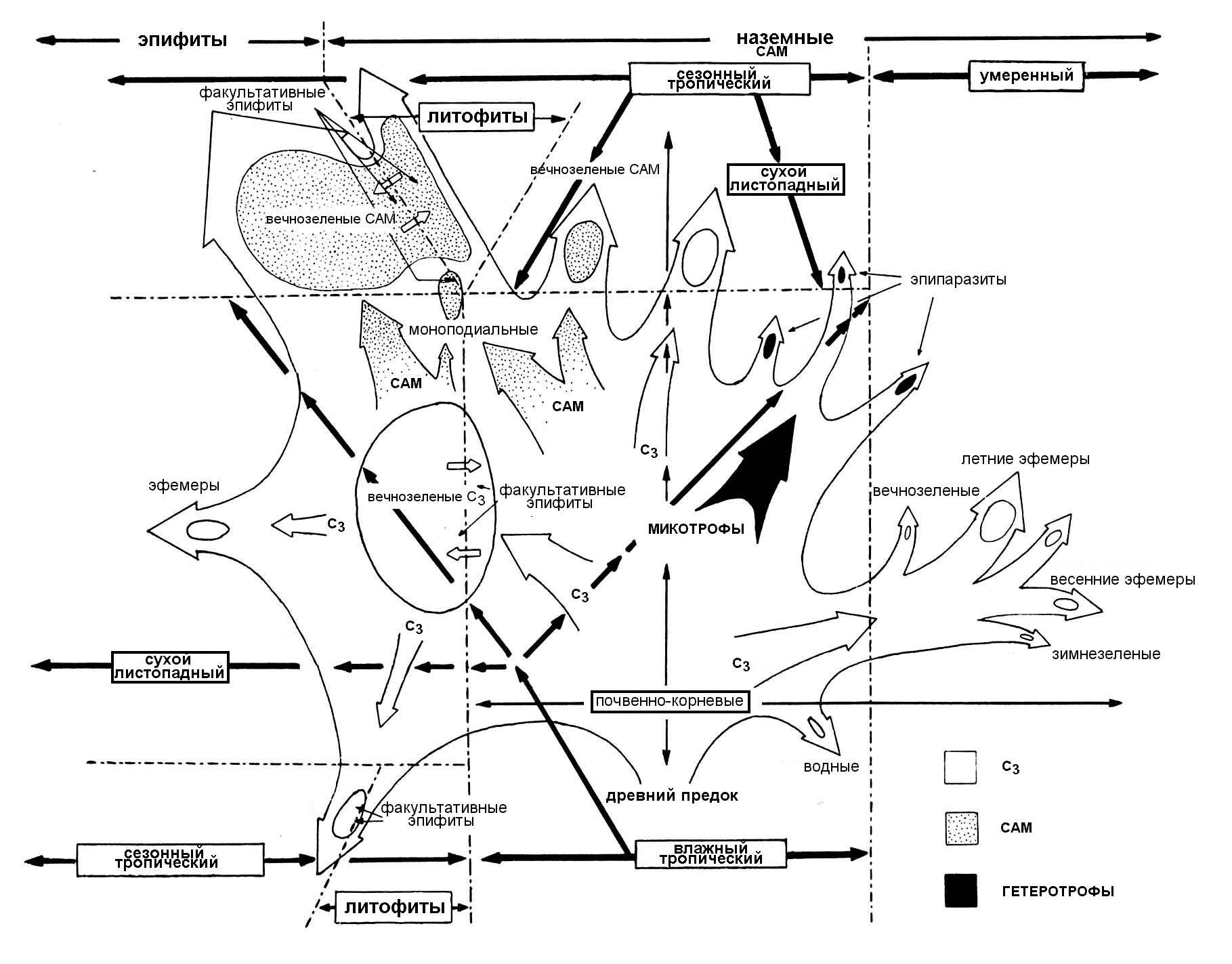
Рисунок 2. Схема предполагаемых путей адаптивной эволюции орхидных в зависимости от типа метаболизма
В фитоценозах популяции орхидных из любых экологических ниш (эпифитных, литофитных, наземных) обычно разомкнуты и, как правило, состоят из ограниченного числа особей, и, конечно, каждый вид имеет индивидуальные черты адаптации к своей экологической нише. В лесных фитоценозах тропической и субтропической зон эпифитные орхидеи занимают ниши в пологе деревьев-хозяев. В зонах саванн, пустынь, тайги и тундры геофитные орхидные со стебле-корневыми тубероидами перешли к эфемероидному образу жизни как весеннее-зеленые, летнее-зеленые или зимнее-зеленые травы, переживающие неблагоприятные (слишком высокие или слишком низкие температуры) в виде подземных клубней или стебле-корневых тубероидов.
Большая часть орхидных относится к подтипу экотопических патиентов, которые приспособлены к жизни в исходно бедных местообитаниях под постоянным прессингом абиотических стрессовых воздействий. Основную группу экотопических патиентов составляют эпифитные орхидные, которые по оценкам разных авторов составляют около 70-75 % от всех орхидных, представленных во флоре Земного шара.
1.3.2 Эпифитные экологические ниши орхидных с ps-стратегией
Экологические ниши эпифитных орхидей, то есть их распределение в пологе дерева, имеют весьма размытые границы.
Большинство исследователей предпочитают делить эпифиты по зонам, которые они занимают в кроне, демонстрируя количественные и видовые различия эпифитной флоры в каждой зоне. Наиболее простая градация предполагает деление надземной части дерева - хозяина на две зоны и выделение настоящих эпифитов, растущих в кроне, а также полуэпифитов, заселяющих ствол. Также выделиляют три зоны, определив, соответственно три экологические ниши для эпифитных орхидей: гумусные эпифиты, поселяющиеся в небольших углублениях на коре, заполненных слоем гумуса, коровые эпифиты, поселяющиеся на стволах и больших ветвях и эпифиты тонких ветвей. Можно разделить модельное дерево на пять зон - базальную часть ствола, ствол, базальную часть больших ветвей, среднюю часть больших ветвей и наружную часть кроны. В результате создана теоретическая модель возможной корреляции между двумя группами экологических факторов и распределением эпифитных орхидей вдоль больших ветвей дерева-хозяина. Согласно этой модели, в первую группу факторов входят относительная влажность воздуха, структура и влажность коры дерева, а также наличие на ней слоя гумуса. Во вторую группу факторов входят освещенность, температура воздуха и сила ветра. По направлению от ствола к концам ветвей влияние первой группы факторов ослабевает, а второй усиливается, и наоборот.
Считают, что определяющим фактором в заселении полога дерева является степень доступности влаги - постоянная или временная. Постоянно снабжаемые водой CS-эпифиты (continuous supplied) растут на гигроскопичном гумусном субстрате из опада или в слое влажного мха; большинство временно снабжаемых водой эпифитов – PS-эпифиты (pulse supplied) потребляют влагу с негигроскопичного, быстро просыхающего субстрата. Как правило, PS-эпифиты - это светолюбивые миниатюрные орхидеи (их называют также «солнечными» или гелиофильными эпифитами), которые заселяют наружную часть кроны. Здесь они находятся в экстремальных условиях из-за частого пересыхания и бедности субстрата, воздействия ветра и прямой солнечной радиации. PS - эпифиты имеют самый короткий интервал от прорастания семени до первого цветения, поскольку им нужно успеть обсемениться и оставить потомство в перерыве между длительными засухами. Некоторые PS-эпифиты имеют удлиненные корневища и легко ветвятся, быстрым вегетативным расселением заменяя семенное размножение, затрудненное в таких суровых условиях.
Очень часто орхидные PS-эпифиты имеют вальковатые или мясистые листья с устьицами на обеих сторонах листа и характеризуются особым метаболизмом кислот, впервые описанным у семейства толстянковых (так называемый Crassulacean Acid Metabolism или САМ-метаболизм) и свойственный суккулентным растениям аридных зон. Вода и углекислый газ для фотосинтеза у таких растений запасаются ночью, когда градиент концентрации водяных паров между листьями и окружающим воздухом минимален, устьица растений в это время открыты и углекислый газ запасается в тканях листьев в виде яблочной кислоты или малата. Днем, при закрытых устьицах, малат декарбоксилируется, а выделяющийся при этом углекислый газ используется в фотосинтетических реакциях. Это позволяет PS-эпифитам днем, в самое жаркое время, держать устьица закрытыми, сводя потери воды к минимуму. САМ-метаболизм не встречается у наземных орхидных из подсемейств Apostasioideae, Сypripedioidea, Orchidoideae, в то время как у представителей подсемейств Epidendroideae и Vandoideae этот тип фиксации углекислого газа встречается довольно часто.
Орхидные CS-эпифиты более пластичны, при определенных изменениях климатических факторов они могут перемещаться вдоль стволов и ветвей, переходя из одной экологической ниши в другую. Это во многом объясняется особенностями их корневой системы, способной развивать как воздушные, так и субстратные корни.
К орхидным CS-эпифитам можно причислить все стволовые эпифиты, а также эпифиты внутренних частей кроны дерева-хозяина. Безусловно, этот тип эпифитов представлен большинством видов орхидных в разных растительных сообществах тропиков земного шара.
Роль эпифитов в растительных сообществах невелика. Поэтому фитоценотические исследавания эпифитов проводят обычно в микромасштабе, на уровне так называемых фитогенных полей. Внутри фитогенного поля (сферы влияния одного растения-хозяина на окружающую его среду и, соответственно, на занимающие эту среду эпифитные растения) взаимоотношения с деревом-хозяином часто играют определяющую роль при поселении на нем эпифитов.
Аллелопатические связи между деревьями и эпифитными орхидеями могут проявляться через угнетение или стимуляцию роста эпифитов, особенно на начальных стадиях их прорастания, наиболее часто влияют на прорастание: органические кислоты, альдегиды, эфирные масла, алкалоиды, фенолы, а также этилен и другие. Изучая прорастание Encyclia tampensis (Lindl.) Small на коре разных видов деревьев, выявлено, что кора исследованных Annona sp. и Taxodium sp. выделяет вещества, задерживающие образование корней у сеянцев орхидей. Некоторые виды эвкалиптов, выделяют таннины и фенолы, токсичные для эндомикоризных грибов, и препятствующие симбиотическому прорастанию семян эпифитных орхидных в своем пологе.
Геофитные орхидные с фотосинтезирующими листьями срединной формации, переживающие неблагоприятные погодные условия в земле (как эфемероиды), занимают самые разнообразные места обитания – от болотистых таежных лесов до полупустынь и прерий. И если в умеренном климате почва защищает почки возобновления геофитных орхидных от низких температур, то ближе к экватору почва обеспечивает защиту почек от перегрева и недостатка влаги. Например, в прериях геофитные орхидные периодически подвергается пожарам и засухам, конкурируют с С-3 и С-4 травами и часто встречаются в нарушенных местообитаниях. В ответ на пожар, геофитные орхидные могут сохранять свои почки в покое и оставаться под землей в течение нескольких лет, потребляя питательные вещества, накопленные в утолщенных корнях и стебле-корневых тубероидах и осуществляя метаболическую активность с помощью эндомикоризных грибов. Надземная часть растения может пострадать от пожара, но подземные части, как правило, остаются незатронутыми огнем. Некоторым африканским орхидеям огонь необходим для цветения, они редко приступают к репродукции в другое время. Южно-африканская эндемичная орхидея Brachycorythis macowaniana Rchb.f. зацветает только после пожара. После пожара прорастают семена Disa neglecta Sand., D. sabulosa Bolus.
Адаптации геофитных орхидных к летней засухе включает быстро отмирающие листья, отдающие накопленные в них питательные вещества подземным утолщенным клубням стебле-корневой структуры (стеблекорневым тубероидам), а также предрасположенность к автоопылению и даже к агамоспермии. Высокий уровень агамоспермии был выявлен в популяциях некоторых видов геофитных орхидных, произрастающих в прериях.
К сожалению, у большинства аллогамных видов геофитных орхидных семенная продуктивность, зависящая от эффективности опыления, результативности плодообразования и флуктуации эмбриологических нарушений, сводится к минимальной эффективности прорастания, поскольку взаимодействие с грибом-симбионтом носит вероятностный характер.
В то же время пределы адаптационной способности вегетативно зрелых особей геофитных орхидных-патиентов иногда чрезвычайно широки. Так, например, геофитный американский вид Spiranthes cernua (L.) Rich. в течение более 25 лет выращивался в условиях Фондовой оранжереи ГБС РАН как полуводное растение с полностью погруженными под воду утолщенными корнями. Выжить в таких условиях экстремального переувлажнения виду помогла способность к образованию деток на корнях - корнеотпрысковое вегетативное размножение. При этом генеративное размножение полностью отсутствовало - растения регулярно цвели, но, в связи с отсутствием естественных опылителей, никогда не завязывали семян.
С другой стороны, в природных местах обитания этот вид способен переживать продолжительную засуху. Обычно это происходит в середине лета, более чем за месяц до цветения. Рано сбрасывая листья, S. cernua затем развивает соцветие. Цветки опыляются шмелями из рода Bombus, но наблюдается и апомиксис. Cпособность вида к регулярному образованию корневых отпрысков в естественных популяциях не описана.
Особую группу геофитных патиентов составляют ахлорофилльные микотрофы с редуцированными листьями срединной формации (Gastrodia elata Blume), у которых репродуктивное усилие определяется длительным подземным накоплением фитомассы с помощью эндомикоризных грибов и образованием единственного соцветия (истинный монокарпик). Как правило, значительное репродуктивное усилие во время длительного накопления питательных веществ для единственного цветения окупается у таких симбиотрофных видов успешностью автогамного опыления. Однако число семязачатков, приходящееся на единицу фитомассы особи все-таки остается небольшим, в связи с чем микосимбиотрофные ахлорофилльные виды всегда проявляют свойства истинной патиентности.
Таким образом, патиентность геофитных орхидных определяется не только вкладом фитомассы в органы репродукции или органы фотосинтеза, но и способностью вида с максимальной эффективностью использовать консортивных партнеров – опылителей и микосимбинтов.